Anticipating critical transitions
1
2012
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
China's urbanization and its ecological environment challenges in the 21st century
1
2019
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
21世纪中国的城市化特点及其生态环境挑战
1
2019
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
Water quality characteristics and variations of Lake Dian
1
2017
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
滇池水质特征及变化
1
2017
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
A drinking water crisis in Lake Taihu, China: Linkage to climatic variability and lake management
1
2010
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
Nutrient dynamics and the eutrophication of shallow lakes Kasumigaura (Japan), Donghu (PR China), and Okeechobee(USA)
2001
Recent advances in the understanding and management of eutrophication
1
2006
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
Succession of macrozoobenthic communities and implications for ecological restoration in an urban Lake Wuli,Jiangsu Province
1
2020
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
典型城市湖泊五里湖底栖动物群落演变特征及其生态修复应用建议
1
2020
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
Phosphorus control is critical to mitigating eutrophication
1
2008
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
1
2011
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
1
2011
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
Self-purification capacity of nitrogen and phosphorus of Lake Taihu on the basis of mass balance
2
2014
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
基于质量平衡的太湖氮、磷自净能力计算
2
2014
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Direct evidence of the explosive release of endogenous phosphorus in large shallow lakes caused by wind wave disturbance
2
2005
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
风浪扰动引起大型浅水湖泊内源磷暴发性释放的直接证据
2
2005
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Effects of common carp on aquatic ecosystems 80 years after "Carp as a Dominant": Ecological insights for fisheries management
2
2009
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
The influence of Tubificid worms bioturbation on the exchange of phosphorus across sediment-water interface in lakes
1
2010
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
水丝蚓(Tubificid worms)扰动对磷在湖泊沉积物—水界面迁移的影响
1
2010
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
Variation trend of total phosphorus and its controlling factors in Lake Taihu, 2010-2017
1
2019
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
2010—2017年太湖总磷浓度变化趋势分析及成因探讨
1
2019
... 社会经济的不合理发展给自然生态系统带来巨大冲击,使得很多生态系统面临突变风险[1].湖泊是地球陆地系统的重要资源,为流域居民提供诸多生态系统服务.人类活动如农业化肥施用、工业排放、鱼类养殖等导致湖泊富营养化日趋严重[2,3].营养盐增加导致的蓝藻水华暴发,给居民生活造成了很大的影响[4~6].如何应对湖泊富营养化是目前的重大难题之一,备受人们关注[7].磷是湖泊的重要营养元素[8],是很多湖泊藻类爆发的限制性因子[9],也是评价湖泊营养状态的关键指标.湖水中的磷主要来自外源负荷和内源负荷两个方面.多数湖泊的外源负荷主要由入湖径流产生,如太湖2010年入湖径流所携带的磷负荷为2 800 t,滞留率约为52%[10].而内源负荷主要来源于底泥中的营养盐释放.相关研究表明,风浪扰动[11]、鱼类扰动[12]和底栖动物的扰动[13]等均能促进底泥的再悬浮从而引起内源磷的释放.与湖泊中氮循环不同,湖泊中磷的流失主要是靠出湖径流,磷呈气态逸出量占比非常小[14]. ...
