The evolution and future of Earth’s nitrogen cycle
1
2010
... 氮元素在地壳中的含量虽然不高,却是生命活动的必需元素,是组成氨基酸、酶、蛋白质、核酸等生物分子的重要成分.研究表明生物每结合100个碳原子,就需要同时结合2~20个氮原子[1 ] ,碳、氮循环的过程密不可分,是生命活动和地球气候系统的主要调控因素.自然界中氮有2种稳定同位素14 N和15 N,丰度分别为99.634%和0.366%.单质分子氮气是大气中氮的主要存在方式(占氮库99%以上),同时自然界中的氮化合物类型丰富,既有氧化态的硝酸盐(硝态氮,NO3 - ) 、亚硝酸盐(亚硝态氮,NO2 - ) ,也有还原态的铵离子(NH4 + ) 和气态氨分子(NH3 )以及有机氮(Organic Nitrogen,ON)等形式,各种形态的氮化合物化学价态变化较大(-3价~+5价),不同价态间的相互转变会产生显著的同位素分馏(-10‰~30‰)(图1 ). ...
Nitrogen stable isotopes in marine environments
1
2008
... 氮元素在地壳中的含量虽然不高,却是生命活动的必需元素,是组成氨基酸、酶、蛋白质、核酸等生物分子的重要成分.研究表明生物每结合100个碳原子,就需要同时结合2~20个氮原子
[1 ] ,碳、氮循环的过程密不可分,是生命活动和地球气候系统的主要调控因素.自然界中氮有2种稳定同位素
14 N和
15 N,丰度分别为99.634%和0.366%.单质分子氮气是大气中氮的主要存在方式(占氮库99%以上),同时自然界中的氮化合物类型丰富,既有氧化态的硝酸盐(硝态氮,NO
3 - ) 、亚硝酸盐(亚硝态氮,NO
2 - ) ,也有还原态的铵离子(NH
4 + ) 和气态氨分子(NH
3 )以及有机氮(Organic Nitrogen,ON)等形式,各种形态的氮化合物化学价态变化较大(-3价~+5价),不同价态间的相互转变会产生显著的同位素分馏(-10‰~30‰)(
图1 ).
图1 海洋环境中的典型稳定氮同位素变化(据参考文献[<xref ref-type="bibr" rid="R2">2</xref>,<xref ref-type="bibr" rid="R3">3</xref>,<xref ref-type="bibr" rid="R4">4</xref>,<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>]修改) Typical stable nitrogen isotope changes in the marine environment (modified after references [2~6]) Fig. 1 ![]()
海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
Inverse kinetic isotope fractionation during bacterial nitrite oxidation
1
2009
... 氮元素在地壳中的含量虽然不高,却是生命活动的必需元素,是组成氨基酸、酶、蛋白质、核酸等生物分子的重要成分.研究表明生物每结合100个碳原子,就需要同时结合2~20个氮原子
[1 ] ,碳、氮循环的过程密不可分,是生命活动和地球气候系统的主要调控因素.自然界中氮有2种稳定同位素
14 N和
15 N,丰度分别为99.634%和0.366%.单质分子氮气是大气中氮的主要存在方式(占氮库99%以上),同时自然界中的氮化合物类型丰富,既有氧化态的硝酸盐(硝态氮,NO
3 - ) 、亚硝酸盐(亚硝态氮,NO
2 - ) ,也有还原态的铵离子(NH
4 + ) 和气态氨分子(NH
3 )以及有机氮(Organic Nitrogen,ON)等形式,各种形态的氮化合物化学价态变化较大(-3价~+5价),不同价态间的相互转变会产生显著的同位素分馏(-10‰~30‰)(
图1 ).
图1 海洋环境中的典型稳定氮同位素变化(据参考文献[<xref ref-type="bibr" rid="R2">2</xref>,<xref ref-type="bibr" rid="R3">3</xref>,<xref ref-type="bibr" rid="R4">4</xref>,<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>]修改) Typical stable nitrogen isotope changes in the marine environment (modified after references [2~6]) Fig. 1 ![]()
海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
Nitrogen isotope effects induced by anammox bacteria
2
2013
... 氮元素在地壳中的含量虽然不高,却是生命活动的必需元素,是组成氨基酸、酶、蛋白质、核酸等生物分子的重要成分.研究表明生物每结合100个碳原子,就需要同时结合2~20个氮原子
[1 ] ,碳、氮循环的过程密不可分,是生命活动和地球气候系统的主要调控因素.自然界中氮有2种稳定同位素
14 N和
15 N,丰度分别为99.634%和0.366%.单质分子氮气是大气中氮的主要存在方式(占氮库99%以上),同时自然界中的氮化合物类型丰富,既有氧化态的硝酸盐(硝态氮,NO
3 - ) 、亚硝酸盐(亚硝态氮,NO
2 - ) ,也有还原态的铵离子(NH
4 + ) 和气态氨分子(NH
3 )以及有机氮(Organic Nitrogen,ON)等形式,各种形态的氮化合物化学价态变化较大(-3价~+5价),不同价态间的相互转变会产生显著的同位素分馏(-10‰~30‰)(
图1 ).
