Effects of non-rainfall water inputs on ecosystem functions
2
2017
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
... [1,17]等. ...
Measurements and ecological implications of nonrainfall water in desert ecosystems—a review
1
2019
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
Relationship between dew presence and bassia dasyphylla plant growth
1
2012
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
The role of dew in negev desert plants
2015
Convergent vegetation fog and dew water use in the Namib Desert
2
2019
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
... [6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
The influence of biological soil crusts on dew deposition in Gurbantunggut Desert, Northwestern China
3
2009
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
... 不同BSCs类型显著影响吸湿凝结水的形成[7,36-37].研究表明,与裸地相比,BSCs表层吸湿凝结水形成量更高,并且与蓝藻结皮相比,由地衣和苔藓组成的BSCs能更有效地形成吸湿凝结水[36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... [7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
Dew formation on the surface of biological soil crusts in central European sand ecosystems
5
2012
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
... 易产生误差
[8, 20, 41] | | 涡度相关法 | 能量平衡 | 代表区域广泛 | 易漏掉不确定水源, ...
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 吸湿凝结水的生成量和持续时间在很大程度上取决于生态系统中的天气条件、微生境和凝结面特征[92].BSCs的发育改变了地表微生境和凝结面特征,通过地表微地形的变化,BSCs增加了其吸收的表面积[65],降低了表层风速;与流沙相比,BSCs的热容量较低,日间变暖和夜间降温速度都较快,增加了昼夜温差;较高的细物质和有机物含量使得BSCs容重降低,持水力增加,黏粉粒含量增加;BSCs具有较高的孔隙率和胞外多糖,因此具有较高的水容量,吸湿凝结水的形成使得地表大气达到饱和的时间延迟,导致大气水汽向着地表转移,也就是向水容量较高的BSCs地表土壤孔隙和胞外多糖中转移[8];这些都有利于吸湿凝结水在其表面的形成.另外,BSCs中生物的形成有利于吸收水分,藓类结皮能通过假根和丝状体等吸收吸湿凝结水[93];虽然齿肋赤藓Syntrichia caninervis Mitt的假根系统(rhizoids)不具有吸收水分的作用,其叶片顶端的白色芒尖有利于凝结水的形成,叶片顶端的芒尖分布有从纳米、微米到厘米等不同尺度的水分收集与传输系统(凹槽和疣状突起),具有采集和运输水分的多种模式,分别将空气中的水分子形成水核、水膜和水滴,然后将这些水分通过毛细管作用迅速运输到芒尖底部的叶面,被叶片吸收[94]. ...
The effect of dew and fog on lithic lichens along an altitudinal gradient in the Negev Desert
4
2013
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Non-rainfall water input determines lichen and cyanobacteria zonation on limestone bedrock in the Negev Highlands
2017
Assessing the likelihood of the soil surface to condense vapour: the Negev experience
1
2020
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
Foggy days and dry nights determine crown-level water balance in a seasonal tropical montane cloud forest
1
2014
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
Variation characteristics of non-rainfall water and its contribution to crop water requirements in China's summer monsoon transition zone
1
2019
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
The incidence and implications of clouds for cloud forest plant water relations
1
2013
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
Foliar water uptake: processes, pathways, and integration into plant water budgets
1
2019
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
Diffuse light and wetting differentially affect tropical tree leaf photosynthesis
1
2020
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
Inferring foliar water uptake using stable isotopes of water
1
2017
... 吸湿凝结水是指近地表附近的气态水分,在合适的气象条件下,通过凝结和吸湿作用,附着在地表土壤颗粒和相关生物体表面的自然现象.吸湿凝结水一方面直接为一些动植物提供水源,另一方面通过水分气—液态转换之间的热量释放,在一定程度上缓解了植物受到的干旱胁迫.吸湿凝结水的生态功能在水分为主要限制因子的荒漠地区显得尤为重要,作为一种经常发生的气象现象,其数量可能超过年降雨量而成为荒漠生态系统稳定的主要水源[1-3],对荒漠地区植物[4-6]、生物土壤结皮(Biological Soil Crusts, BSCs)[7-8]、小动物和微生物[9-11]等的生存起着不可替代的作用.吸湿凝结水也决定了干旱期间水分和能量流的量和生态过程[12-13],对土壤—植物相互关系具有重要作用[6,14],影响植物的叶片吸水[15-16],增加光合,降低蒸腾[1,17]等. ...
1
2017
... BSCs是指由隐花植物(非维管植物)如蓝藻、绿藻、地衣、藓类和土壤中微生物以及相关的其他生物体,通过菌丝体、假根和多聚糖分泌物等与土壤表层微小颗粒胶结形成的十分复杂的复合体,作为干旱、半干旱荒漠地表景观的重要组成,其存在是区域生态系统健康的重要标志之一[18].BSCs个体矮小,需水量较少,一般生长在维管束植物之间的裸地表层,具有非常强的极端干旱环境适应能力,长时间无降雨条件下仍然能够存活.结合吸湿凝结水数量少、频率高、表层化的生成特点,相较其他植物,BSCs对其生成更为敏感,一些学者认为,吸湿凝结水是荒漠区BSCs生存的主要水分来源.BSCs作为一种微小的生态系统,具有简单、反应迅速、个体小、具代表性和普遍性等模式系统的特点,可以将其作为一种模式系统来研究复杂的生态学问题[19].因此,研究BSCs与吸湿凝结水的关系,能够更为系统全面地揭示荒漠生态系统吸湿凝结水的生态功能,为更好地理解全球变化背景下旱地生态系统的变化过程提供重要借鉴.有关凝结水的研究国内外起步较早,以Dew为主题词的检索结果表明,近50年来相关文献呈指数上升,从20世纪70年代的几十篇到2020年的2 000余篇.以“Dew”和“Biological soil crusts”为主题词的检索结果显示,关于BSCs与吸湿凝结水形成关系的研究在2010年之前每年仅有1~2篇,之后逐渐增加,2018年达到最高值,但数量仍然很有限(图1).吸湿凝结水生成量少、测量困难,受边界条件影响大、形成过程复杂等,都成为阻碍其研究进展的重要原因. ...
1
2017
... BSCs是指由隐花植物(非维管植物)如蓝藻、绿藻、地衣、藓类和土壤中微生物以及相关的其他生物体,通过菌丝体、假根和多聚糖分泌物等与土壤表层微小颗粒胶结形成的十分复杂的复合体,作为干旱、半干旱荒漠地表景观的重要组成,其存在是区域生态系统健康的重要标志之一[18].BSCs个体矮小,需水量较少,一般生长在维管束植物之间的裸地表层,具有非常强的极端干旱环境适应能力,长时间无降雨条件下仍然能够存活.结合吸湿凝结水数量少、频率高、表层化的生成特点,相较其他植物,BSCs对其生成更为敏感,一些学者认为,吸湿凝结水是荒漠区BSCs生存的主要水分来源.BSCs作为一种微小的生态系统,具有简单、反应迅速、个体小、具代表性和普遍性等模式系统的特点,可以将其作为一种模式系统来研究复杂的生态学问题[19].因此,研究BSCs与吸湿凝结水的关系,能够更为系统全面地揭示荒漠生态系统吸湿凝结水的生态功能,为更好地理解全球变化背景下旱地生态系统的变化过程提供重要借鉴.有关凝结水的研究国内外起步较早,以Dew为主题词的检索结果表明,近50年来相关文献呈指数上升,从20世纪70年代的几十篇到2020年的2 000余篇.以“Dew”和“Biological soil crusts”为主题词的检索结果显示,关于BSCs与吸湿凝结水形成关系的研究在2010年之前每年仅有1~2篇,之后逐渐增加,2018年达到最高值,但数量仍然很有限(图1).吸湿凝结水生成量少、测量困难,受边界条件影响大、形成过程复杂等,都成为阻碍其研究进展的重要原因. ...
Biological soil crusts (biocrusts) as a model system in community, landscape and ecosystem ecology
2
2014
... BSCs是指由隐花植物(非维管植物)如蓝藻、绿藻、地衣、藓类和土壤中微生物以及相关的其他生物体,通过菌丝体、假根和多聚糖分泌物等与土壤表层微小颗粒胶结形成的十分复杂的复合体,作为干旱、半干旱荒漠地表景观的重要组成,其存在是区域生态系统健康的重要标志之一[18].BSCs个体矮小,需水量较少,一般生长在维管束植物之间的裸地表层,具有非常强的极端干旱环境适应能力,长时间无降雨条件下仍然能够存活.结合吸湿凝结水数量少、频率高、表层化的生成特点,相较其他植物,BSCs对其生成更为敏感,一些学者认为,吸湿凝结水是荒漠区BSCs生存的主要水分来源.BSCs作为一种微小的生态系统,具有简单、反应迅速、个体小、具代表性和普遍性等模式系统的特点,可以将其作为一种模式系统来研究复杂的生态学问题[19].因此,研究BSCs与吸湿凝结水的关系,能够更为系统全面地揭示荒漠生态系统吸湿凝结水的生态功能,为更好地理解全球变化背景下旱地生态系统的变化过程提供重要借鉴.有关凝结水的研究国内外起步较早,以Dew为主题词的检索结果表明,近50年来相关文献呈指数上升,从20世纪70年代的几十篇到2020年的2 000余篇.以“Dew”和“Biological soil crusts”为主题词的检索结果显示,关于BSCs与吸湿凝结水形成关系的研究在2010年之前每年仅有1~2篇,之后逐渐增加,2018年达到最高值,但数量仍然很有限(图1).吸湿凝结水生成量少、测量困难,受边界条件影响大、形成过程复杂等,都成为阻碍其研究进展的重要原因. ...
