



1 引 言
因此本文围绕钡的地球化学特征、源汇循环及分馏机制展开系统综述,并探讨其在古海洋环境示踪中的应用潜力。通过梳理最新研究进展,整合矿物—流体—熔体体系的微观分馏机制,为钡同位素分馏机制研究提供系统性框架,助力深化海洋演化历史与生态变迁的认知。
2 钡地球化学特征
钡在地球圈层中的含量与同位素分布呈现出显著的分异特征。其含量自地核至地壳递增,而同位素表现出“源轻汇重”的特点。在岩石圈和水圈中,因地质、岩性与环境的差异,钡含量及同位素值变化多样。在海洋环境中钡垂向分布受陆源、热液和生物物质输入,生物地球化学过程,及水团垂向运动、沉积物垂向再悬浮等物理过程影响;横向分布则因不同海域物质输入强度、生物地球化学过程区域特性,以及水团水平运动、沉积物水平再悬浮等物理过程的区域差异而变化。
2.1 钡含量及分布
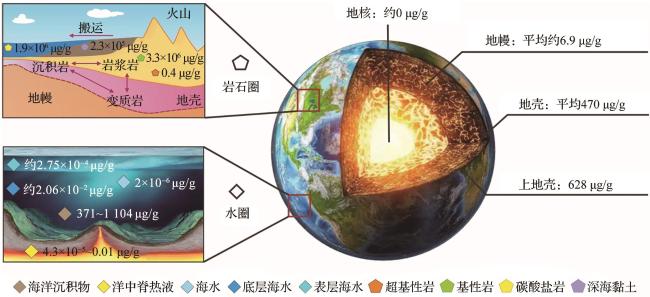
2.2 钡同位素分布
(x=137,138)
| 基本属性 | 130Ba | 132Ba | 134Ba | 135Ba | 136Ba | 137Ba | 138Ba |
|---|---|---|---|---|---|---|---|
| 原子质量 | 129.906 | 131.905 | 133.905 | 134.906 | 135.905 | 136.906 | 137.905 |
| 丰度/% | 0.106 | 0.101 | 2.417 | 6.592 | 7.854 | 11.232 | 71.698 |
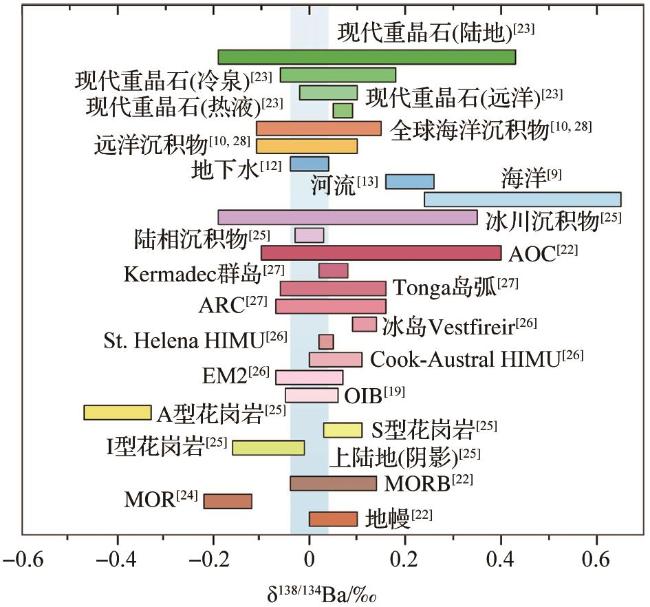
3 海洋中钡的来源
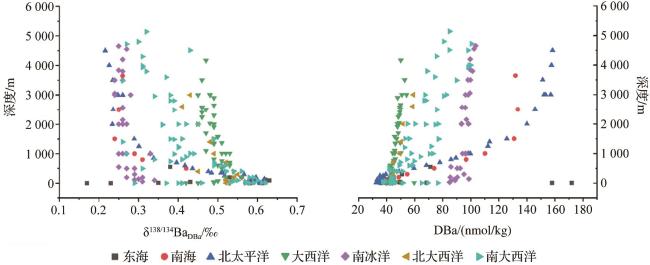
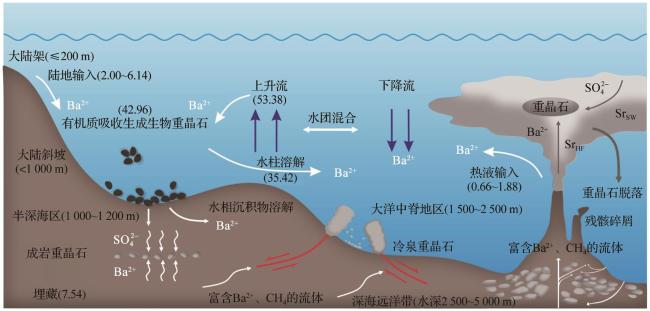
海洋中的钡主要来源于陆地输入和海底热液活动[1],此外,海洋生物也是来源之一(图4)。依据表2中的数据估算,陆源输入在全球海洋钡通量占比为56.65%±35.45%,热液输入为7.5%±5.5%,生物输入为30%±10%;此外仍有约5.85%的钡来源尚未明确,需进一步研究。且早期对全球海洋钡通量的估算为18.1 Gmol/a,不过该估算仅涵盖部分河流与热液喷口钡通量,难以反映实际情况[38],对全球海洋沉积物钍(Th)—归一化质量积累率(Thorium-normalized Mass Accumulation Rate,AR)汇编分析后,发现水深超1 000 m处,全球海洋钡通量至少为(19±10) Gmol/a[39],此外,钡在海洋的滞留时间从原来的约8 500年缩短为3 500~5 000年(颗粒钡)[39]。
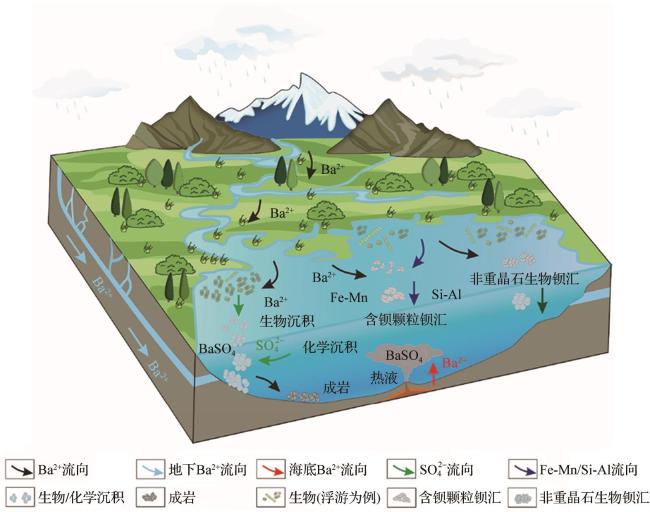
表2 钡来源及通量情况Table 2 Flux situation of barium sources |
| 钡的来源 | 通量 | 影响因素 |