图1 海洋环境中的典型稳定氮同位素变化(据参考文献[<xref ref-type="bibr" rid="R2">2</xref>,<xref ref-type="bibr" rid="R3">3</xref>,<xref ref-type="bibr" rid="R4">4</xref>,<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>]修改) Typical stable nitrogen isotope changes in the marine environment (modified after references [2~6]) Fig. 1 ![]()
海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... 在沿海近岸地区,反硝化作用在脱氮过程中占主导作用,但在开放大洋中,厌氧氨氧化在脱氮过程发挥的作用甚至超过反硝化,改变了传统观念中反硝化作用为唯一氮素移除途径的认识.富含有机质的厌氧或低氧沉积物也是厌氧氨氧化的典型生境.据估计海洋的厌氧氨氧化过程可占到全球海洋脱氮过程一半左右[26 ,27 ] ,该过程中的氮同位素效应叠加在低氧区硝酸盐还原为亚硝酸盐的氮同位素分馏作用上,可能增强了氧气最小值区域(Oxygen-Minimum Zone, OMZ)氮去除过程中的总氮同位素效应,因而,在OMZ水域中硝酸盐的15 N富集(>10‰)和溶解N2 的15 N耗竭(-0.6‰)也可能与厌氧氨氧化过程相关,而不仅仅局限于水柱反硝化有关的氮同位素效应[4 ] . ...
N and O isotope effects during nitrate assimilation by unicellular prokaryotic and eukaryotic plankton cultures
1
2010
... 氮元素在地壳中的含量虽然不高,却是生命活动的必需元素,是组成氨基酸、酶、蛋白质、核酸等生物分子的重要成分.研究表明生物每结合100个碳原子,就需要同时结合2~20个氮原子
[1 ] ,碳、氮循环的过程密不可分,是生命活动和地球气候系统的主要调控因素.自然界中氮有2种稳定同位素
14 N和
15 N,丰度分别为99.634%和0.366%.单质分子氮气是大气中氮的主要存在方式(占氮库99%以上),同时自然界中的氮化合物类型丰富,既有氧化态的硝酸盐(硝态氮,NO
3 - ) 、亚硝酸盐(亚硝态氮,NO
2 - ) ,也有还原态的铵离子(NH
4 + ) 和气态氨分子(NH
3 )以及有机氮(Organic Nitrogen,ON)等形式,各种形态的氮化合物化学价态变化较大(-3价~+5价),不同价态间的相互转变会产生显著的同位素分馏(-10‰~30‰)(
图1 ).
图1 海洋环境中的典型稳定氮同位素变化(据参考文献[<xref ref-type="bibr" rid="R2">2</xref>,<xref ref-type="bibr" rid="R3">3</xref>,<xref ref-type="bibr" rid="R4">4</xref>,<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>]修改) Typical stable nitrogen isotope changes in the marine environment (modified after references [2~6]) Fig. 1 ![]()
海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
Amino acid composition and δ15 N of suspended matter in the Arabian Sea: Implications for organic matter sources and degradation
3
2013
... 氮元素在地壳中的含量虽然不高,却是生命活动的必需元素,是组成氨基酸、酶、蛋白质、核酸等生物分子的重要成分.研究表明生物每结合100个碳原子,就需要同时结合2~20个氮原子
[1 ] ,碳、氮循环的过程密不可分,是生命活动和地球气候系统的主要调控因素.自然界中氮有2种稳定同位素
14 N和
15 N,丰度分别为99.634%和0.366%.单质分子氮气是大气中氮的主要存在方式(占氮库99%以上),同时自然界中的氮化合物类型丰富,既有氧化态的硝酸盐(硝态氮,NO
3 - ) 、亚硝酸盐(亚硝态氮,NO
2 - ) ,也有还原态的铵离子(NH
4 + ) 和气态氨分子(NH
3 )以及有机氮(Organic Nitrogen,ON)等形式,各种形态的氮化合物化学价态变化较大(-3价~+5价),不同价态间的相互转变会产生显著的同位素分馏(-10‰~30‰)(
图1 ).