... BSCs位于土壤表层,对环境的敏感性高,组成简单,易于控制,是研究旱地气候变化和生态恢复的模式生态系统,其对气候变化的响应也受到了广泛关注[19].水分是影响BSCs生长发育最重要的环境因子,没有水,其他因子都不会起作用.非降雨水作为可以被BSCs利用的一种持续水资源,在未来全球气候变化背景下,其对BSCs的水碳平衡将产生重要影响,但目前相关研究还比较薄弱.Mónica等[84]在西班牙中部和东南部的研究证明了非降雨水在BSCs应对气候变化中的重要性,并强调变暖引起的非降雨水分输入改变对BSCS碳平衡的影响比降雨更大.Baldauf 等[97]成功地将一个基于过程的模型参数化,量化了大气CO2浓度、气温、降雨量和相对湿度变化对西班牙地中海沿岸地衣结皮Diploschistes diacapsis (Ach.) Lumbsch光合活性和盖度的影响,结果表明,单纯变暖会增强光合作用从而增加结皮盖度,变暖引起的结皮盖度降低主要来源于同时发生的相对湿度的降低和非降雨水分输入的减少.该研究认为在未来全球气候变化背景下,影响地衣结皮Diploschistes diacapsis (Ach.) Lumbsch存活的关键因子是相对湿度的降低,而不是气温的增加,大气CO2浓度升高的作用也可以被非降雨水分输入的减少抵消.因此,全球变化背景下吸湿凝结水形成特征的改变将成为调节BSCs盖度和分布的重要因子. ...
A simple model for potential dewfall in an arid region
2
2002
... 在没有降雨的情况下,能够增加土壤表层水分含量的3种主要机制是雾的沉积、凝结水的形成和水蒸气的吸附.大气中的水汽含量、近地表气温和地表温度相互作用决定这3种机制的发生:①只要空气中的水汽含量达到饱和,不管地表条件如何,雾都会形成[20];②当地表温度等于或低于露点温度时,空气中的水蒸气与地表接触形成凝结水;③当地表温度高于露点温度,土壤孔隙的相对湿度低于空气相对湿度时,水蒸气的吸附作用发生,即吸湿水[21].非降雨水分有3个来源:大气、土壤和植被[22].简单的水分凝结过程分析认为,当空气气温下降,气象条件适宜(湿度与温差)时,沙丘沙开始吸取空气中的水分即吸湿水部分,这部分水分主要来源于近地层空气的水汽或沙面植株茎叶物理蒸发产生的水分.沙面继续散热使近地气层的空气水汽趋于饱和,而产生凝结过程即热凝结水,这是凝结水来源的主要部分.一般夜间沙丘上地温低于气温,热运动使水汽向下运动称为土壤水凝结.汽态水受土壤热力的规律支配,由表层向深层迁移或深层向表层迁移,属于水分的土内迁移[23].由于土壤内部水汽的凝结不会导致土壤水总量发生变化,因此有关吸湿凝结水的研究主要是指空气中的水汽在土壤和植被表面形成的吸湿凝结水. ...
... 易产生误差 | [8, 20, 41] |
| 涡度相关法 | 能量平衡 | 代表区域广泛 | 易漏掉不确定水源, ...
The improvement on the determining method of hygroscopic water of the soil
1
1996
... 在没有降雨的情况下,能够增加土壤表层水分含量的3种主要机制是雾的沉积、凝结水的形成和水蒸气的吸附.大气中的水汽含量、近地表气温和地表温度相互作用决定这3种机制的发生:①只要空气中的水汽含量达到饱和,不管地表条件如何,雾都会形成[20];②当地表温度等于或低于露点温度时,空气中的水蒸气与地表接触形成凝结水;③当地表温度高于露点温度,土壤孔隙的相对湿度低于空气相对湿度时,水蒸气的吸附作用发生,即吸湿水[21].非降雨水分有3个来源:大气、土壤和植被[22].简单的水分凝结过程分析认为,当空气气温下降,气象条件适宜(湿度与温差)时,沙丘沙开始吸取空气中的水分即吸湿水部分,这部分水分主要来源于近地层空气的水汽或沙面植株茎叶物理蒸发产生的水分.沙面继续散热使近地气层的空气水汽趋于饱和,而产生凝结过程即热凝结水,这是凝结水来源的主要部分.一般夜间沙丘上地温低于气温,热运动使水汽向下运动称为土壤水凝结.汽态水受土壤热力的规律支配,由表层向深层迁移或深层向表层迁移,属于水分的土内迁移[23].由于土壤内部水汽的凝结不会导致土壤水总量发生变化,因此有关吸湿凝结水的研究主要是指空气中的水汽在土壤和植被表面形成的吸湿凝结水. ...
土壤吸湿水测定方法的改进
1
1996
... 在没有降雨的情况下,能够增加土壤表层水分含量的3种主要机制是雾的沉积、凝结水的形成和水蒸气的吸附.大气中的水汽含量、近地表气温和地表温度相互作用决定这3种机制的发生:①只要空气中的水汽含量达到饱和,不管地表条件如何,雾都会形成[20];②当地表温度等于或低于露点温度时,空气中的水蒸气与地表接触形成凝结水;③当地表温度高于露点温度,土壤孔隙的相对湿度低于空气相对湿度时,水蒸气的吸附作用发生,即吸湿水[21].非降雨水分有3个来源:大气、土壤和植被[22].简单的水分凝结过程分析认为,当空气气温下降,气象条件适宜(湿度与温差)时,沙丘沙开始吸取空气中的水分即吸湿水部分,这部分水分主要来源于近地层空气的水汽或沙面植株茎叶物理蒸发产生的水分.沙面继续散热使近地气层的空气水汽趋于饱和,而产生凝结过程即热凝结水,这是凝结水来源的主要部分.一般夜间沙丘上地温低于气温,热运动使水汽向下运动称为土壤水凝结.汽态水受土壤热力的规律支配,由表层向深层迁移或深层向表层迁移,属于水分的土内迁移[23].由于土壤内部水汽的凝结不会导致土壤水总量发生变化,因此有关吸湿凝结水的研究主要是指空气中的水汽在土壤和植被表面形成的吸湿凝结水. ...
On the contribution to dew formation
1
1988
... 在没有降雨的情况下,能够增加土壤表层水分含量的3种主要机制是雾的沉积、凝结水的形成和水蒸气的吸附.大气中的水汽含量、近地表气温和地表温度相互作用决定这3种机制的发生:①只要空气中的水汽含量达到饱和,不管地表条件如何,雾都会形成[20];②当地表温度等于或低于露点温度时,空气中的水蒸气与地表接触形成凝结水;③当地表温度高于露点温度,土壤孔隙的相对湿度低于空气相对湿度时,水蒸气的吸附作用发生,即吸湿水[21].非降雨水分有3个来源:大气、土壤和植被[22].简单的水分凝结过程分析认为,当空气气温下降,气象条件适宜(湿度与温差)时,沙丘沙开始吸取空气中的水分即吸湿水部分,这部分水分主要来源于近地层空气的水汽或沙面植株茎叶物理蒸发产生的水分.沙面继续散热使近地气层的空气水汽趋于饱和,而产生凝结过程即热凝结水,这是凝结水来源的主要部分.一般夜间沙丘上地温低于气温,热运动使水汽向下运动称为土壤水凝结.汽态水受土壤热力的规律支配,由表层向深层迁移或深层向表层迁移,属于水分的土内迁移[23].由于土壤内部水汽的凝结不会导致土壤水总量发生变化,因此有关吸湿凝结水的研究主要是指空气中的水汽在土壤和植被表面形成的吸湿凝结水. ...
Condensed vapor and its role in the ecological environment of Shapotou region
2
1992
... 在没有降雨的情况下,能够增加土壤表层水分含量的3种主要机制是雾的沉积、凝结水的形成和水蒸气的吸附.大气中的水汽含量、近地表气温和地表温度相互作用决定这3种机制的发生:①只要空气中的水汽含量达到饱和,不管地表条件如何,雾都会形成[20];②当地表温度等于或低于露点温度时,空气中的水蒸气与地表接触形成凝结水;③当地表温度高于露点温度,土壤孔隙的相对湿度低于空气相对湿度时,水蒸气的吸附作用发生,即吸湿水[21].非降雨水分有3个来源:大气、土壤和植被[22].简单的水分凝结过程分析认为,当空气气温下降,气象条件适宜(湿度与温差)时,沙丘沙开始吸取空气中的水分即吸湿水部分,这部分水分主要来源于近地层空气的水汽或沙面植株茎叶物理蒸发产生的水分.沙面继续散热使近地气层的空气水汽趋于饱和,而产生凝结过程即热凝结水,这是凝结水来源的主要部分.一般夜间沙丘上地温低于气温,热运动使水汽向下运动称为土壤水凝结.汽态水受土壤热力的规律支配,由表层向深层迁移或深层向表层迁移,属于水分的土内迁移[23].由于土壤内部水汽的凝结不会导致土壤水总量发生变化,因此有关吸湿凝结水的研究主要是指空气中的水汽在土壤和植被表面形成的吸湿凝结水. ...