The responses of the benthic-pelagic coupling to eutrophication and regime shifts in shallow lakes: Implication for lake restoration
1
2020
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
浅水湖泊底栖—敞水生境耦合对富营养化的响应与稳态转换机理:对湖泊修复的启示
1
2020
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
3
1998
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
... [16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
... [16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
Phytoplankton and their dynamics in oligotrophic and eutrophic lakes
1
1978
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
Shallow lakes from the central plains of argentina: An overview and worldwide comparative analysis of their basic limnological features
1
2015
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
Lake and catchment management in Denmark
1
1999
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
Early-warning signals for critical transitions
2
2009
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
... 图1b展示了模型模拟的浅水湖泊的稳态转换过程.在驱动力梯度上系统状态存在2个突变点,即F1和F2,当驱动力越过临界点时,生态系统状态会发生临界转换.在F1和F2之间,平衡曲线向后折叠,因此,该模型又被称为折叠交叉模型[43].在此条件区间内系统存在3个平衡.上分支和下分支曲线都是实线,代表稳定的平衡,中间虚线部分是指不稳定的平衡,代表了上分支和下分支2个稳定状态吸引域的边界.图1中的灰色虚线箭头是指如果系统不处于平衡状态时(即不在曲线上时),系统移动的方向.随着驱动力的增加,系统的稳态没有发生改变,仍在上分支,但是系统的吸引域或弹性会随着驱动力的增加而减小,当系统接近折叠灾变点F2时,驱动力的微小变化都可能导致系统突变为下分支的稳态.但是系统若想从下分支回到上分支,驱动力减小到F2大小时是远远不够的,要继续减小,当减小到F1时,系统才能从下分支突变到上分支.2个稳态相互转换发生在不同的驱动力突变点下,这种情况就称为滞后效应[20].具体到湖泊生态系统,图2表现了水体总磷浓度在外源驱动下出现的清水态和浊水态之间稳态转换现象和滞后效应.磷动力学模型结果如图2a(P-t图)和图2b(P-α图)所示.图2a中存在突变时间A和B,其中A是指湖水磷浓度由低浓度突变到高浓度的突变时间,B是指湖水磷浓度从高浓度降回低浓度时的突变时间.图2b中同样存在2个突变点F1和F2,突变点F2是指改变参数后,清水稳态突变为浊水稳态时外源磷输入的阈值大小,突变点F1是指由浊水稳态修复回清水稳态时的外源磷输入值.X(F1,F2)是指稳态间转变的滞后时长;稳态大小是指湖泊由清水态或者浊水态时的P浓度. ...
Catastrophic regime shifts in ecosystems: Linking theory to observation
3
2003
... 湖水磷浓度的上升会造成浅水湖泊生态系统状态发生改变[15].在低磷浓度的水平上,湖水清澈透明,沉水植物占优势,表现为清水态湖泊[16];而在高磷浓度的水平上,水体浮游藻类占优势,透明度下降,表现为浊水态湖泊[17,18].随着外源负荷的不断增多,一旦湖水中营养物质浓度超过某个阈值时,沉水植物消失,藻类大量生长,湖泊就会从清水态转变为浊水态[19],发生稳态转换[16].已有模型研究表明,在低营养状况下,湖泊外源营养的增加不会显著改变湖泊生态系统结构,湖泊仍能够维持清水态[16];但是持续的营养增加会逐渐破坏生态系统结构,当外源营养盐输入达到一定阈值时,微小的营养盐增量就能促使生态系统结构发生重大调整,湖泊从清水态突变到浊水态,生态系统功能严重下降[20].模型研究结果还表明,一旦湖泊生态系统发生稳态转换,在一定的营养盐区间内,湖泊可以存在多个“稳态”[21],使得湖泊修复存在“时滞效应”,即要使得湖泊生态系统恢复到稳定的清水态,必须将外力驱动降的更低.因此,湖泊生态系统一旦发生稳态转换,修复将变得非常困难.在湖泊生态系统管理中,一方面要避免湖泊从清水态到浊水态之间稳态转换的发生;另外一方面,要促使湖泊浊水态向清水态转换的尽快发生.对湖泊稳态转换机制的了解有利于我们更好地应对未来全球变化下湖泊生态系统的响应,提出更切实有效的湖泊生态系统管理对策. ...