图1 海洋环境中的典型稳定氮同位素变化(据参考文献[<xref ref-type="bibr" rid="R2">2</xref>,<xref ref-type="bibr" rid="R3">3</xref>,<xref ref-type="bibr" rid="R4">4</xref>,<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>]修改) Typical stable nitrogen isotope changes in the marine environment (modified after references [2~6]) Fig. 1 ![]()
海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... 固氮作用的强度受到很多环境因子的限制,除普遍认可的大气风尘沉降的铁的可用性这一重要限制因素外[36 ,37 ] ,海表温度(海表太阳辐射量)也被认为是造成氮固定空间变化的主导因素[38 ] .传统认为上升流区域的某些条件(例如,海水温度较低)不利于氮固定,但在热带、亚热带上升流区(例如南海西北部夏季上升流区),越来越多的野外观测数据表明固氮作用的发生,且沿岸上升流作用似乎使该地区的氮固定不受温度或铁的限制[39 ] .也有学者认为贫营养海区混合层及深海中的颗粒物有可能不是受氮源影响,而是由滞留时间较长的细颗粒物及抗降解的有机质控制[6 ] . ...
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Dissolved organic nitrogen: A dynamic participant in aquatic ecosystems
1
2003
... 海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
Marine nitrogen cycle recorded by nitrogen and oxygen isotope fractionation of nitrate
3
2013
... 海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... [8 ,9 ].同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
硝酸盐氮氧稳定同位素分馏过程记录的海洋氮循环研究进展
3
2013
... 海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... [8 ,9 ].同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
Isotopic tracers of the marine nitrogen cycle: Present and past
1
2007
... 海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
The dynamics of the marine nitrogen cycle across the last deglaciation
2
2013
... 海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
The acceleration of oceanic denitrification during deglacial warming
4
2013
... 海水中的氮主要以NO3 - 4 + 2 - 3 - [7 ] .海洋中的氮循环是一个复杂多变的体系,主要与一系列微生物介导的生物过程密切联系,如固氮作用、反硝化作用、同化作用、硝化作用、矿化作用和厌氧氨氧化等[8 ] .仅靠传统的浓度测量难以准确计算海洋氮的收支、转化.但不同来源的氮物质因为形成方式不同具有特殊的同位素特征(图1 ),可以用来判断氮循环中生物过程变化的类型和强度,有力地推动了对海洋氮循环及其关键过程的研究[8 ,9 ] .同时氮同位素分馏会保存在地质记录中,是追溯古海洋氮循环变化以及古环境气候的重要参数,近年来已经取得了许多重要进展.但海洋环境的时间、空间的复杂多变使得海洋稳定氮同位素受到多重因素的共同影响,不仅是氮循环过程,其他生物、物理、化学过程都会不同程度地影响氮同位素的分馏.因此,海洋δ 15 N既受预先形成的水团δ 15 N影响,也受沿次表层水流路径变化导致的氮变化影响,次表层的原始信号也会因为表层新固定氮的添加或局部硝酸盐的消耗而改变[10 ] .除此以外,大洋环流也是一个永久的残余硝酸盐的再分配过程,通过海水混合和平流,一个海域组分的吸收会对其他海域硝酸盐δ 15 N产生影响,使得海洋氮的生物地球化学研究具有复杂性和多重不确定性[11 ] . ...
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(δ 15 Nbulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物δ 15 Nbulk 和上覆海水下透光带的硝酸盐δ 15 N-NO3 - 图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大[25 ] .Galbraith等[11 ] 汇聚全球现代海底样品δ 15 N数据结果表明,现代海底沉积物的δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物δ 15 N高,固氮作用显著的海域则相反(图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(δ 15 Nbulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其δ 15 Nbulk <5‰,主要位于固氮作用强的海域,因此δ 15 Nbulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征 [50 ,51 ,52 ,53 ] . ...
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
... 整体而言,在冰期—间冰期旋回中,全球温跃层反硝化速率发生了明显一致的变化,但局部变化受到区域环流和生产力波动的调节.考虑到全球同位素质量平衡,末次盛冰期(Last Glacial Maximum,LGM)氮循环接近稳定状态,冰消期水柱反硝化作用加强,而在全新世早期反硝化作用再次接近稳定状态.依据现代海底反硝化数据估算出在15~8 ka阶段底栖反硝化增加了30%~120%.根据全球平均δ 15 N在末次盛冰期和全新世早期的相似性,推断水柱反硝化作用在两种稳定状态之间有相似的增加量[11 ] . ...