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
沙坡头地区凝结水及其在生态环境中的意义
2
1992
... 在没有降雨的情况下,能够增加土壤表层水分含量的3种主要机制是雾的沉积、凝结水的形成和水蒸气的吸附.大气中的水汽含量、近地表气温和地表温度相互作用决定这3种机制的发生:①只要空气中的水汽含量达到饱和,不管地表条件如何,雾都会形成[20];②当地表温度等于或低于露点温度时,空气中的水蒸气与地表接触形成凝结水;③当地表温度高于露点温度,土壤孔隙的相对湿度低于空气相对湿度时,水蒸气的吸附作用发生,即吸湿水[21].非降雨水分有3个来源:大气、土壤和植被[22].简单的水分凝结过程分析认为,当空气气温下降,气象条件适宜(湿度与温差)时,沙丘沙开始吸取空气中的水分即吸湿水部分,这部分水分主要来源于近地层空气的水汽或沙面植株茎叶物理蒸发产生的水分.沙面继续散热使近地气层的空气水汽趋于饱和,而产生凝结过程即热凝结水,这是凝结水来源的主要部分.一般夜间沙丘上地温低于气温,热运动使水汽向下运动称为土壤水凝结.汽态水受土壤热力的规律支配,由表层向深层迁移或深层向表层迁移,属于水分的土内迁移[23].由于土壤内部水汽的凝结不会导致土壤水总量发生变化,因此有关吸湿凝结水的研究主要是指空气中的水汽在土壤和植被表面形成的吸湿凝结水. ...
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
Dew fall in sand dune areas of India
1
1983
| [24-26] |
| 称重法 | Hiltner凝结水天平等 | 重量变化 | 连续测定 | 与天然凝结面有差别、水分不能进入土壤 | [27-29] |
自动称重式蒸渗仪、 ...
Dew variability within a small arid drainage basin in the Negev Highlands, Israel
1
2000
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
The role of dew as a moisture source for sand microbiotic crusts in the Negev Desert, Israel
4
2002
| [24-26] |
| 称重法 | Hiltner凝结水天平等 | 重量变化 | 连续测定 | 与天然凝结面有差别、水分不能进入土壤 | [27-29] |
自动称重式蒸渗仪、 ...
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Six years of dew observmion in the Negev Desert, lsrael
2
1996
| [24-26] |
| 称重法 | Hiltner凝结水天平等 | 重量变化 | 连续测定 | 与天然凝结面有差别、水分不能进入土壤 | [27-29] |
自动称重式蒸渗仪、 ...
... 影响精度 | [27-37] |
间接 ...
On the relationship between forests and occult precipitation (dew and fog precipitation)
2001
The role of dew in the water and heat balance of bare loess soil in the Negev Desert: quantifying the actual dew deposition on the soil surface
1
2002
| [24-26] |
| 称重法 | Hiltner凝结水天平等 | 重量变化 | 连续测定 | 与天然凝结面有差别、水分不能进入土壤 | [27-29] |
自动称重式蒸渗仪、 ...
Dew effects on passive microwave observations of land surfaces
2
1999
| 热传导能力变化 | 简单、易操作、连续测定 | 干扰表土层,热导度影响凝结和蒸发,降低了测量精度 | [30, 49] |
模型模拟 ...
... 微波雷达等 | 遥感信号变化 | 无破坏、范围广、时间长 | 要求遥感信号灵敏度高,受环境因子影响大,精确度不高 | [30, 51-52] |
许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
Measurement and simulation of dew accumulation and drying in a potato canopy
1
1999
... 导致精确度不高
[31-32, 34-35, 50] | | 遥感测算法 | 微波辐射计、 ...
Measuring dew formation and its threshold value for net radiation loss on top leaves in a paddy rice crop by using the dewball: a new and simple instrument
1
2000
| [31-32, 34-35, 50] |
| 遥感测算法 | 微波辐射计、 ...
Dew measurements along a longitudinal sand dune transect, Negev Desert, Israel
1
2000
| [33, 42-43] |
| 稳定同位素技术 | 氢氧同位素分析仪 | 同位素数量变化 | 无破坏、定量化程度高 | 对仪器设备要求高, ...
A simple cloud-based energy balance model to estimate dew
1
2002
| [31-32, 34-35, 50] |
| 遥感测算法 | 微波辐射计、 ...
Measurement and modelling of dew in island, coastal and alpine areas
1
2005
| [31-32, 34-35, 50] |
| 遥感测算法 | 微波辐射计、 ...
Effects of microbiotic crusts on dew deposition in the restored vegetation area at Shapotou, Northwest China
4
2006
... 不同BSCs类型显著影响吸湿凝结水的形成[7,36-37].研究表明,与裸地相比,BSCs表层吸湿凝结水形成量更高,并且与蓝藻结皮相比,由地衣和苔藓组成的BSCs能更有效地形成吸湿凝结水[36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... [36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... [36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... [36-37]. ...
Dew formation characteristics in a revegetation-stabilized desert ecosystem in Shapotou area, Northern China
5
2010
| [27-37] |
间接 ...
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 不同BSCs类型显著影响吸湿凝结水的形成[7,36-37].研究表明,与裸地相比,BSCs表层吸湿凝结水形成量更高,并且与蓝藻结皮相比,由地衣和苔藓组成的BSCs能更有效地形成吸湿凝结水[36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... [37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... -37]. ...
Dew contribution to the annual water balances in semi-arid desert valleys
1
1999
| [38-40] |
| Penman-Monteith公式 | 水量平衡 | 参数容易获得、比直接测量方便 | 受水平衡计算过程中 ...
The energy balance, evapo-transpiration and nocturnal dew deposition of an arid valley in the Andes
2006
Dew formation and its variation in haloxylon ammodendron plantations at the edge of a desert oasis, northwestern China
1
2017
| [38-40] |
| Penman-Monteith公式 | 水量平衡 | 参数容易获得、比直接测量方便 | 受水平衡计算过程中 ...
Determining dew and hoar frost formation for a low mountain range and alpine grassland site by weighable lysimeter
1
2018
| [8, 20, 41] |
| 涡度相关法 | 能量平衡 | 代表区域广泛 | 易漏掉不确定水源, ...
Estimating fog deposition at a Puerto Rican elfin cloud forest site
1
2006
| [33, 42-43] |
| 稳定同位素技术 | 氢氧同位素分析仪 | 同位素数量变化 | 无破坏、定量化程度高 | 对仪器设备要求高, ...
Dew measurement by Eddy covariance and wetness sensor in a semiarid ecosystem of SE Spain
1
2007
| [33, 42-43] |
| 稳定同位素技术 | 氢氧同位素分析仪 | 同位素数量变化 | 无破坏、定量化程度高 | 对仪器设备要求高, ...
Evaluating water fluxes of field-grown alfalfa from diurnal observations of natural isotopes concentrations, energy budget and ecophysiological parameters
1
1989
| [44-47] |
电子传导 ...
δ18O of water vapor, evapotranspiration and the site of leaf water evaporation in a soybean canopy
2008
Transition of stable isotope ratios of leaf water under simulated dew formation
2011
Dew water isotopic ratios and their relationships to ecosystem water pools and fluxes in a cropland and a grass land in China
1
2012
| [44-47] |
电子传导 ...
Role of dewfall in the water balance of a semiarid coastal steppe ecosystem
2
2014
| 推算误差较大 | [48] |
电子传导土壤—水汽 ...
... 吸湿凝结水生态作用评价方面的主要限制是精确测量水分输入较为困难.不同的测量方法都有其优缺点和适用条件,可根据实际情况选择最优测量方法.随着现代技术的发展,吸湿凝结水直接和间接测定方法的联合使用将进一步提高测量的精度.Uclés等[48]使用了一种组合凝结水估计方法(the Combined Dewfall Estimation Methods, CDEM),联合了Penman-Monteith 蒸发模型、叶面湿度传感器和补充气象信息,通过叶面湿度传感器的数据,CDEM 消除了测量和界定凝结水事件的主观性.同位素技术的应用为吸湿凝结水测量提供了新的契机,吸湿凝结水的同位素组成及与其他生态系统水源的关系受到广泛关注,大大提高了吸湿凝结水测量的精度. ...
An electrical conductance method for determining condensation and evaporation processes in arid soils with high spatial resolution
1
1980
| 热传导能力变化 | 简单、易操作、连续测定 | 干扰表土层,热导度影响凝结和蒸发,降低了测量精度 | [30, 49] |
模型模拟 ...
Method exploring on dew condensation monitoring in wetland ecosystem
1
2010
| [31-32, 34-35, 50] |
| 遥感测算法 | 微波辐射计、 ...