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
... [21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
Flickering gives early warning signals of a critical transition to a eutrophic lake state
2
2012
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
A palaeolimnological perspective to understand regime-shift dynamics in two Yangtze-basin lakes
1
2019
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
Generic indicators for loss of resilience before a tipping point leading to population collapse
1
2012
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
Recovery rates reflect distance to a tipping point in a living system
1
2012
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
Predicting ecosystem state changes in shallow lakes using an aquatic ecosystem model: Lake Hinge, Denmark, an example
1
2020
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
A sediment resuspension and water quality model of lake Okeechobee
1997
A review of researches on the lake numerical modeling
1
2013
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
湖泊数值模拟研究现状综述
1
2013
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
1
2003
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
Diatom flickering prior to regime shift
1
2013
... 目前对浅水湖泊稳态转换研究的数据主要来源于野外观测[22,23]、实验室实验[24,25]和模型模拟[21,26~28],研究人员可以从野外获取观测数据,通过分析这些数据来得到稳态转换存在的证据[29].但是,自然生态系统通常异常复杂,某些现象出现的原因往往存在多解性,研究结果通常争议较大[30].实验室实验的优势在于可以对实验系统进行人为设计,针对特定问题观测特定生态系统过程.但是,由于实验系统往往过于简单,在较小生态系统中稳态转换研究的结果并不一定完全适用于自然生态系统[21].除了现场观测和实验外,模型是建立稳态转换理论和探究湖泊稳态转换机制不可或缺的手段.模型模拟的优势在于方便快捷,是迅速验证假设的最有效手段.但是其可靠性以及适用性常常遭受质疑.因此,在稳态转换的研究中,通常3种手段互相结合,交叉验证得到可靠的结果. ...
Identification of regime shifts and their potential drivers in the shallow eutrophic Lake Yilong, Southwest China
1
2020
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
Regime shifts revealed by paleoecological records in Lake Taibai's ecosystem in the middle and lower Yangtze River Basin during the last century
1
2016
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
古生态记录揭示的长江中下游太白湖生态系统稳态转换过程
1
2016
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
Tracking ecosystem regime shifts in Lake Xijiu(Taihu Basin) based on chironomid sub-fossil assemblages
1
2020
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
太湖流域西氿摇蚊亚化石群落对湖泊生态系统稳态转换的响应
1
2020
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
Recent progress and perspectives in paleoenvironmental and paleoclimatic research based on chironomidae (Diptera)
1
2016
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
基于摇蚊的古环境和古气候国内外研究进展与展望
1
2016
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
Sedimentary diatom records reveal the succession of ecosystem in Lake Xihu,Dali over the past 50 years
1
2020
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
硅藻群落指示的近50年来大理西湖湖泊生态系统演变规律
1
2020
... 针对湖泊生态系统稳态转换的研究,国际上已经相继开展了实验室实验、野外观测研究及模型模拟.在中国的长江中下游地区、云南等重要湖泊也通过多种手段对湖泊生态系统稳态转换过程、机制等进行了研究[31,32].然而,相关研究主要针对典型湖泊以及单一影响要素,如摇蚊群落[33,34]和硅藻群落[35].未来人类活动和气候变化将以多种不同方式,通过湖泊不同过程如营养盐的富集、循环效率、生物富集等的变化来影响湖泊生态系统稳态转换.因此,亟须一项关于全球变化背景下湖泊稳态转换的综合性研究,为湖泊的综合管理提供科学支撑.本文将通过磷动力学模型模拟湖泊生态系统稳态转换过程,结合相关的野外调查、模型模拟以及实验室实验结果,在湖泊稳态转换理论框架下,分析各环境要素对湖泊稳态转换主要特征的影响,为湖泊管理提供科学参考. ...
Mechanisms and research methods of phosphorus migration and transformation across sediment-water interface
1
2019
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
湖泊沉积物—水界面磷的迁移转化机制与定量研究方法
1
2019
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Nutrient cycling flux of Lake Dianchi: A three-dimensional water quality modelling approach
1
2017
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
湖泊营养盐通量平衡的三维数值模拟
1
2017
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Management of eutrophication for lakes subject to potentially irreversible change
1
1999
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Seasonal algal blooms support sediment release of phosphorus via positive feedback in a eutrophic lake: Insights from a nutrient flux tracking modeling
1
2020
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Analyzing the mutual feedbacks between lake pollution and human behaviour in a mathematical social-ecological model
1
2020
... 在湖泊磷循环中,磷可通过大气沉降和地表径流等途径流入湖泊,或在沉积物、上覆水和生物体间相互转化,或通过生物捕获和出湖径流流出湖泊[36].湖泊中影响水体磷浓度主要有3个方面:人类活动和气候变化所导致的外源磷负荷输入增多[10]以及改变内源磷释放效率[11,12],还有出湖径流携带走大量的磷[37].本文选用以下磷动力学模型公式来模拟湖泊中的磷循环[38],该模型已被广泛应用在湖泊生态系统稳态转换的研究中[22,39,40]. ...