|---|---|---|
| 陆源河流[1,34-36] | 全球平均2.00~6.14 nmol/(cm2·a) | 河流流域的岩石类型、土壤性质、水文条件;河口地区水体混合、吸附—解吸附、近岸陆源输入影响较大、远洋海域影响较小 |
| 三江源和祁连山海洋输出钡为9.03×10-3 Gmol/a | ||
| 黄河Baraw年通量为(0.079±0.028) Gmol/a | ||
| 河口超额钡供应通量约4.5 Gmol/a | ||
| 陆源地下水[1,12] | 0.46 nmol/(cm2·a)(约占河流通量的25%) | 排放体积流量不确定、溶质通量难约束 |
| 全球溶质通量范围为0.078~0.706 Gmol/a | ||
| 热液[1,10-11,17] | 1.00~1.39 nmol/(cm2·a) | 热液活动持续时间、海底深度、溶液温度;重晶石沉淀 |
| 2.40~3.35 Gmol/a | ||
| 2.4~6.8 Gmol/a | ||
| 生物[2,37] | 大西洋NAP站点:约2.585 nmol/(cm2·a) | 海洋生产力、生物群落结构、海域差异和环境钡浓度 |
| 赤道太平洋M站点:约5.388 nmol/(cm2·a) | ||
| 赤道太平洋H站点:约9.801 nmol/(cm2·a) |
3.1 陆源输入
陆源输入是海洋非生物钡的重要来源,河流为主要输送途径。在海洋中,轻钡同位素在真光层与有机质结合,有机质在弱光层被分解,并产生BaSO4过饱和微环境,轻钡同位素会优先吸附并沉淀,形成颗粒钡[18]。陆源河流对海洋钡通量的贡献显著,但数据差异较大,存在诸多不确定性(表2)。全球陆源河流颗粒钡平均通量因样品代表性不足以及未考虑时空变化等因素,存在一定的不确定性[1]。此外,不同区域因岩石类型、风化及降水差异,输送钡的能力不同,导致钡的通量容易出现波动,如三江源和祁连山区域生态较为脆弱,而黄河等流域受人类活动影响[34-35],这些因素都会影响钡通量。此外通过盐度—钡浓度线性外推法(将高盐度区钡浓度的保守混合线外推至零盐度),其理论值(Baeff)与河流实测零盐度钡浓度(Bariv)的差值,乘以河水流量(Q),即(Baeff-Bariv)×Q,获得的河口超额钡通量约为4.5 Gmol/a,约占全球河流供应通量的37%[36],对海洋物质收支影响重大。但由于河口的生态复杂,生物活动、盐度和海平面变化等因素会使超额钡供应通量不稳定,从而增加了海洋钡收支长期预测的难度。
3.2 热液输入
热液钡输入对海洋钡循环影响重大(表2)。Hsieh等[17]的研究表明,全球热液钡排放通量估计值为0.66~0.93 nmol/(cm2·a),其输入占部分大西洋深层水钡含量的3%~9%,影响海水钡同位素组成及分布。Carter等[1]的研究指出,热液喷口对海洋中钡输入贡献为2%~13%,通量为1.00~1.39 nmol/(cm2·a),因重晶石沉淀,有效热液钡通量远低于初始排放通量,且他们均认为通量被高估[1,17]。Zhang等[11]近期给出全球热液钡通量为0.66~1.88 nmol/(cm2·a),刷新了热液的通量记录,整体稍低于河流输入量[2.00~6.14 nmol/(cm2·a)]。而这些差异主要源于研究的区域、时间和方法的不同,且重晶石沉淀过程较为复杂,导致有效通量的估算存在一定的不确定性。