Monitoring water interception by crop fields from passive microwave observations
1
1996
| 遥感信号变化 | 无破坏、范围广、时间长 | 要求遥感信号灵敏度高,受环境因子影响大,精确度不高 | [30, 51-52] |
许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
Radar backscatter characteristics of a desert surface
1
1996
... 微波雷达等
遥感信号变化 | 无破坏、范围广、时间长 | 要求遥感信号灵敏度高,受环境因子影响大,精确度不高 | [30, 51-52] | 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
Dynamic measurement and theoretical calculation on water absorption and condensation of sandy soil in Shapotou region
1
1998
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
沙坡头地区土壤水分吸湿凝结的动态观测与理论计算
1
1998
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
An experimental study on the condensation water in the plain area of the northern slope of Tianshan Mountains
1
2002
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
天山北麓平原区凝结水的观测试验分析
1
2002
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
An overview on soil condensate in arid and semiarid regions in China
1
2005
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
中国干旱半干旱地区土壤凝结水研究综述
1
2005
... 许多学者在观测凝结水时,发现空气相对湿度还远未达到饱和,但微型测渗计的重量就开始增加,这时测量到的是土壤吸湿水而不是真正意义上的土壤凝结水;也就是说,在土壤特别干燥的情况下,微型测渗计重量的增加过程分2个阶段:第一阶段是从午后沙土含水量降至最低点开始至凌晨凝结水形成前,历时长,变化慢,水量增加不大(最大吸水量不超过沙土吸湿水量),这是沙土的吸湿凝结;第二阶段出现在凌晨日出前,历时短、变化快、呈跃变态,这是由气象因素决定的水分凝结过程[23,53].如某些试验空气相对湿度在20%~40%就观测到土壤吸湿水[54].我国学者在不同地区进行的凝结水试验,大多数测量结果是吸湿水和凝结水的混合量,确切地应该称作吸湿凝结量.为了准确测定凝结量,试验中应该密切配合温度和相对湿度的观测,只有温度低于等于露点或相对湿度达到饱和时,土壤所增加的重量才是真正的凝结量;或者,在测定凝结水之前,事先测定好土壤的最大吸湿量,把测到的超过最大吸湿量的部分作为凝结量[55]. ...
Microclimatic boundary conditions for activity of soil lichen crusts in sand dunes of the north-western Negev Desert, Israel
3
2001
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... 总之,BSCs主要分布于土壤表层,对小降水事件反应敏感,相对应的,吸湿凝结水生成数量小,集中在土壤表层,几乎每天都发生,因此,相较降雨,干旱区频繁发生的吸湿凝结水事件对BSCs的生存尤为重要[88].BSCs在干旱条件下生存的关键是能够在有水的条件下,短时间内有效激活代谢和生长,并在脱水期间延缓代谢活动[90].因此,吸湿凝结水导致的昼夜干湿交替过程中,BSCs中的生物激活光合和呼吸作用活动的机制相当重要[65],但目前知之甚少[56].Potts[89]和Billi等[91]就地木耳(Nostoc commune)对水合/脱水循环的干旱忍受能力的适应性研究得出了一些重要成果,认为脱水过程中现有蛋白质的稳定性确保了地木耳在有利条件下快速重新生长;在水合和脱水期间,光能通量转移和 CO2 固定的协调也是特别重要的;另外,胞外多糖(Exopoly Saccharides,EPS)的分泌被认为是荒漠藻类结皮中蓝藻存活的主要保护途径之一,其也可以通过调节碳水化合物代谢来提高其脱水耐性而减少氧化损伤.吸湿凝结水还有一个特点,夜间生成时间较长,可能对呼吸作用的影响更大,且不同种类BSCs的光合和呼吸作用对水分的敏感程度不同,因此其对BSCs固碳过程的影响比我们预想的更为复杂.目前我们还没有完全理解BSCs净光合作用和土壤碳吸收的必要条件,为了更好地分离影响旱地BSCs碳流相互依存的生物和物理变量,还需要大量的野外和室内试验相结合. ...
Dewfall and its geo-ecological implication for biological surface crusts in desert sand dunes (northwestern Negev, Israel)
1
2006
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Effects of sand burial on dew deposition on moss soil crust in a revegetated area of the tennger desert, northern china
2
2014
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 不同BSCs类型显著影响吸湿凝结水的形成[7,36-37].研究表明,与裸地相比,BSCs表层吸湿凝结水形成量更高,并且与蓝藻结皮相比,由地衣和苔藓组成的BSCs能更有效地形成吸湿凝结水[36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
Taxonomic composition and photosynthetic characteristics of the biological crusts covering sand dunes in the western Negev
2
1992
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
Eight days in the life of a desert lichen: water relations and photosynthesis of Teloschistes capensis in the coastal fog zone in the Namib Desert
1
1990
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Cyanobacteria are confined to dewless habitats within a dew desert: implications for past and future climate change for lithic microorganisms
1
2014
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
An investigation of the role of water(rain and dew) in controlling the growth form of lichens on cobbles in the Negev Desert
1
2011
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Lichen colonization on cobbles in the Negev Desert following 15 years in the field
1
2010
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
The CO2 exchange of biological soil crusts in a semiarid grass-shrubland at the northern transition zone of the Negev Desert, Israel
2
2008
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Influence of dew on biomass and photosystem II activity of cyanobacterial crusts in the Hopq Desert, northwest China
4
2009
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... 总之,BSCs主要分布于土壤表层,对小降水事件反应敏感,相对应的,吸湿凝结水生成数量小,集中在土壤表层,几乎每天都发生,因此,相较降雨,干旱区频繁发生的吸湿凝结水事件对BSCs的生存尤为重要[88].BSCs在干旱条件下生存的关键是能够在有水的条件下,短时间内有效激活代谢和生长,并在脱水期间延缓代谢活动[90].因此,吸湿凝结水导致的昼夜干湿交替过程中,BSCs中的生物激活光合和呼吸作用活动的机制相当重要[65],但目前知之甚少[56].Potts[89]和Billi等[91]就地木耳(Nostoc commune)对水合/脱水循环的干旱忍受能力的适应性研究得出了一些重要成果,认为脱水过程中现有蛋白质的稳定性确保了地木耳在有利条件下快速重新生长;在水合和脱水期间,光能通量转移和 CO2 固定的协调也是特别重要的;另外,胞外多糖(Exopoly Saccharides,EPS)的分泌被认为是荒漠藻类结皮中蓝藻存活的主要保护途径之一,其也可以通过调节碳水化合物代谢来提高其脱水耐性而减少氧化损伤.吸湿凝结水还有一个特点,夜间生成时间较长,可能对呼吸作用的影响更大,且不同种类BSCs的光合和呼吸作用对水分的敏感程度不同,因此其对BSCs固碳过程的影响比我们预想的更为复杂.目前我们还没有完全理解BSCs净光合作用和土壤碳吸收的必要条件,为了更好地分离影响旱地BSCs碳流相互依存的生物和物理变量,还需要大量的野外和室内试验相结合. ...
... 吸湿凝结水的生成量和持续时间在很大程度上取决于生态系统中的天气条件、微生境和凝结面特征[92].BSCs的发育改变了地表微生境和凝结面特征,通过地表微地形的变化,BSCs增加了其吸收的表面积[65],降低了表层风速;与流沙相比,BSCs的热容量较低,日间变暖和夜间降温速度都较快,增加了昼夜温差;较高的细物质和有机物含量使得BSCs容重降低,持水力增加,黏粉粒含量增加;BSCs具有较高的孔隙率和胞外多糖,因此具有较高的水容量,吸湿凝结水的形成使得地表大气达到饱和的时间延迟,导致大气水汽向着地表转移,也就是向水容量较高的BSCs地表土壤孔隙和胞外多糖中转移[8];这些都有利于吸湿凝结水在其表面的形成.另外,BSCs中生物的形成有利于吸收水分,藓类结皮能通过假根和丝状体等吸收吸湿凝结水[93];虽然齿肋赤藓Syntrichia caninervis Mitt的假根系统(rhizoids)不具有吸收水分的作用,其叶片顶端的白色芒尖有利于凝结水的形成,叶片顶端的芒尖分布有从纳米、微米到厘米等不同尺度的水分收集与传输系统(凹槽和疣状突起),具有采集和运输水分的多种模式,分别将空气中的水分子形成水核、水膜和水滴,然后将这些水分通过毛细管作用迅速运输到芒尖底部的叶面,被叶片吸收[94]. ...
Lichens as biomarkers for dew precipitation in the Negev Desert
1
2011
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
The effect of dew on flint and limestone lichen communities in the Negev Desert
1
2015
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Early morning photosynthesis of the moss Tortula ruralis following summer dew fall in a Hungarian temperate dry sandy grassland
3
2000
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... [68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
Ecological effect of hygroscopic and condensate water on biological soil crusts in Shapotou region of China
1
2013
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
沙坡头地区吸湿凝结水对生物土壤结皮的生态作用
1
2013
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Photosynthetic responses of a moss, Tortula ruralis ssp. ruralis, and the lichens Cladonia convoluta and C. furcata to water deficit and short periods of desiccation, and their ecophysiological significance: a baseline study at present-day CO2 concentration
2
1996
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
Angle and aspect dependent dew and fog precipitation in the Negev Desert
1
2005
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Do vehicle track disturbances affect the productivity of soil-growing lichens in a fog desert?