Analysis of telemedicine promotion strategy based on Vensim simulation
1
2020
... 模型参数对应的湖泊属性如图1a所示,其中时间t在模型中实际上是指步长.建模软件是Vensim PLE 6.0.0.1[41,42].在Vensim中构建好模型后,反复调整α的数值,当α设置为分段函数时: ...
基于Vensim仿真的远程医疗推广策略分析
1
2020
... 模型参数对应的湖泊属性如图1a所示,其中时间t在模型中实际上是指步长.建模软件是Vensim PLE 6.0.0.1[41,42].在Vensim中构建好模型后,反复调整α的数值,当α设置为分段函数时: ...
Development of green transportation based on Vensim simulation
1
2016
... 模型参数对应的湖泊属性如图1a所示,其中时间t在模型中实际上是指步长.建模软件是Vensim PLE 6.0.0.1[41,42].在Vensim中构建好模型后,反复调整α的数值,当α设置为分段函数时: ...
基于Vensim仿真的城市绿色交通发展问题研究
1
2016
... 模型参数对应的湖泊属性如图1a所示,其中时间t在模型中实际上是指步长.建模软件是Vensim PLE 6.0.0.1[41,42].在Vensim中构建好模型后,反复调整α的数值,当α设置为分段函数时: ...
Catastrophic shifts in ecosystems
1
2001
... 图1b展示了模型模拟的浅水湖泊的稳态转换过程.在驱动力梯度上系统状态存在2个突变点,即F1和F2,当驱动力越过临界点时,生态系统状态会发生临界转换.在F1和F2之间,平衡曲线向后折叠,因此,该模型又被称为折叠交叉模型[43].在此条件区间内系统存在3个平衡.上分支和下分支曲线都是实线,代表稳定的平衡,中间虚线部分是指不稳定的平衡,代表了上分支和下分支2个稳定状态吸引域的边界.图1中的灰色虚线箭头是指如果系统不处于平衡状态时(即不在曲线上时),系统移动的方向.随着驱动力的增加,系统的稳态没有发生改变,仍在上分支,但是系统的吸引域或弹性会随着驱动力的增加而减小,当系统接近折叠灾变点F2时,驱动力的微小变化都可能导致系统突变为下分支的稳态.但是系统若想从下分支回到上分支,驱动力减小到F2大小时是远远不够的,要继续减小,当减小到F1时,系统才能从下分支突变到上分支.2个稳态相互转换发生在不同的驱动力突变点下,这种情况就称为滞后效应[20].具体到湖泊生态系统,图2表现了水体总磷浓度在外源驱动下出现的清水态和浊水态之间稳态转换现象和滞后效应.磷动力学模型结果如图2a(P-t图)和图2b(P-α图)所示.图2a中存在突变时间A和B,其中A是指湖水磷浓度由低浓度突变到高浓度的突变时间,B是指湖水磷浓度从高浓度降回低浓度时的突变时间.图2b中同样存在2个突变点F1和F2,突变点F2是指改变参数后,清水稳态突变为浊水稳态时外源磷输入的阈值大小,突变点F1是指由浊水稳态修复回清水稳态时的外源磷输入值.X(F1,F2)是指稳态间转变的滞后时长;稳态大小是指湖泊由清水态或者浊水态时的P浓度. ...