3.3 生物输入
4 海洋中钡的汇聚
海洋中的钡以重晶石为依据,分为重晶石钡汇和非重晶石钡汇,明确其成因与特征意义重大。也可按照生物与非生物区分:生物钡由浮游生物吸收、微生物代谢以及珊瑚矿化等生物活动形成,与生物泵相关,可用于重建古生产力;而非生物钡则由热液活动和化学沉淀等物理化学过程形成,分布受地质构造控制,二者均为海洋钡循环的重要组成部分。本节将综合考虑一同叙述。
4.1 重晶石钡汇
重晶石钡汇主要包括沉积型和成岩型两类,其形成机理不同,鉴定特征各异(表3)。
表3 重晶石成因分类与环境、鉴定特征汇总(据参考文献[1, 41]修改)Table 3 Summary of the genetic classification, environments, and identification characteristics of barite (modified after references [1, 41]) |
| 类型 | 成因分类 | 形成机理 | 形成环境 | 鉴定特征 |
|---|---|---|---|---|
| 沉积型 | 生物 | 表层水体重晶石的过饱和状态由初级生产者分解,并伴生有机质降解,在微环境中触发 | 开阔海洋及浅海陆棚带受光照、温度和营养盐驱动形成生物繁育核心带 | 微晶颗粒,粒径<5 μm,常呈自形微级椭圆体沉淀 |
| 化学 | 海水过饱和条件下Ba2+与SO 自发结合生成微溶硫酸钡沉淀 | 局限海相富Ba2+-SO 水体 | 晶体形态多样,有规则板状、柱状晶体,粒径通常较大 | |
| 成岩型 | I型 | 硫酸盐亏损带富Ba2+流体上涌与海水硫酸盐下渗在硫酸盐—甲烷转换带顶部附近交汇 | 沉积物孔隙水区域 | 晶体较大(>20 μm),扁平板柱状,以铁锰结核、透镜体和纹层等形式分布于沉积物中 |
| II型 | 冷泉区富Ba2+流体与海水SO 垂向交汇低温成矿形成自生硫酸钡 | 冷泉附近,有冷泉流体活动,气液和含甲烷冷溶液 | 晶体较大且不规则,呈脉状、烟囱体、丘状等,如20 m高重晶石丘,重晶石体积占比25%~80% | |
| III型 | 深部富Ba2+热液通过断裂带垂向运移,与海水SO 混合形成硫酸钡 | 洋中脊及弧后盆地等高热液通量区的硫酸盐—硫化物成矿 | 晶体常呈粗大柱状和板状,晶形完整,常伴金属硫化物共生 |
4.1.1 沉积型
沉积型包括生物沉积和化学沉积两种类型。