1
2006
... 隐花植物可能是新的陆地表层的第一批殖民者,从极地到沙漠的许多生态系统中土壤表层都有BSCs覆盖[8],尤其是旱地生态系统中,比如智利的阿塔卡马沙漠、纳米比亚的纳米布沙漠、南非的多汁卡鲁沙漠、以色列的内盖夫沙漠以及中国的腾格里沙漠和古尔班通古特沙漠等.在这些生态系统中,干旱是影响BSCs发育的最显著的生态因素,而吸湿凝结水被认为是其主要的水分来源之一[9,25,56-57].吸湿凝结水产生的水脉冲在大小和频率上与降雨产生的水脉冲不同,其生成量小,不能补充BSCs下层土壤,但频率高,能够引起土壤表层频繁地水合/脱水循环,影响表层BSCs的存活[8,37].BSCs中的生物体是随水植物,生物量低,其功能对水分的变化很敏感[58].由蓝绿藻组成的BSCs在水分含量大于0.1 mm的条件下即具有生物活性[59];在热带和沿海沙漠中,地衣物种广泛分布,有时甚至构成该区主要的植被类型,这很大程度上是因为地衣利用吸湿凝结水作为水源的能力[60];因此,吸湿凝结水是控制BSCs中蓝藻和地衣分布的一个重要因子[61],对其生长[9,62]和快速繁殖[63]起着重要作用.地衣结皮对小的水分脉冲具有迅速的生理响应[64-65],通过对内盖夫沙漠中部鹅卵石上地衣群落结构的比较,地衣结皮叶绿素含量的变化具有明显的海拔梯度[9],特定地衣物种可以作为吸湿凝结水生成量和持续时间的生物指标[66-67].苔藓结皮也能够吸附非降雨水分用于自身的光合作用[68],潘颜霞等[69]的研究表明,吸湿凝结水对苔藓结皮含水量无影响,但其能弥补日间表层水分的迅速蒸发,其形成量与苔藓结皮中的叶绿素含量呈正相关关系,能够提高苔藓结皮生长活性,有利于其生物量的积累.吸湿凝结水数量较大时,其能够直接湿润藓类的拟茎体(caulidium)[26],有利于清晨光合生物量的积累[68,70]和生殖器官的成熟[26],当吸湿凝结水量较少时,仍能降低蒸腾作用,使藓类结皮处于预激活状态,以响应下一次降雨事件,从而提高降雨的利用效率[64].吸湿凝结水数量和可用性减少的长期后果将间接影响碳平衡,并改变荒漠生态系统中苔藓结皮的分布和演替[26].另外,吸湿凝结水对苔藓结皮的作用极易受到环境因素变化[71]和诸如践踏等干扰的影响[72]. ...
Ecophysiological investigations on lichens of the Negev Desert. VI. Annual course of the photosynthetic production of Ramalina maciformis (Del.) Bory
1
1979
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
2
2006
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
... [74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate
1
1992
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
On carbon sequestration in desert ecosystems
1
2009
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
Large annual net ecosystem CO2 uptake of a Mojave Desert ecosystem
1
2008
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
CO2 absorption by alkaline soils and its implication to the global carbon cycle
1
2008
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
A proposed mechanism for the pulse in carbon dioxide production commonly observed following the rapid rewetting of a dry soil
1
2003
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
Response of soil CO2 efflux to water manipulation in a tallgrass prairie ecosystem
1
2002
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
Carbon dioxide fluxes from cyanobacteria crusted soils in the Kalahari
2008
How soil moisture, rain pulses and growth alter the response of ecosystem respiration to temperature
1
2004
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
Carbon dioxide fluxes from biologically-crusted Kalahari Sands after simulated wetting
1
2010
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
Simulated climate change reduced the capacity of lichen-dominated biocrusts to act as carbon sinks in two semi-arid Mediterranean ecosystems
2
2014
... 荒漠草地在全球碳循环中占有重要地位,BSCs的光合和呼吸活动是荒漠碳循环的重要组成成分,而吸湿凝结水对其水、碳循环之间的联系有强烈的调节作用.吸湿凝结水的水分脉冲可以激活BSCs内的微生物群落,促进土壤微生物矿化和有机质的分解.有研究发现0.03 mm为吸湿凝结水对BSCs中微生物有效性的阈值[73].微生物分解有机碳产生的土壤呼吸是大气CO2的最重要贡献者之一,每年释放的碳量比目前全球人为排放的碳量多1个数量级[74-75].土壤呼吸的任何增加都可能耗尽土壤碳,对肥力产生不利影响,并提高大气中的CO2浓度.尽管具有广泛的环境意义,我们对土壤呼吸控制的认知仍然不全[74],不同过程的影响率和CO2通量的数量一直是争论的焦点[76-78].提高旱地土壤CO2排放预测,在很大程度上取决于更好地了解土壤表层和地下微生物种群对水分和温度变化的响应.表层BSCs复湿后,土壤中的CO2呼吸速率通常比保持持续湿润的土壤高500%[79].因此,吸湿凝结水脉冲可能构成土壤CO2年排放总量的很大一部分[80-82].研究表明,在一个昼夜循环中,来自干燥土壤(当水分含量低于仪器的测定精度时)的CO2通量范围为2.8~14.8 mg C/(m2·h)[83],这表明BSCs中的蓝藻可以利用含量极低的吸湿凝结水激活其呼吸作用,非降雨水分的变化可能会比降雨量的变化对BSCs的碳平衡产生更大的影响[84]. ...
... BSCs位于土壤表层,对环境的敏感性高,组成简单,易于控制,是研究旱地气候变化和生态恢复的模式生态系统,其对气候变化的响应也受到了广泛关注[19].水分是影响BSCs生长发育最重要的环境因子,没有水,其他因子都不会起作用.非降雨水作为可以被BSCs利用的一种持续水资源,在未来全球气候变化背景下,其对BSCs的水碳平衡将产生重要影响,但目前相关研究还比较薄弱.Mónica等[84]在西班牙中部和东南部的研究证明了非降雨水在BSCs应对气候变化中的重要性,并强调变暖引起的非降雨水分输入改变对BSCS碳平衡的影响比降雨更大.Baldauf 等[97]成功地将一个基于过程的模型参数化,量化了大气CO2浓度、气温、降雨量和相对湿度变化对西班牙地中海沿岸地衣结皮Diploschistes diacapsis (Ach.) Lumbsch光合活性和盖度的影响,结果表明,单纯变暖会增强光合作用从而增加结皮盖度,变暖引起的结皮盖度降低主要来源于同时发生的相对湿度的降低和非降雨水分输入的减少.该研究认为在未来全球气候变化背景下,影响地衣结皮Diploschistes diacapsis (Ach.) Lumbsch存活的关键因子是相对湿度的降低,而不是气温的增加,大气CO2浓度升高的作用也可以被非降雨水分输入的减少抵消.因此,全球变化背景下吸湿凝结水形成特征的改变将成为调节BSCs盖度和分布的重要因子. ...
Patterns of CO2 exchange in biological soil crusts of successional age
1
2000
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
Photosynthesis and water relations of lichen soil crusts: field measurements in the coastal fog zone of the Namib Desert
3
1994
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... [86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... ,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
Non-rainfall water inputs: a key water source for biocrust carbon fixation
1
2021
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
Recovery of photosynthetic systems during rewetting is quite rapid in a terrestrial cyanobacterium, Nostoc commune
2
2002
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... 总之,BSCs主要分布于土壤表层,对小降水事件反应敏感,相对应的,吸湿凝结水生成数量小,集中在土壤表层,几乎每天都发生,因此,相较降雨,干旱区频繁发生的吸湿凝结水事件对BSCs的生存尤为重要[88].BSCs在干旱条件下生存的关键是能够在有水的条件下,短时间内有效激活代谢和生长,并在脱水期间延缓代谢活动[90].因此,吸湿凝结水导致的昼夜干湿交替过程中,BSCs中的生物激活光合和呼吸作用活动的机制相当重要[65],但目前知之甚少[56].Potts[89]和Billi等[91]就地木耳(Nostoc commune)对水合/脱水循环的干旱忍受能力的适应性研究得出了一些重要成果,认为脱水过程中现有蛋白质的稳定性确保了地木耳在有利条件下快速重新生长;在水合和脱水期间,光能通量转移和 CO2 固定的协调也是特别重要的;另外,胞外多糖(Exopoly Saccharides,EPS)的分泌被认为是荒漠藻类结皮中蓝藻存活的主要保护途径之一,其也可以通过调节碳水化合物代谢来提高其脱水耐性而减少氧化损伤.吸湿凝结水还有一个特点,夜间生成时间较长,可能对呼吸作用的影响更大,且不同种类BSCs的光合和呼吸作用对水分的敏感程度不同,因此其对BSCs固碳过程的影响比我们预想的更为复杂.目前我们还没有完全理解BSCs净光合作用和土壤碳吸收的必要条件,为了更好地分离影响旱地BSCs碳流相互依存的生物和物理变量,还需要大量的野外和室内试验相结合. ...
Desiccation tolerance of prokaryotes
2
1994
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... 总之,BSCs主要分布于土壤表层,对小降水事件反应敏感,相对应的,吸湿凝结水生成数量小,集中在土壤表层,几乎每天都发生,因此,相较降雨,干旱区频繁发生的吸湿凝结水事件对BSCs的生存尤为重要[88].BSCs在干旱条件下生存的关键是能够在有水的条件下,短时间内有效激活代谢和生长,并在脱水期间延缓代谢活动[90].因此,吸湿凝结水导致的昼夜干湿交替过程中,BSCs中的生物激活光合和呼吸作用活动的机制相当重要[65],但目前知之甚少[56].Potts[89]和Billi等[91]就地木耳(Nostoc commune)对水合/脱水循环的干旱忍受能力的适应性研究得出了一些重要成果,认为脱水过程中现有蛋白质的稳定性确保了地木耳在有利条件下快速重新生长;在水合和脱水期间,光能通量转移和 CO2 固定的协调也是特别重要的;另外,胞外多糖(Exopoly Saccharides,EPS)的分泌被认为是荒漠藻类结皮中蓝藻存活的主要保护途径之一,其也可以通过调节碳水化合物代谢来提高其脱水耐性而减少氧化损伤.吸湿凝结水还有一个特点,夜间生成时间较长,可能对呼吸作用的影响更大,且不同种类BSCs的光合和呼吸作用对水分的敏感程度不同,因此其对BSCs固碳过程的影响比我们预想的更为复杂.目前我们还没有完全理解BSCs净光合作用和土壤碳吸收的必要条件,为了更好地分离影响旱地BSCs碳流相互依存的生物和物理变量,还需要大量的野外和室内试验相结合. ...