Nitrogen cycling in maine sediments
1
2001
... 由于氮是海洋环境中的限制性营养元素,表层海水的硝酸盐常常被完全利用,因此生物有机质δ 15 N大多反映了海洋硝酸盐δ 15 N特征,大洋中一般为5‰~7‰,在上升流地区可达17‰.在整个海洋氮循环中,浮游生物吸收海水中的营养盐并固定在有机物中,最终通过扩散或沉降运输到达海底形成表层沉积物并逐渐堆积,在这个过程中会发生一系列生物地球化学反应(如水解、微生物分解降解或者再矿化作用等),其中大部分有机氮还原成可被生产者利用的无机形式重新进入营养循环,仅小部分沉降到海底并随成岩作用埋藏于海底,短期内不再参与循环[12 ] .当生物体吸收各种来源的氮盐的同时,其相应的同位素特征也被保存在有机质中,海洋中真光层氮同位素组成可被储存在水体颗粒物中,并最终埋藏在沉积物中.海洋颗粒物从生成到埋藏经历诸多复杂过程,理解海洋硝酸盐及颗粒物稳定氮同位素的信号对充分了解海洋中沉降过程中的控制因素至关重要.因此通过研究海洋颗粒物中的氮同位素组成示踪颗粒有机物的来源、生物地球化学过程,对深刻理解现代海洋氮循环过程及机制有重要意义. ...
海洋沉积物中的氮循环
1
2001
... 由于氮是海洋环境中的限制性营养元素,表层海水的硝酸盐常常被完全利用,因此生物有机质δ 15 N大多反映了海洋硝酸盐δ 15 N特征,大洋中一般为5‰~7‰,在上升流地区可达17‰.在整个海洋氮循环中,浮游生物吸收海水中的营养盐并固定在有机物中,最终通过扩散或沉降运输到达海底形成表层沉积物并逐渐堆积,在这个过程中会发生一系列生物地球化学反应(如水解、微生物分解降解或者再矿化作用等),其中大部分有机氮还原成可被生产者利用的无机形式重新进入营养循环,仅小部分沉降到海底并随成岩作用埋藏于海底,短期内不再参与循环[12 ] .当生物体吸收各种来源的氮盐的同时,其相应的同位素特征也被保存在有机质中,海洋中真光层氮同位素组成可被储存在水体颗粒物中,并最终埋藏在沉积物中.海洋颗粒物从生成到埋藏经历诸多复杂过程,理解海洋硝酸盐及颗粒物稳定氮同位素的信号对充分了解海洋中沉降过程中的控制因素至关重要.因此通过研究海洋颗粒物中的氮同位素组成示踪颗粒有机物的来源、生物地球化学过程,对深刻理解现代海洋氮循环过程及机制有重要意义. ...
An Earth-system perspective of the global nitrogen cycle
3
2008
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
... 沉积物中有机物稳定氮同位素变化同样是了解地质历史上海洋氮循环变化的有效手段,虽然其作为海洋氮动力学记录的保真度仍有疑问,但通过结合其他指标对不同沉积状态记录进行对比可以更好地约束环境参数,在重建古海洋古气候方面取得了显著的进展.事实上,迄今为止许多δ 15 N记录已显示了不同空间和时间尺度上的相互关联,证明δ 15 N记录与全球气候变化的紧密联系[13 ,66 ,67 ] . ...
Managing soil denitrification
1
2002
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
Nitrogen isotopic evidence for micronutrient control of fractional NO3 -
2
2001
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O
2 浓度小于5
μ mol/L)
[13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件
[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域
[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除
14 N硝酸盐导致剩余硝酸盐的
δ 15 N明显升高(
图2 a),而释放出贫重同位素的N
2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境
[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的
δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(
图2 b),主要受控于该区上涌的硝酸盐
δ 15 N(
δ 15 NO
3 - 约为11‰)影响,因而这些地区的稳定氮同位素变化可用于重建该区上升流信号
[22 ] .
图2 不同海域垂向硝酸盐含量(空心圆圈)、硝酸盐(实心圆圈)及颗粒物(蓝色实心三角)稳定氮同位素变化模式(据参考文献[<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R16">16</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R18">18</xref>]修改) Vertical variability of nitrate content (hollow circle) and stable nitrogen isotope of nitrate (solid circle) and sinking particles (blue solid triangle) in different sea areas (modified after references [15~18]) Fig. 2 ![]()
大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
... 而在高营养低叶绿素海区(High-Nutrient Low-Chlorophyll,HNLC),例如亚北极太平洋、东赤道太平洋和南大洋等,表层硝酸盐没有完全被利用(图2 c),主要受到同化作用的影响.生物优先吸收轻同位素营养盐,沉降颗粒物δ 15 N比海水δ 15 N-NO3 - δ 15 N相对偏轻,海底表层沉积物δ 15 N同样受到影响,低于表层硝酸盐δ 15 N,因此在极地和东赤道太平洋中颗粒物δ 15 N的空间分布模式反映了上层硝酸盐的利用情况[15 ] .另外在超富营养海区还能观察到硝化作用的影响,使海水NH4 + 15 N富集.一般大洋中的NH4 + 3 - δ 15 N,被认为与该区域的不完全硝化作用有关,使得生物吸收同位素正偏的剩余NH4 + δ 15 N变重[28 ] . ...