Eutrophication in cold-water lakes driven by combined effects of climate change and human activities
1
2017
... 气候变化和人类活动是影响湖泊磷浓度变化的两大因素,其中人类活动往往在湖泊水体磷的贡献上起主导作用.近年来,人类活动所导致的外源磷负荷输入增多和沉积物中磷释放增加是加剧湖泊富营养化的主要驱动因素[44,45].人类活动对湖水磷浓度的影响主要表现在改变入湖径流水质、水位、生物扰动强度、微生物和换水周期等方面.而气候变化则主要表现在降雨模式改变、风浪扰动、水温和光照强度等因素的变化上.气候变化的影响,往往会叠加在人类活动之上进一步加剧湖泊的磷污染,增强湖泊富营养化.气候变化和人类活动对湖水磷浓度的具体影响因素所造成的结果及机理分析如表3所示. ...
气候变化与人类活动双重驱动的冷水湖泊富营养化
1
2017
... 气候变化和人类活动是影响湖泊磷浓度变化的两大因素,其中人类活动往往在湖泊水体磷的贡献上起主导作用.近年来,人类活动所导致的外源磷负荷输入增多和沉积物中磷释放增加是加剧湖泊富营养化的主要驱动因素[44,45].人类活动对湖水磷浓度的影响主要表现在改变入湖径流水质、水位、生物扰动强度、微生物和换水周期等方面.而气候变化则主要表现在降雨模式改变、风浪扰动、水温和光照强度等因素的变化上.气候变化的影响,往往会叠加在人类活动之上进一步加剧湖泊的磷污染,增强湖泊富营养化.气候变化和人类活动对湖水磷浓度的具体影响因素所造成的结果及机理分析如表3所示. ...
Assessing and managing nutrient-enhanced eutrophication in estuarine and coastal waters: Interactive effects of human and climatic perturbations
1
2006
... 气候变化和人类活动是影响湖泊磷浓度变化的两大因素,其中人类活动往往在湖泊水体磷的贡献上起主导作用.近年来,人类活动所导致的外源磷负荷输入增多和沉积物中磷释放增加是加剧湖泊富营养化的主要驱动因素[44,45].人类活动对湖水磷浓度的影响主要表现在改变入湖径流水质、水位、生物扰动强度、微生物和换水周期等方面.而气候变化则主要表现在降雨模式改变、风浪扰动、水温和光照强度等因素的变化上.气候变化的影响,往往会叠加在人类活动之上进一步加剧湖泊的磷污染,增强湖泊富营养化.气候变化和人类活动对湖水磷浓度的具体影响因素所造成的结果及机理分析如表3所示. ...
Interaction of climate change and eutrophication
2
2010
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Allied attack: Climate change and eutrophication
1
2011
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Dynamic behavior of suspended sediment concentrations in a shallow lake perturbed by episodic wind events
1
1990
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
The dynamic mechanism of sediment suspension in Taihu Lake and the conceptual model of endogenous release
1
2003
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
太湖沉积物悬浮的动力机制及内源释放的概念性模式
1
2003
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Phosphorus release from lake sediments: Effects of pH, temperature and dissolved oxygen
1
2013
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Temperature dependence of slow adsorption and desorption kinetics of organic compounds in sediments
1997
The role of microorganisms in mobilization and fixation of phosphorus in sediments
1
1993
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Sediment phosphorus release at beaver reservoir, Northwest Arkansas, USA, 2002-2003: A preliminary investigation
1
2007
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Simulated study on phosphorus release of Chao Lake sediment
1
2002
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
巢湖底泥磷的释放模拟实验研究
1
2002
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Study on effects of light on phosphorus release and phosphorus form change in lake sediments
1
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
光照对湖泊沉积物磷释放及磷形态变化的影响研究
1
2004
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Effects of pH and salinity on phosphorus sorption and desorption in the surface sediments of the main stream of the Haihe River
1
2009
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
pH和盐度对海河干流表层沉积物吸附解吸磷(P)的影响
1
2009
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Phosphorus fractions and the effect of pH on the phosphorus release of the sediments from different trophic areas in Taihu Lake, China
1
2006
... 人类活动,如农业和工业污水排放的增加会增加入湖营养盐负荷,导致湖泊外力驱动的加强,对应于模型α参数的增加.同时,乱砍乱伐和过度放牧等会造成森林等植被面积减少,林草地退化和土地荒漠化[57],水土流失严重,最终导致入湖营养物质增多,与模型中磷输入速率α上升速率增大相对应.因此,人类活动的加强会导致湖泊清水态—浊水态的突变时间提前(突变时间A提前).发生稳态转换时湖泊所能承受的外源负荷阈值增大(突变点F2增大),稳态间转变的滞后时长增加[X(F1,F2)增大].稳态不变,说明入湖径流水质差所导致的α上升斜率增大不会进一步提高湖泊富营养化时的污染程度. ...