Activation of photosynthesis and resistance to photoinhibition in cyanobacteria within biological desert crust
2
2004
... 尽管很多研究已经证明了BSCs是土壤表层碳循环的重要组成部分[59,85],但在这些试验期间,土壤都没有表现出显著的持续吸收碳的过程,这与Wohlfahrt等[77]的观点有差异,他们认为BSCs至少在一定程度上是荒漠生态系统碳净收益的原因.这可能是由于小的降雨事件后,BSCs中微生物的自养呼吸作用掩盖了光合作用的碳吸收.相关研究结果表明[68,70,86],蓝藻结皮需要大于2.5 h的光合作用来补偿夜间呼吸过程中的碳损失.对蓝藻和地衣结皮的研究表明,雾水提高了呼吸,使得地衣结皮的净光合释放量为0.68 μmol/(m2·s),而凝结水增加了光合,有利于结皮的碳固定,其净光合吸收率为0.66 μmol/(m2·s)[87].纳米布沙漠3种地衣结皮的研究表明,夜间吸湿凝结水的水合作用激活了暗呼吸,日出后进行短暂的正净光合作用[引起正净光合作用的最低水分含量非常小(为0.13~0.26 mm的降水量)],之后吸湿凝结水蒸发引起的干燥导致代谢失活[86].2种蓝藻具鞘微鞘藻(Microcoleus vaginatus Gom)和爪哇伪枝藻[Scytonema javanicum (Kütz.) Born et Flah]接种形成的蓝藻结皮对吸湿凝结水的响应结果表明[65],再水合50 min内,大约80%的PSII(photo system II)活性可以在黑暗、5 °C 低温下被激活,只有约20%的PSII活性具有光照和温度依赖性.但其他种类的BSCs是否能在黑暗低温条件下迅速进行光合作用有待研究,这可能也是蓝藻结皮能够在干旱环境中广泛分布的原因之一.日出之后的吸湿凝结水生成过程对BSCs生理过程的影响更大,光照条件下频繁的水分补充过程表明,BSCs的Fo值迅速增加并在大约3 min后达到最大值,然后在大约50 min后逐渐下降至恒定值.值得注意的是,之前报道的复水后光合作用的激活率存在很大差异[56,86,88].再水合之前的干旱时间也是一个重要的影响因子,干旱时间越短,光合能力的激活越快[89];如果经历长时间的干旱,比如几个月,那么再水合会很快激发呼吸作用,然后才是光合.对发菜Nostoc的研究结果表明,复水0.5 h后,呼吸作用首先被激活,6 h后测量到光合作用,50 h后才开始氮的固定.对微鞘藻占90%以上的蓝藻结皮实施相同的试验测定,如果复水前的干旱时间较长,比如6个月,那么复水后的1 h之内都测量不到光合作用,但如果模拟吸湿凝结水的频繁生成,那么复水后几分钟之内PSII活性就被激活[90].另外,生长条件的差异、物种的不同等都可能影响复水后光合作用的激活率. ...
... 总之,BSCs主要分布于土壤表层,对小降水事件反应敏感,相对应的,吸湿凝结水生成数量小,集中在土壤表层,几乎每天都发生,因此,相较降雨,干旱区频繁发生的吸湿凝结水事件对BSCs的生存尤为重要[88].BSCs在干旱条件下生存的关键是能够在有水的条件下,短时间内有效激活代谢和生长,并在脱水期间延缓代谢活动[90].因此,吸湿凝结水导致的昼夜干湿交替过程中,BSCs中的生物激活光合和呼吸作用活动的机制相当重要[65],但目前知之甚少[56].Potts[89]和Billi等[91]就地木耳(Nostoc commune)对水合/脱水循环的干旱忍受能力的适应性研究得出了一些重要成果,认为脱水过程中现有蛋白质的稳定性确保了地木耳在有利条件下快速重新生长;在水合和脱水期间,光能通量转移和 CO2 固定的协调也是特别重要的;另外,胞外多糖(Exopoly Saccharides,EPS)的分泌被认为是荒漠藻类结皮中蓝藻存活的主要保护途径之一,其也可以通过调节碳水化合物代谢来提高其脱水耐性而减少氧化损伤.吸湿凝结水还有一个特点,夜间生成时间较长,可能对呼吸作用的影响更大,且不同种类BSCs的光合和呼吸作用对水分的敏感程度不同,因此其对BSCs固碳过程的影响比我们预想的更为复杂.目前我们还没有完全理解BSCs净光合作用和土壤碳吸收的必要条件,为了更好地分离影响旱地BSCs碳流相互依存的生物和物理变量,还需要大量的野外和室内试验相结合. ...
Life and death of dried prokaryotes
1
2002
... 总之,BSCs主要分布于土壤表层,对小降水事件反应敏感,相对应的,吸湿凝结水生成数量小,集中在土壤表层,几乎每天都发生,因此,相较降雨,干旱区频繁发生的吸湿凝结水事件对BSCs的生存尤为重要[88].BSCs在干旱条件下生存的关键是能够在有水的条件下,短时间内有效激活代谢和生长,并在脱水期间延缓代谢活动[90].因此,吸湿凝结水导致的昼夜干湿交替过程中,BSCs中的生物激活光合和呼吸作用活动的机制相当重要[65],但目前知之甚少[56].Potts[89]和Billi等[91]就地木耳(Nostoc commune)对水合/脱水循环的干旱忍受能力的适应性研究得出了一些重要成果,认为脱水过程中现有蛋白质的稳定性确保了地木耳在有利条件下快速重新生长;在水合和脱水期间,光能通量转移和 CO2 固定的协调也是特别重要的;另外,胞外多糖(Exopoly Saccharides,EPS)的分泌被认为是荒漠藻类结皮中蓝藻存活的主要保护途径之一,其也可以通过调节碳水化合物代谢来提高其脱水耐性而减少氧化损伤.吸湿凝结水还有一个特点,夜间生成时间较长,可能对呼吸作用的影响更大,且不同种类BSCs的光合和呼吸作用对水分的敏感程度不同,因此其对BSCs固碳过程的影响比我们预想的更为复杂.目前我们还没有完全理解BSCs净光合作用和土壤碳吸收的必要条件,为了更好地分离影响旱地BSCs碳流相互依存的生物和物理变量,还需要大量的野外和室内试验相结合. ...
The importance of dew in the water balance of a continental semiarid grassland
1
2019
... 吸湿凝结水的生成量和持续时间在很大程度上取决于生态系统中的天气条件、微生境和凝结面特征[92].BSCs的发育改变了地表微生境和凝结面特征,通过地表微地形的变化,BSCs增加了其吸收的表面积[65],降低了表层风速;与流沙相比,BSCs的热容量较低,日间变暖和夜间降温速度都较快,增加了昼夜温差;较高的细物质和有机物含量使得BSCs容重降低,持水力增加,黏粉粒含量增加;BSCs具有较高的孔隙率和胞外多糖,因此具有较高的水容量,吸湿凝结水的形成使得地表大气达到饱和的时间延迟,导致大气水汽向着地表转移,也就是向水容量较高的BSCs地表土壤孔隙和胞外多糖中转移[8];这些都有利于吸湿凝结水在其表面的形成.另外,BSCs中生物的形成有利于吸收水分,藓类结皮能通过假根和丝状体等吸收吸湿凝结水[93];虽然齿肋赤藓Syntrichia caninervis Mitt的假根系统(rhizoids)不具有吸收水分的作用,其叶片顶端的白色芒尖有利于凝结水的形成,叶片顶端的芒尖分布有从纳米、微米到厘米等不同尺度的水分收集与传输系统(凹槽和疣状突起),具有采集和运输水分的多种模式,分别将空气中的水分子形成水核、水膜和水滴,然后将这些水分通过毛细管作用迅速运输到芒尖底部的叶面,被叶片吸收[94]. ...