A bacterial method for the nitrogen isotopic analysis of nitrate in seawater and freshwater
2
2001
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O
2 浓度小于5
μ mol/L)
[13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件
[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域
[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除
14 N硝酸盐导致剩余硝酸盐的
δ 15 N明显升高(
图2 a),而释放出贫重同位素的N
2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境
[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的
δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(
图2 b),主要受控于该区上涌的硝酸盐
δ 15 N(
δ 15 NO
3 - 约为11‰)影响,因而这些地区的稳定氮同位素变化可用于重建该区上升流信号
[22 ] .
图2 不同海域垂向硝酸盐含量(空心圆圈)、硝酸盐(实心圆圈)及颗粒物(蓝色实心三角)稳定氮同位素变化模式(据参考文献[<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R16">16</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R18">18</xref>]修改) Vertical variability of nitrate content (hollow circle) and stable nitrogen isotope of nitrate (solid circle) and sinking particles (blue solid triangle) in different sea areas (modified after references [15~18]) Fig. 2 ![]()
大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
. Examining N cycling in the northern South China Sea from N isotopic signals in nitrate and particulate phases
2
2017
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O
2 浓度小于5
μ mol/L)
[13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件
[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域
[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除
14 N硝酸盐导致剩余硝酸盐的
δ 15 N明显升高(
图2 a),而释放出贫重同位素的N
2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境
[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的
δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(
图2 b),主要受控于该区上涌的硝酸盐
δ 15 N(
δ 15 NO
3 - 约为11‰)影响,因而这些地区的稳定氮同位素变化可用于重建该区上升流信号
[22 ] .
图2 不同海域垂向硝酸盐含量(空心圆圈)、硝酸盐(实心圆圈)及颗粒物(蓝色实心三角)稳定氮同位素变化模式(据参考文献[<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R16">16</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R18">18</xref>]修改) Vertical variability of nitrate content (hollow circle) and stable nitrogen isotope of nitrate (solid circle) and sinking particles (blue solid triangle) in different sea areas (modified after references [15~18]) Fig. 2 ![]()
大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
... 沉积物中的无机组分对于稳定氮同位素的研究也有很大的影响,无机氮是沉积物总氮中的重要组成部分,含量从40%(亚热带大西洋沉积物中)[62 ] 到约60%不等(北冰洋、亚北极西部太平洋,中国南海沉积物中)[17 ,63 ,64 ] .高无机氮组分会导致沉积物稳定氮同位素呈现完全不同的特征,造成C/N和δ 15 Nbulk 偏移,使用KOH-KOBr处理去除有机氮间接获得有机氮数据或者直接酸处理去除无机氮的影响[65 ] .快速沉积环境中的大量沉积物基质可能含有许多不稳定碳以及与矿物相关的无机氮,特别是高矿物质但低有机物的环境,物种差异及其同位素组成对沉积物稳定氮同位素分析都有重要影响. ...
Stable isotope dynamics in elasmobranch fishes
2
2010
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O
2 浓度小于5
μ mol/L)
[13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件
[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域
[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除
14 N硝酸盐导致剩余硝酸盐的
δ 15 N明显升高(
图2 a),而释放出贫重同位素的N
2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境
[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的
δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(
图2 b),主要受控于该区上涌的硝酸盐
δ 15 N(
δ 15 NO
3 - 约为11‰)影响,因而这些地区的稳定氮同位素变化可用于重建该区上升流信号
[22 ] .
图2 不同海域垂向硝酸盐含量(空心圆圈)、硝酸盐(实心圆圈)及颗粒物(蓝色实心三角)稳定氮同位素变化模式(据参考文献[<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R16">16</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R18">18</xref>]修改) Vertical variability of nitrate content (hollow circle) and stable nitrogen isotope of nitrate (solid circle) and sinking particles (blue solid triangle) in different sea areas (modified after references [15~18]) Fig. 2 ![]()
大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
The nitrogen isotope biogeochemistry of sinking particles from the margin of the Eastern North Pacific
3
1999
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
... 卡里亚科海盆(蓝色)沉积物δ 15 Nbulk [79 ] ;阿拉伯海(棕红)沉积物δ 15 Nbulk [19 ] ;南海沉积物δ 15 Nbulk (黑色)[86 ] 和FB-δ 15 N(绿色)[89 ] ;阴影部分为间冰期,数字为海洋同位素阶段 ...