生物沉积型重晶石的形成与生物活动密切相关(表3),主要分布于海洋水深≤200 m的大陆边缘与大陆架区域(图5)。生物驱动作用包括直接与间接作用: 直接作用方面,微生物(如异足虫和蓝藻)体内可发生Ba2+与SO 反应,形成微晶重晶石颗粒。当生物死亡后,颗粒释放并参与沉积物构建[41]。浮游生物(如Acantharia)的代谢活动或细胞破裂也可能释放钡,并与硫酸根结合成矿[42-43]。此外,放射虫外壳中的天青石溶解可能释放出高浓度的钡,从而触发过饱和沉淀[1]。 间接作用则是通过生物活动改变微环境化学性质来实现的。微生物代谢产物可调节水体的酸碱度,从而影响钡和硫酸根的溶解度;细菌生物膜或沉积物中的富含磷组分优先吸附钡离子,随后硫酸根替代磷,从而形成重晶石[44-45]。在高生产力区域,浮游生物分泌的胞外聚合物(Extracellular Polymeric Substances,EPS)为Ba2+富集提供成核位点[45]。值得注意的是,生物膜对钡同位素存在选择性利用:轻钡同位素因脱水能垒较低,更容易被富集;而有机质降解释放的重钡同位素则改变环境中的同位素分布[42,46](图6)。然而,生物驱动的直接机制仍存在复杂争议[47]。
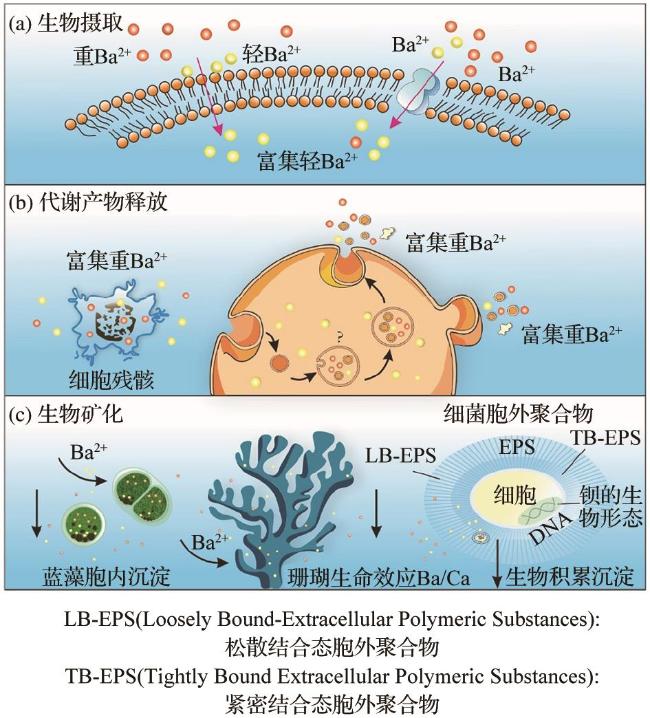
在正常海洋环境的特定区域(如硫酸盐还原区、甲烷生成区及大洋中脊热液喷口处),会产生大量H2S。在富含有机质且有硫酸盐还原菌的海洋沉积物环境中,SO 被还原为H2S。H2S进入缺氧环境,HS-被氧化使SO -增多,进而与Ba2+反应生成重晶石沉淀[49]。
4.1.2 成岩型
成岩型重晶石按照成矿流体来源(孔隙水、冷泉和热液)分为I型、II型和III型[2]。
III型热液型重晶石形成于大洋中脊(图5,水深1 500~2 500 m)、火山活动区等地质活跃带,热液沿岩石裂隙上升至海底,与冷海水或周围岩石孔隙流体剧烈混合,导致温度、压力骤降及化学成分改变,Ba2+与海水中的SO 因溶解度大幅降低而快速沉淀[57],形成重晶石(表3)。与热液活动直接相关,热液为其提供Ba2+来源,沉淀发生于热液与海水、岩石孔隙流体等周围介质的混合区域,温度和压力变化显著影响其形成过程。如密西西比河谷型和爱尔兰型矿床,其重晶石形成分别与白云石化作用、热液活动及深部热液循环等有关[58]。热液流体富集轻钡同位素,形成的热液喷流重晶石钡同位素偏轻,为0.17‰±0.05‰[11]。该类型重晶石常与硫化物和石英等热液矿物共生,且形成的地质环境多具热液蚀变岩石等明显的热液活动痕迹(表3)。
4.2 非重晶石钡汇
5 生物钡重建古生产力
浮游生物、底栖生物及珊瑚等造礁生物的微观分馏机制深刻影响着钡同位素的海洋分布,对海洋及古生产力具有重要的指示意义。