Cementing mechanism of algal crusts from desert area
1
2002
... 吸湿凝结水的生成量和持续时间在很大程度上取决于生态系统中的天气条件、微生境和凝结面特征[92].BSCs的发育改变了地表微生境和凝结面特征,通过地表微地形的变化,BSCs增加了其吸收的表面积[65],降低了表层风速;与流沙相比,BSCs的热容量较低,日间变暖和夜间降温速度都较快,增加了昼夜温差;较高的细物质和有机物含量使得BSCs容重降低,持水力增加,黏粉粒含量增加;BSCs具有较高的孔隙率和胞外多糖,因此具有较高的水容量,吸湿凝结水的形成使得地表大气达到饱和的时间延迟,导致大气水汽向着地表转移,也就是向水容量较高的BSCs地表土壤孔隙和胞外多糖中转移[8];这些都有利于吸湿凝结水在其表面的形成.另外,BSCs中生物的形成有利于吸收水分,藓类结皮能通过假根和丝状体等吸收吸湿凝结水[93];虽然齿肋赤藓Syntrichia caninervis Mitt的假根系统(rhizoids)不具有吸收水分的作用,其叶片顶端的白色芒尖有利于凝结水的形成,叶片顶端的芒尖分布有从纳米、微米到厘米等不同尺度的水分收集与传输系统(凹槽和疣状突起),具有采集和运输水分的多种模式,分别将空气中的水分子形成水核、水膜和水滴,然后将这些水分通过毛细管作用迅速运输到芒尖底部的叶面,被叶片吸收[94]. ...
The upside-down water collection system of Syntrichia caninervis
1
2016
... 吸湿凝结水的生成量和持续时间在很大程度上取决于生态系统中的天气条件、微生境和凝结面特征[92].BSCs的发育改变了地表微生境和凝结面特征,通过地表微地形的变化,BSCs增加了其吸收的表面积[65],降低了表层风速;与流沙相比,BSCs的热容量较低,日间变暖和夜间降温速度都较快,增加了昼夜温差;较高的细物质和有机物含量使得BSCs容重降低,持水力增加,黏粉粒含量增加;BSCs具有较高的孔隙率和胞外多糖,因此具有较高的水容量,吸湿凝结水的形成使得地表大气达到饱和的时间延迟,导致大气水汽向着地表转移,也就是向水容量较高的BSCs地表土壤孔隙和胞外多糖中转移[8];这些都有利于吸湿凝结水在其表面的形成.另外,BSCs中生物的形成有利于吸收水分,藓类结皮能通过假根和丝状体等吸收吸湿凝结水[93];虽然齿肋赤藓Syntrichia caninervis Mitt的假根系统(rhizoids)不具有吸收水分的作用,其叶片顶端的白色芒尖有利于凝结水的形成,叶片顶端的芒尖分布有从纳米、微米到厘米等不同尺度的水分收集与传输系统(凹槽和疣状突起),具有采集和运输水分的多种模式,分别将空气中的水分子形成水核、水膜和水滴,然后将这些水分通过毛细管作用迅速运输到芒尖底部的叶面,被叶片吸收[94]. ...
Mechanism of biocrusts boosting and utilizing non-rainfall water in Hobq Desert of China
1
2017
... 不同BSCs类型显著影响吸湿凝结水的形成[7,36-37].研究表明,与裸地相比,BSCs表层吸湿凝结水形成量更高,并且与蓝藻结皮相比,由地衣和苔藓组成的BSCs能更有效地形成吸湿凝结水[36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
Biocrusts enhance non-rainfall water deposition and alter its distribution in dryland soils
2
2021
... 不同BSCs类型显著影响吸湿凝结水的形成[7,36-37].研究表明,与裸地相比,BSCs表层吸湿凝结水形成量更高,并且与蓝藻结皮相比,由地衣和苔藓组成的BSCs能更有效地形成吸湿凝结水[36,95-96].在中国古尔班通古特沙漠地区的研究表明,苔藓结皮表层形成的吸湿凝结水量最大,地衣和蓝藻结皮表层吸湿凝结水形成量相似且相较苔藓结皮显著减少,流沙表层的吸湿凝结水形成量最少;而流沙表层吸湿凝结水的生成间期最长,其次是蓝藻结皮和苔藓结皮,生成间期最短的是地衣结皮[7].在中国腾格里沙漠的研究表明,流沙和物理结皮表层吸湿凝结水形成量没有显著差异,但两者与BSCs表层的吸湿凝结水形成量差异显著(p<0.05),不同凝结面表层吸湿凝结水形成间期的研究结果与以上相反,苔藓结皮表层吸湿凝结水的形成间期最长[37].腾格里沙漠3种藓类结皮对沙埋造成的吸湿凝结水生成量降低的抵抗力比较结果表明,真藓结皮Bryum argenteum表现出最高的耐受性,其次是以土生对齿藓Didymodon vinealis和齿肋赤藓Syntrichia caninervis为主的BSCs [58].中国黄土高原地区的研究表明,相较裸地,苔藓结皮表层吸湿凝结水形成量增加了约70%[96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
... [96].因此,隐花植物的出现可以作为凝结水形成数量迅速增加的指示剂[36].一方面,BSCs的发育有利于吸湿凝结水的生成,另一方面,吸湿凝结水的生成促进了BSCs的发育.BSCs的出现和发展指示了流动沙漠向固定和半固定沙漠转化的重要阶段,可被用作生态环境健康评价的指标,其时空变化可揭示干旱沙漠地区生态环境的演变趋势和对全球变化的响应规律,吸湿凝结水通过影响BSCs从而成为荒漠生态系统演化的重要驱动力[36-37]. ...
Relative humidity predominantly determines long-term biocrust-forming lichen cover in drylands under climate change
1
2021
... BSCs位于土壤表层,对环境的敏感性高,组成简单,易于控制,是研究旱地气候变化和生态恢复的模式生态系统,其对气候变化的响应也受到了广泛关注[19].水分是影响BSCs生长发育最重要的环境因子,没有水,其他因子都不会起作用.非降雨水作为可以被BSCs利用的一种持续水资源,在未来全球气候变化背景下,其对BSCs的水碳平衡将产生重要影响,但目前相关研究还比较薄弱.Mónica等[84]在西班牙中部和东南部的研究证明了非降雨水在BSCs应对气候变化中的重要性,并强调变暖引起的非降雨水分输入改变对BSCS碳平衡的影响比降雨更大.Baldauf 等[97]成功地将一个基于过程的模型参数化,量化了大气CO2浓度、气温、降雨量和相对湿度变化对西班牙地中海沿岸地衣结皮Diploschistes diacapsis (Ach.) Lumbsch光合活性和盖度的影响,结果表明,单纯变暖会增强光合作用从而增加结皮盖度,变暖引起的结皮盖度降低主要来源于同时发生的相对湿度的降低和非降雨水分输入的减少.该研究认为在未来全球气候变化背景下,影响地衣结皮Diploschistes diacapsis (Ach.) Lumbsch存活的关键因子是相对湿度的降低,而不是气温的增加,大气CO2浓度升高的作用也可以被非降雨水分输入的减少抵消.因此,全球变化背景下吸湿凝结水形成特征的改变将成为调节BSCs盖度和分布的重要因子. ...
Projected climate change impacts upon dew yield in the Mediterranean Basin
1
2016
... 在最近的全球变暖期间,中国西北干旱地区的地表变暖大于湿润和半湿润地区,这导致地表相对湿度的下降幅度更大,进一步会导致吸湿凝结水的生成频率降低.中国597个连续观测站点的研究(1961—2010年)表明,由于近地层变暖和相对湿度降低,凝结水的生成频率每10年减少了5.2天,与20世纪60年代相比,21世纪初的凝结水生成频率在湿润、半湿润(降雨量为400~800 mm/a)和干旱区分别下降了28%、40%和50%.在地中海地区的研究也表明,未来数十年内,随着全球变暖,旱地的气温升高,蒸发增加,吸湿凝结水的数量和间期都可能减少[98].由于吸湿凝结水极大地促进了干旱地区的地表水平衡,因此吸湿凝结水频率的大幅下降可能使得干旱区对全球变暖做出“更干燥的反应”[99].最近的研究指出,吸湿凝结水对未来气候变暖有显著响应,并强调在气候变暖条件下,微生境条件和植物特性共同影响吸湿凝结水的形成,对未来的生态系统过程具有重要的潜在作用[100].中国腾格里沙漠南缘的全球变暖长期模拟试验结果表明,长期增温和减雨使得藓类结皮的盖度显著降低,从而导致凝结水数量的减少[101]. ...
Near-surface warming reduces dew frequency in China
1
2021
... 在最近的全球变暖期间,中国西北干旱地区的地表变暖大于湿润和半湿润地区,这导致地表相对湿度的下降幅度更大,进一步会导致吸湿凝结水的生成频率降低.中国597个连续观测站点的研究(1961—2010年)表明,由于近地层变暖和相对湿度降低,凝结水的生成频率每10年减少了5.2天,与20世纪60年代相比,21世纪初的凝结水生成频率在湿润、半湿润(降雨量为400~800 mm/a)和干旱区分别下降了28%、40%和50%.在地中海地区的研究也表明,未来数十年内,随着全球变暖,旱地的气温升高,蒸发增加,吸湿凝结水的数量和间期都可能减少[98].由于吸湿凝结水极大地促进了干旱地区的地表水平衡,因此吸湿凝结水频率的大幅下降可能使得干旱区对全球变暖做出“更干燥的反应”[99].最近的研究指出,吸湿凝结水对未来气候变暖有显著响应,并强调在气候变暖条件下,微生境条件和植物特性共同影响吸湿凝结水的形成,对未来的生态系统过程具有重要的潜在作用[100].中国腾格里沙漠南缘的全球变暖长期模拟试验结果表明,长期增温和减雨使得藓类结皮的盖度显著降低,从而导致凝结水数量的减少[101]. ...