... Cariaco basin (blue) sediments δ 15 Nbulk [79 ] ; Arabian sea (brown red) sediments δ 15 Nbulk [19 ] The South China Sea sediments δ 15 Nbulk (black)[86 ] and FB-δ 15 N (green)[89 ] . The shaded part is the interglacial period, and the figure is the Marine Isotope Stage(MIS) ...
Nitrogen isotope patterns in the oxygen-deficient waters of the Eastern Tropical North Pacific Ocean
1
2001
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
The distribution of nitrate 15 N/14 N in marine sediments and the impact of benthic nitrogen loss on the isotopic composition of oceanic nitrate
1
2007
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
Amino acid composition and δ15 N of suspended matter in the Arabian Sea: Implications for organic matter sources and degradation
1
2013
... 海水颗粒物稳定氮同位素的区域差异与当地海洋环境的物理水文条件有关,主要受垂向混合控制的次表层硝酸盐的供给控制.在大洋低氧水层和高生产力沿海陆架沉积物等环境,主要发生反硝化细菌介导的反硝化作用.水柱反硝化作用主要受海水缺氧程度的影响,普遍发生在水柱缺氧区(O2 浓度小于5 μ [13 ] ,充足的有机质以及硝酸盐也是发生反硝化作用的重要条件[14 ] .典型的反硝化海域主要有阿拉伯海上升流区、热带东太平洋上升流区和秘鲁上升流区等高表层生产力的低氧海域[8 ] .反硝化作用会引起显著的同位素分馏(25‰~30‰),该过程优先去除14 N硝酸盐导致剩余硝酸盐的δ 15 N明显升高(图2 a),而释放出贫重同位素的N2 ,因此海洋硝酸盐的同位素组成可用于鉴定反硝化环境[19 ,20 ,21 ] .上升流活跃海区的表层海水颗粒物的δ 15 N一般偏重,如阿拉伯海为8.7‰~9.2‰,显著高于非上升流影响的南海(4‰~5‰)(图2 b),主要受控于该区上涌的硝酸盐δ 15 N(δ 15 NO3 - [22 ] . ...
Sinking organic matter spreads the nitrogen isotope signal of pelagic denitrification in the North Pacific
1
2009
... 大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
The δ15 N of nitrate in the Southern Ocean: Nitrogen cycling and circulation in the ocean interior
1
2000
... 大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
Nitrogen isotope dynamics of the Cariaco Basin, Venezuela
2
2004
... 大洋海水硝酸盐的同位素平均为5‰(变化范围为3‰~14‰),中层水体在弱氧水柱中通过反硝化作用使残余硝酸盐δ 15 N富集,因而在反硝化作用强的海域中,部分水层中硝酸盐的同位素可达20‰[23 ] ,造成浅层和中层水体中硝酸盐的δ 15 N与深海硝酸盐的平均偏差为4.8‰±0.2‰,该同位素差值可以反映与同位素分馏有关的海洋氮供应和转化过程[24 ] .但卡里亚科(Cariaco)海盆由于水柱氧化还原界面的反硝化作用几乎将水中的硝酸盐消耗完全,因此海盆的硝酸盐没有明显的同位素记录,反而导致反硝化同位素效应最弱[25 ] . ...
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(δ 15 Nbulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物δ 15 Nbulk 和上覆海水下透光带的硝酸盐δ 15 N-NO3 - 图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大[25 ] .Galbraith等[11 ] 汇聚全球现代海底样品δ 15 N数据结果表明,现代海底沉积物的δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物δ 15 N高,固氮作用显著的海域则相反(图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(δ 15 Nbulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其δ 15 Nbulk <5‰,主要位于固氮作用强的海域,因此δ 15 Nbulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征 [50 ,51 ,52 ,53 ] . ...
Recent advances in the biogeochemistry of nitrogen in the ocean
1
2008
... 在沿海近岸地区,反硝化作用在脱氮过程中占主导作用,但在开放大洋中,厌氧氨氧化在脱氮过程发挥的作用甚至超过反硝化,改变了传统观念中反硝化作用为唯一氮素移除途径的认识.富含有机质的厌氧或低氧沉积物也是厌氧氨氧化的典型生境.据估计海洋的厌氧氨氧化过程可占到全球海洋脱氮过程一半左右[26 ,27 ] ,该过程中的氮同位素效应叠加在低氧区硝酸盐还原为亚硝酸盐的氮同位素分馏作用上,可能增强了氧气最小值区域(Oxygen-Minimum Zone, OMZ)氮去除过程中的总氮同位素效应,因而,在OMZ水域中硝酸盐的15 N富集(>10‰)和溶解N2 的15 N耗竭(-0.6‰)也可能与厌氧氨氧化过程相关,而不仅仅局限于水柱反硝化有关的氮同位素效应[4 ] . ...