钡作为海洋生产力的示踪指标的前提是与有机碳(Corg)存在耦合关系,海洋中颗粒有机碳(Particulate Organic Carbon,POC)与钡通量强相关,颗粒Corg/Ba值约为200,且随深度减小,反映出有机碳降解与钡吸收的耦合,表明钡的去除与生物过程相连。浮游、底栖生物及珊瑚等造礁生物的微观分馏机制影响钡同位素分布,关联海洋生产力。学者们据此建立了定量关系,如Dymond等[64]用深海沉积物数据估算新生产力,Francois等[65]建立了有机碳与颗粒生物源钡的通量关系,Murray等[66]以Ba/Ti值作为输出生产代理指标。在海洋古生产力重建的研究中,利用生物钡精准指示古生产力,并有效去除非生物钡的干扰,是重要的研究方向,当前主流的研究方法有过剩钡法、重晶石积累速率法、Ba/Ti与Ba/Al元素比值法、综合多元素比值法以及新兴的钡同位素法,但这些方法仍在不断发展和完善中,且各自面临着诸多挑战(表4)。
表4 古生产力重建方法及局限性分析Table 4 Methods and limitations analysis of paleoproductivity reconstruction |
| 重建方法 | 原理 | 局限性 |
|---|---|---|
| 过剩钡(Baexcess)[46,67-70] | 总钡含量减去碎屑铝硅酸盐贡献
式中: 为沉积物中总钡含量; 为沉积物中铝元素含量 | ①生物因素复杂:物种特异性差异,生长阶段及生理状态,死亡后成岩改造; ②环境因素多样:化学条件,环流与水团混合,钡源及迁移复杂; ③分析方法和数据解释不确定:参数关系不明,校准关系适用性受限; ④地质样品干扰剔除不彻底; ⑤假设模型局限性,无法全面适配 |
| 重晶石积累速率 |
式中: (Mass Accumulation Rate)为物质堆积速率; (Dry Bulk Density)为干体积密度,指单位体积干沉积物的质量; 为沉积时间 计算重晶石在沉积物中的积累速率 | |
| Ba/Ti值和Ba/Al值[73] | 假设铝、钛主要源于陆源碎屑且通量稳定,通过计算钡与铝、钛比值,排除陆源碎屑干扰,凸显生物源钡信号 | |
| 综合多元素比值[74] | 分析元素(钡、磷、铝、钛、钙等)比值作为输出生产代理指标 | |
| 钡同位素[10,46,75-76] | 过剩钡和沉积物δ138/134Ba正相关 |
6 非生物钡同位素分馏机制
表5 微观分馏机制汇总Table 5 Summary of microscopic fractionation mechanisms |
| 体系 | 模型及主导因素 | 影响因素 | 数据支撑 | |||
|---|---|---|---|---|---|---|
| 平衡 | 动力学 | 平衡 | 动力学 | 平衡 | 动力学 | |
| 矿物—矿物[77,79] | 质量因素 | 扩散速率、沉淀溶解速率和离子交换速率 | 晶体结构、健长和替代离子 | 温度和浓度 | 重晶石矿物间103lnβ≈0.063[79] | 矿物间扩散β为0.010~0.011[77] |
| 重晶石—流体[68,77-78,80-82] | Ba2+与SO 形成特定配位键,吸附 | 离子交换 | 配位、健长和结构 | 固液比 | 沉淀α precip=0.99968±0.00002,溶解α diss=0.99985±0.00006[77] | 重晶石扩散实验钡β因子在0.010~0.011[77] |