Remarkable efficiency of phosphate removal: Ferrate(VI)-induced in situ sorption on core-shell nanoparticles
1
2016
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Ecological water treatment system for removal of phosphorus and nitrogen from polluted water biological application
1
1997
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
The effect of biomanipulation on phosphorus exchange between sediment and water in shallow, tropical Huizhou West Lake, China
1
2017
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Advances in impacts of climate change on surface water quality
1
2012
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
全球气候变化对地表水环境质量影响研究进展
1
2012
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Temporal and spatial variability of phytoplankton in Lake Poyang: The largest freshwater lake in China
2
2013
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
Phosphorus release from resuspended sediment in the shallow and wind-exposed Lake Arres?, Denmark
2
1992
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
Changes to flow and turbulence caused by different concentrations of fish in a circular tank
2
2015
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
Effects of common carp on aquatic ecosystems 80 years after "Carp as a Dominant": Ecological insights for fisheries management
2009
The influence of Tubificid worms bioturbation on the exchange of phosphorus across sediment-water interface in lakes
2010
水丝蚓(Tubificid worms)扰动对磷在湖泊沉积物—水界面迁移的影响
2010
Effects of deposit-feeding tubificid worms and filter-feeding bivalves on benthic-pelagic coupling: Implications for the restoration of eutrophic shallow lakes
2014
Mesocosm experiment reveals a strong positive effect of snail presence on macrophyte growth, resulting from control of epiphyton and nuisance filamentous algae: Implications for shallow lake management
2
2020
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
Effects of light, temperature and algae on phosphorus release from sediment
2
1992
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
光照、温度和藻类对底泥释放磷的影响
2
1992
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
Simulation experiment study on the effect of microorganism on phosphorus release from Taihu Lake sediments
2
2006
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
微生物作用对太湖沉积物磷释放影响的模拟实验研究
2
2006
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
The spatial-temporal characteristics of eutrophication in Poyang Lake and its relationship with the water level
2
2019
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 人类活动,如筑坝对湖泊的换水周期有较大的影响[71].由于s参数在模型中是指磷的输出速率,s增大意味着磷的输出量增加.因此,换水周期短与s增大相对应.若湖泊处于清水态,换水周期变短,会推后湖泊清水态—浊水态的突变时间(突变时间A推后),并且在发生稳态转换时所能承受的外源负荷增多(突变点F2增大),突变到浊水态时的富营养化程度降低(稳态减小).而若当湖泊处于浊水态时,换水周期短能够使湖泊浊水态—清水态的修复时间提前(突变点B提前),并且在发生稳态转换时外源磷的削减程度降低(恢复点F1增大),稳态间转变的滞后时长减短[X(F1,F2)减小],这些变化都有利于湖泊修复和治理.相反,较长的换水周期,意味着磷的滞留时间长,湖泊磷浓度增大,给湖泊带来的影响则相反,不利于湖泊管理和修复. ...