Production of N2 through Anaerobic Ammonium Oxidation Coupled to Nitrate Reduction in marine sediments
1
2002
... 在沿海近岸地区,反硝化作用在脱氮过程中占主导作用,但在开放大洋中,厌氧氨氧化在脱氮过程发挥的作用甚至超过反硝化,改变了传统观念中反硝化作用为唯一氮素移除途径的认识.富含有机质的厌氧或低氧沉积物也是厌氧氨氧化的典型生境.据估计海洋的厌氧氨氧化过程可占到全球海洋脱氮过程一半左右[26 ,27 ] ,该过程中的氮同位素效应叠加在低氧区硝酸盐还原为亚硝酸盐的氮同位素分馏作用上,可能增强了氧气最小值区域(Oxygen-Minimum Zone, OMZ)氮去除过程中的总氮同位素效应,因而,在OMZ水域中硝酸盐的15 N富集(>10‰)和溶解N2 的15 N耗竭(-0.6‰)也可能与厌氧氨氧化过程相关,而不仅仅局限于水柱反硝化有关的氮同位素效应[4 ] . ...
14 N in particulate organic matter, zooplankton, and diatom frustule-bound nitrogen in the ice-covered water column of the Bering Sea eastern shelf
1
2014
... 而在高营养低叶绿素海区(High-Nutrient Low-Chlorophyll,HNLC),例如亚北极太平洋、东赤道太平洋和南大洋等,表层硝酸盐没有完全被利用(图2 c),主要受到同化作用的影响.生物优先吸收轻同位素营养盐,沉降颗粒物δ 15 N比海水δ 15 N-NO3 - δ 15 N相对偏轻,海底表层沉积物δ 15 N同样受到影响,低于表层硝酸盐δ 15 N,因此在极地和东赤道太平洋中颗粒物δ 15 N的空间分布模式反映了上层硝酸盐的利用情况[15 ] .另外在超富营养海区还能观察到硝化作用的影响,使海水NH4 + 15 N富集.一般大洋中的NH4 + 3 - δ 15 N,被认为与该区域的不完全硝化作用有关,使得生物吸收同位素正偏的剩余NH4 + δ 15 N变重[28 ] . ...
Nitrogen sources in the South China Sea, as discerned from stable nitrogen isotopic ratios in rivers, sinking particles, and sediments
3
2009
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
... 由于海洋和陆地有机物δ 15 N的差异,陆源物质的输入也会导致海洋沉积物δ 15 N的变化.典型的海洋沉积物δ 15 N与大洋溶解硝酸盐(平均δ 15 N为5‰)相近.而陆地植物通过固氮菌吸收大气N2 进行光合作用,其δ 15 N一般较轻,往往小于5‰.因此沉积物δ 15 N从陆架区向深海区呈现增加趋势,在南海从4.2‰~5‰增加到6‰[29 ] ,在北印度洋从小于4‰增加到大于11‰[46 ] ,北安达曼海从小于4‰增加到大于5‰ [59 ] .同时近几十年来随着人类活动的增加,大气中大幅增加的含氮化合物通过干湿沉降到地表,导致湖泊和海洋富营养化.因此干湿沉降中的氮不仅有闪电、土壤释放等自然来源还有化石燃料燃烧等人类来源,前者δ 15 N一般很低(<-20‰),而后者影响因素复杂,δ 15 N有较大变化范围(-23.3‰~25.6‰)[60 ] .外源氮可能会影响和改变海洋氮库的平衡,总体上占比为12%~25%,在开放大洋中更低[61 ] .但陆源输入的氮同位素很难被界定,包括河流到海岸带过程中存在的不同形态的氮化合物以及各种反应,因此目前仍缺乏对陆源输入的氮的同位素组成进行系统研究. ...
N2 fixation variability in the oligotrophic Sulu Sea, western equatorial Pacific region over the past 83 kyr
4
2006
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
... [30 ,35 ]. ...