| 毒重石—流体[19,77-78,80] | 溶液Ba2+结构 | 溶液酸碱度和离子种类 | 晶体结构、配位和健长 | 固液比 | 300 K时△138/134Baminerals-fluid=0.094‰[19] | 沉淀过程(α)=0.99993±0.00004[78] |
| 岩浆—热液[83] | 水化作用、Ba2+水合数和水化壳结构 | 结晶速率 | 钡浓度、Al-Si无序,钡浓度 | 矿物结晶和热液流体 | 103lnβ累计平均值=0.0798±0.005‰[83] | — |
| 地幔熔体—流体[19,84-86] | 力常数和健长 | 地幔部分熔融及熔体—地幔流体作用 | 温度、流体盐度和铝指数 | — | 103lnα 流体-熔体落在-0.62‰~-0.14‰内[84] | — |
|
6.1 矿物—矿物
6.2 矿物—流体
(1) 重晶石—流体
(2) 毒重石—流体
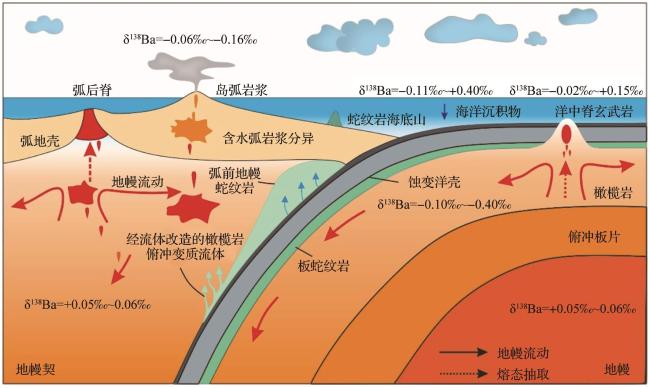
6.3 熔体—流体
熔体—流体间分馏机制研究主要围绕平衡分馏,涵盖岩浆—热液和地幔熔体—流体两大体系,动力学分馏尚处于分馏行为分析阶段。
(1) 岩浆—热液
(2) 地幔熔体—流体
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
7 结语与展望
本文聚焦于钡及其同位素海洋地球化学循环驱动机制,得出以下结论:①钡在地球圈层中的分布呈现显著的“源轻汇重”特征,其含量从地核到地壳递增,同位素组成则表现为岩石圈较轻、水圈较重。这一分异规律为理解钡的跨圈层迁移及海洋循环机制提供了重要依据。②海洋钡循环受陆源、热液及生物过程协同驱动,其中生物端元的代谢释放形成动态源;后随陆源热液端元一同沉降埋藏形成物质汇,其在源与汇间的动态切换,驱动钡循环的持续运转与动态平衡。③阐明钡同位素微观分馏机制受平衡与动力学分馏共同控制,在不同体系中多因素交织,决定其在海洋环境中的分布格局。
当前钡及其同位素在海洋地球化学循环的研究中存在微观分馏机制认知不足、欠缺对多源输入与分馏协同研究、生物—矿物—流体交互机制不明、钡的通量计算不确定性强及古海洋演化模型不完善等局限。未来亟待优化钡通量算法、统一标准并降低空间异质性影响,以实现海洋钡循环的精准量化,同时整合多源数据构建高分辨率古海洋演化模型,深入探究不同地质时期海洋钡循环主控因素及对古环境的响应,利用钡同位素分馏识别古海洋环境变化事件并重建生态系统演变,借助原位微区技术深化生物—矿物—流体交互机制研究,优化古生产力重建方法,为海洋资源勘探与生态保护提供支撑,特别是,油气勘探迈入万米深层领域[90-91],古老烃源岩发育的海洋环境与生产力重建[92-93]、万米深层白云岩优质储层形成机理[94]等,都需要高端的分析测试技术[95-96],因此钡同位素作为地质过程重建的新示踪指标,在未来一定能发挥重要作用。