Dew formation reduction in global warming experiments and the potential consequences
1
2021
... 在最近的全球变暖期间,中国西北干旱地区的地表变暖大于湿润和半湿润地区,这导致地表相对湿度的下降幅度更大,进一步会导致吸湿凝结水的生成频率降低.中国597个连续观测站点的研究(1961—2010年)表明,由于近地层变暖和相对湿度降低,凝结水的生成频率每10年减少了5.2天,与20世纪60年代相比,21世纪初的凝结水生成频率在湿润、半湿润(降雨量为400~800 mm/a)和干旱区分别下降了28%、40%和50%.在地中海地区的研究也表明,未来数十年内,随着全球变暖,旱地的气温升高,蒸发增加,吸湿凝结水的数量和间期都可能减少[98].由于吸湿凝结水极大地促进了干旱地区的地表水平衡,因此吸湿凝结水频率的大幅下降可能使得干旱区对全球变暖做出“更干燥的反应”[99].最近的研究指出,吸湿凝结水对未来气候变暖有显著响应,并强调在气候变暖条件下,微生境条件和植物特性共同影响吸湿凝结水的形成,对未来的生态系统过程具有重要的潜在作用[100].中国腾格里沙漠南缘的全球变暖长期模拟试验结果表明,长期增温和减雨使得藓类结皮的盖度显著降低,从而导致凝结水数量的减少[101]. ...
Hydrological response of biological soil crusts to global warming: a ten-year simulative study
1
2018
... 在最近的全球变暖期间,中国西北干旱地区的地表变暖大于湿润和半湿润地区,这导致地表相对湿度的下降幅度更大,进一步会导致吸湿凝结水的生成频率降低.中国597个连续观测站点的研究(1961—2010年)表明,由于近地层变暖和相对湿度降低,凝结水的生成频率每10年减少了5.2天,与20世纪60年代相比,21世纪初的凝结水生成频率在湿润、半湿润(降雨量为400~800 mm/a)和干旱区分别下降了28%、40%和50%.在地中海地区的研究也表明,未来数十年内,随着全球变暖,旱地的气温升高,蒸发增加,吸湿凝结水的数量和间期都可能减少[98].由于吸湿凝结水极大地促进了干旱地区的地表水平衡,因此吸湿凝结水频率的大幅下降可能使得干旱区对全球变暖做出“更干燥的反应”[99].最近的研究指出,吸湿凝结水对未来气候变暖有显著响应,并强调在气候变暖条件下,微生境条件和植物特性共同影响吸湿凝结水的形成,对未来的生态系统过程具有重要的潜在作用[100].中国腾格里沙漠南缘的全球变暖长期模拟试验结果表明,长期增温和减雨使得藓类结皮的盖度显著降低,从而导致凝结水数量的减少[101]. ...
Dew deposition suppresses transpiration and carbon uptake in leaves
1
2018
... BSCs内部的生物除了会直接吸收吸湿凝结水外,一个更普遍的现象是吸湿凝结水在BSCs表层的形成影响地表的能量平衡,因为吸湿凝结水会增加表层的反照率和扩散率,通过蒸发降低表层温度.对美国加利福尼亚州热带雨林中植物的模型模拟研究表明,对于长度50 cm的叶片,凝结水的蒸腾抑制可显著减少25%的水分丢失,除去在风速低于1 m/s的条件下,较大叶片的CO2同化率有升高的可能,其他叶片的CO2同化率降低了12%[102].Ouyang等[103]的研究表明,尽管不同种类BSCs利用非降雨水分的能力不同,但绝大多数BSCs都可以利用非降雨水分进行碳固定,且非降雨水形成的夜间BSCs的碳交换特征与日间明显不同,日间光照较强时,增温通常能够导致BSCs光合作用降低而呼吸增强,在非降雨水形成的夜间,光照弱,此时的变暖对光合作用的增加比呼吸作用更强.在未来全球气候变化的条件下,吸湿凝结水的生成频率和数量都会发生变化,这一变化会对BSCs的水分状态、生长发育及荒漠生态系统的碳、水、能量平衡产生什么结果还未可知.Rodriguez-Caballero等[104]通过观测和环境模拟估计了BSCs的全球分布特征及对未来气候变化的响应,认为当前覆盖了陆地表层约12%的BSCs,在未来65年内会由于气候变化和土地利用方式的改变而降低25%~40%,对气候变化的响应比维管束植物要强烈的多.吸湿凝结水和BSCs盖度的联合变化必然影响土壤水分的可利用性和碳流,将对旱地功能产生不可预计的结果,在中长期尺度上可能影响全球碳水平衡. ...
Insight into climate change from the carbon exchange of biocrusts utilizing non-rainfall water
1
2017
... BSCs内部的生物除了会直接吸收吸湿凝结水外,一个更普遍的现象是吸湿凝结水在BSCs表层的形成影响地表的能量平衡,因为吸湿凝结水会增加表层的反照率和扩散率,通过蒸发降低表层温度.对美国加利福尼亚州热带雨林中植物的模型模拟研究表明,对于长度50 cm的叶片,凝结水的蒸腾抑制可显著减少25%的水分丢失,除去在风速低于1 m/s的条件下,较大叶片的CO2同化率有升高的可能,其他叶片的CO2同化率降低了12%[102].Ouyang等[103]的研究表明,尽管不同种类BSCs利用非降雨水分的能力不同,但绝大多数BSCs都可以利用非降雨水分进行碳固定,且非降雨水形成的夜间BSCs的碳交换特征与日间明显不同,日间光照较强时,增温通常能够导致BSCs光合作用降低而呼吸增强,在非降雨水形成的夜间,光照弱,此时的变暖对光合作用的增加比呼吸作用更强.在未来全球气候变化的条件下,吸湿凝结水的生成频率和数量都会发生变化,这一变化会对BSCs的水分状态、生长发育及荒漠生态系统的碳、水、能量平衡产生什么结果还未可知.Rodriguez-Caballero等[104]通过观测和环境模拟估计了BSCs的全球分布特征及对未来气候变化的响应,认为当前覆盖了陆地表层约12%的BSCs,在未来65年内会由于气候变化和土地利用方式的改变而降低25%~40%,对气候变化的响应比维管束植物要强烈的多.吸湿凝结水和BSCs盖度的联合变化必然影响土壤水分的可利用性和碳流,将对旱地功能产生不可预计的结果,在中长期尺度上可能影响全球碳水平衡. ...
Dryland photoautotrophic soil surface communities endangered by global change
1
2018
... BSCs内部的生物除了会直接吸收吸湿凝结水外,一个更普遍的现象是吸湿凝结水在BSCs表层的形成影响地表的能量平衡,因为吸湿凝结水会增加表层的反照率和扩散率,通过蒸发降低表层温度.对美国加利福尼亚州热带雨林中植物的模型模拟研究表明,对于长度50 cm的叶片,凝结水的蒸腾抑制可显著减少25%的水分丢失,除去在风速低于1 m/s的条件下,较大叶片的CO2同化率有升高的可能,其他叶片的CO2同化率降低了12%[102].Ouyang等[103]的研究表明,尽管不同种类BSCs利用非降雨水分的能力不同,但绝大多数BSCs都可以利用非降雨水分进行碳固定,且非降雨水形成的夜间BSCs的碳交换特征与日间明显不同,日间光照较强时,增温通常能够导致BSCs光合作用降低而呼吸增强,在非降雨水形成的夜间,光照弱,此时的变暖对光合作用的增加比呼吸作用更强.在未来全球气候变化的条件下,吸湿凝结水的生成频率和数量都会发生变化,这一变化会对BSCs的水分状态、生长发育及荒漠生态系统的碳、水、能量平衡产生什么结果还未可知.Rodriguez-Caballero等[104]通过观测和环境模拟估计了BSCs的全球分布特征及对未来气候变化的响应,认为当前覆盖了陆地表层约12%的BSCs,在未来65年内会由于气候变化和土地利用方式的改变而降低25%~40%,对气候变化的响应比维管束植物要强烈的多.吸湿凝结水和BSCs盖度的联合变化必然影响土壤水分的可利用性和碳流,将对旱地功能产生不可预计的结果,在中长期尺度上可能影响全球碳水平衡. ...




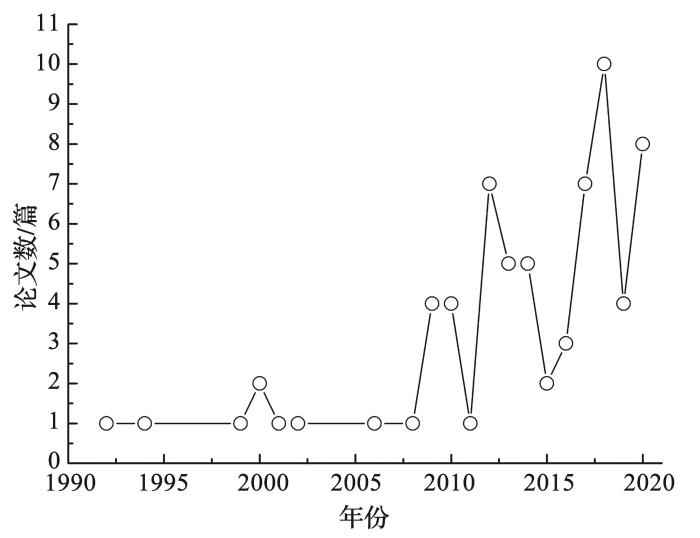

{kind=link}
{kind=link}