鄱阳湖富营养化时空变化特征及其与水位的关系
2
2019
... Influencing factors and mechanism of phosphorus concentration in lake water
Table 3| 分类 | 影响因素 | 结果 | 原因 | 参考文献 |
|---|
| 气候 变化 | 降雨模式改变 | 增加水体营养物质 | 增加营养负荷面源;改变营养物质的入湖通量和水力停留时间 | [46,47] |
| 风浪扰动 | 促进底泥磷释放 | 增加沉积物的再悬浮量;抑制沉水植物生长 | [48,49] |
| 水温 | 水温升高,促进底泥磷释放 | 水温升高能促进底泥中Fe-P和Ca-P的释放 | [50~52] |
| 溶解氧 浓度 | 厌氧条件下,促进底泥磷释放 | 难溶的(Fe(OH)3)x转化为可溶性的 Fe(OH)2,使 PO脱离沉积物进入间隙水 | [53,54] |
| 光照强度 | 照度增强抑制底泥磷释放 | 促进底栖藻类生长,形成阻碍底泥磷释放的屏障 | [55] |
| pH | 酸性和碱性条件下,促进底泥磷释放;中性条件下,抑制底泥磷释放 | 酸性条件下,促进Ca-P释放;碱性条件下,促进Fe-P释放;中性条件下,有利于底泥的磷吸附 | [56~58] |
| 沉水植物 | 吸收水体和沉积物中的营养盐;抑制底泥磷释放 | 抑制底泥再悬浮;提高湖水—底泥界面的氧化还原电位,从而抑制底泥中Fe-P释放 | [59,60] |
| 人类 活动 | 入湖径流水质 | 水质差会导致湖泊磷浓度增多 | 携带大量营养盐进入湖泊 | [61] |
| 水位 | 水位升高,湖泊富营养化程度降低;水位下降,湖泊富营养化程度增加 | 水位升高会稀释湖水营养物质,水位降低会加剧底泥的再悬浮和磷的释放 | [62,63] |
| 生物扰动 | 经济鱼类(鲤、鲢和鳙等)和一些底栖动物(水丝蚓)促进底泥磷释放;一些螺类(如田螺、石螺等)和双壳类(如河蚌、河岘等)抑制底泥磷释放 | 鱼类密度的增加及其摄食活动会促进底泥的再悬浮;双壳类可以降低徜水生境中的悬浮物浓度,提高透明度;螺类能促进沉水植物的生长,降低浮游植物的密度 | [64~68] |
| 微生物 | 有微生物促进底泥磷释放;无微生物抑制底泥磷释放 | 微生物可通过微生物作用将沉积物中的不溶性磷转化为可溶性磷 | [69,70] |
| 换水周期 | 换水周期短,湖泊富营养化程度降低;换水周期长,富营养化程度加深 | 换水周期短,导致湖泊大量营养盐流出 | [71] |
<strong>4.1</strong> 气候变化对湖泊生态系统稳态转换的影响全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
... 人类活动,如筑坝对湖泊的换水周期有较大的影响[71].由于s参数在模型中是指磷的输出速率,s增大意味着磷的输出量增加.因此,换水周期短与s增大相对应.若湖泊处于清水态,换水周期变短,会推后湖泊清水态—浊水态的突变时间(突变时间A推后),并且在发生稳态转换时所能承受的外源负荷增多(突变点F2增大),突变到浊水态时的富营养化程度降低(稳态减小).而若当湖泊处于浊水态时,换水周期短能够使湖泊浊水态—清水态的修复时间提前(突变点B提前),并且在发生稳态转换时外源磷的削减程度降低(恢复点F1增大),稳态间转变的滞后时长减短[X(F1,F2)减小],这些变化都有利于湖泊修复和治理.相反,较长的换水周期,意味着磷的滞留时间长,湖泊磷浓度增大,给湖泊带来的影响则相反,不利于湖泊管理和修复. ...
Study on the spatiotemporal evolution of temperature and precipitation in China from 1951 to 2018
1
2020
... 全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
基于自然分区的1960—2018年中国气候变化特征
1
2020
... 全球气候变化给湖泊带来很大的影响,加剧湖泊的富营养化进程[46,72].气候变化带来的降雨模式改变、风浪变化以及间接对湖泊带来的影响,如通过影响沉水植物、水体温度、光照强度、溶解氧浓度和pH来改变湖泊水体中的磷浓度,最终都会导致湖泊生态系统稳态转换模式的改变(表4). ...
Research advance of changing biogenic substance cycling in river systems by damming
1
2018
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
河流筑坝对生源物质循环的改变研究进展
1
2018
... 一些人类活动,如向湖泊中投入大量经济鱼类(如鲤、鲢和鳙等)和一些螺类(如田螺和石螺等)和双壳类的底栖动物(如河蚌和河岘等)所造成的生物扰动都会对湖水磷浓度造成一定的影响[64~68].微生物可通过微生物作用将沉积物中的不溶性磷转化成可溶性磷,而人为的投入抑菌剂就会对微生物活性造成影响,从而间接影响湖泊磷浓度[69,70].另外,如兴建蓄水库、蓄洪区、堤坝和水闸等人类活动[73]会对湖泊水位造成影响.而水位的升高和降低都能够影响湖泊中的磷浓度[62,63]. ...
A general one-dimensional vertical ecosystem model of Lake Shira (Russia, Khakasia): Description, parametrization and analysis
1
2010
... (3)本研究从气候变化和人类活动这两方面入手来探究湖泊磷浓度的影响因素,按照影响湖泊稳态转换过程的关键要素对气候变化和人类活动进行了分类探讨,但是二者的交互作用在本文中没有详细讨论.此外,本研究的结果并不是针对具体的湖泊,而是给出了一个宏观的框架;通过稳态转换模型和试验研究、自然生态系统研究相结合,联系了全球变化主要因子与稳态转换模型的主要参数,讨论了各因子驱动下湖泊稳态转换的特征.具体湖泊的情况要综合考虑各种要素的交互作用影响,已经超出了本研究采用的简单模型的能力范围,需要采用更加复杂的稳态转换模型,如PCLake等[74].研究建议将稳态转换理论引入湖泊生态管理的科学支撑研究体系中,这样能够更好的为湖泊修复和治理提供参考意义. ...




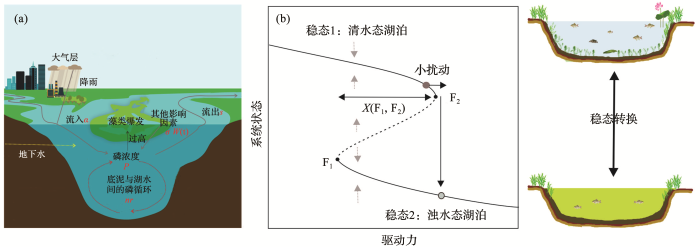
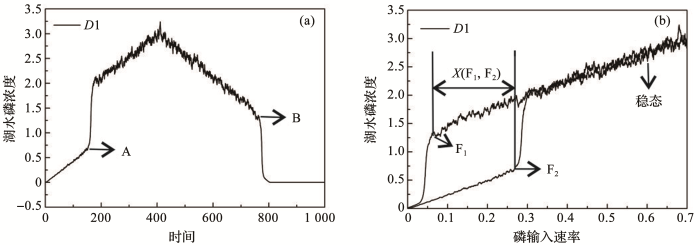
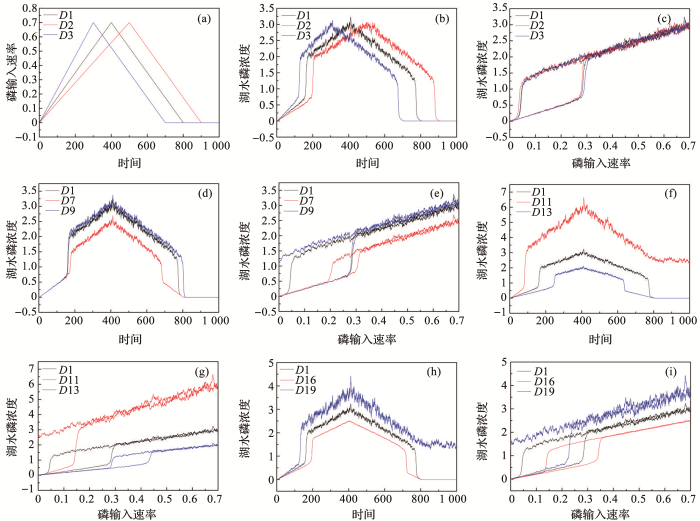

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}