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
The evolution of Earth's biogeochemical nitrogen cycle
1
2016
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
Marine ecosystem dynamics and biogeochemical cycling in the community Earth system model [CESM1(BGC)]: Comparison of the 1990s with the 2090s under the RCP4.5 and RCP8.5 scenarios
3
2013
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
... [32 ,75 ,76 ,77 ].这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Dinitrogen fixation in the World's oceans
1
57
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
N isotopic composition of dissolved organic nitrogen and nitrate at the Bermuda Atlantic time-series study site
1
2005
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
Stable isotope constraints on the nitrogen cycle of the Mediterranean Sea water column
1
2002
... 在寡营养海域中,硝酸盐几乎被完全利用,水体颗粒物和沉积物中的δ 15 N主要反映了次表层和大气来源的新氮信号[29 ,30 ] .对这些海区而言,固氮作用是最重要的向真光层内提供生物可利用氮的途径,是海洋新生氮的主要外源,在表层海水中蓝藻的氮固定几乎主导了氮向海洋的输入[13 ] .固氮生物将N2 还原为NH4 + δ 15 N比基质低1‰),而在富Fe2+ 或嗜热培养中,分馏程度可达-4‰,用钒或铁代替钼的固氮酶时同位素分馏可达-4‰~-8‰[31 ] .固氮作用使海洋NO3 - δ 15 N较之大气中的氮气同位素偏负(0~-2‰),因此降低了海洋硝酸盐的平均δ 15 N[32 ] .在副热带海洋地区,温跃层比深海大洋的平均值低2‰~3‰,其原因与固氮作用直接有关,表层固氮作用合成的低同位素特征的有机氮的分解降低了水体中硝酸盐的δ 15 N[33 ,34 ] .在苏禄海和地中海等贫养区域,固氮生物的作用也使海洋颗粒物的δ 15 N显著低于次表层水的δ 15 N-NO3 - [30 ,35 ] . ...
Evolution of the nitrogen cycle and its influence on the biological sequestration of CO2 in the ocean
1
1997
... 固氮作用的强度受到很多环境因子的限制,除普遍认可的大气风尘沉降的铁的可用性这一重要限制因素外[36 ,37 ] ,海表温度(海表太阳辐射量)也被认为是造成氮固定空间变化的主导因素[38 ] .传统认为上升流区域的某些条件(例如,海水温度较低)不利于氮固定,但在热带、亚热带上升流区(例如南海西北部夏季上升流区),越来越多的野外观测数据表明固氮作用的发生,且沿岸上升流作用似乎使该地区的氮固定不受温度或铁的限制[39 ] .也有学者认为贫营养海区混合层及深海中的颗粒物有可能不是受氮源影响,而是由滞留时间较长的细颗粒物及抗降解的有机质控制[6 ] . ...
A unifying framework for dinitrogen fixation in the terrestrial biosphere
1
2008
... 固氮作用的强度受到很多环境因子的限制,除普遍认可的大气风尘沉降的铁的可用性这一重要限制因素外[36 ,37 ] ,海表温度(海表太阳辐射量)也被认为是造成氮固定空间变化的主导因素[38 ] .传统认为上升流区域的某些条件(例如,海水温度较低)不利于氮固定,但在热带、亚热带上升流区(例如南海西北部夏季上升流区),越来越多的野外观测数据表明固氮作用的发生,且沿岸上升流作用似乎使该地区的氮固定不受温度或铁的限制[39 ] .也有学者认为贫营养海区混合层及深海中的颗粒物有可能不是受氮源影响,而是由滞留时间较长的细颗粒物及抗降解的有机质控制[6 ] . ...
Data-based assessment of environmental controls on global marine nitrogen fixation
1
2013
... 固氮作用的强度受到很多环境因子的限制,除普遍认可的大气风尘沉降的铁的可用性这一重要限制因素外[36 ,37 ] ,海表温度(海表太阳辐射量)也被认为是造成氮固定空间变化的主导因素[38 ] .传统认为上升流区域的某些条件(例如,海水温度较低)不利于氮固定,但在热带、亚热带上升流区(例如南海西北部夏季上升流区),越来越多的野外观测数据表明固氮作用的发生,且沿岸上升流作用似乎使该地区的氮固定不受温度或铁的限制[39 ] .也有学者认为贫营养海区混合层及深海中的颗粒物有可能不是受氮源影响,而是由滞留时间较长的细颗粒物及抗降解的有机质控制[6 ] . ...
Physical-biological coupling of N2 fixation in the northwestern South China Sea coastal upwelling during summer
1
2015
... 固氮作用的强度受到很多环境因子的限制,除普遍认可的大气风尘沉降的铁的可用性这一重要限制因素外[36 ,37 ] ,海表温度(海表太阳辐射量)也被认为是造成氮固定空间变化的主导因素[38 ] .传统认为上升流区域的某些条件(例如,海水温度较低)不利于氮固定,但在热带、亚热带上升流区(例如南海西北部夏季上升流区),越来越多的野外观测数据表明固氮作用的发生,且沿岸上升流作用似乎使该地区的氮固定不受温度或铁的限制[39 ] .也有学者认为贫营养海区混合层及深海中的颗粒物有可能不是受氮源影响,而是由滞留时间较长的细颗粒物及抗降解的有机质控制[6 ] . ...
Nitrogen isotopic composition of sinking particles from the southern Bay of Bengal: Evidence for variable nitrogen sources
1
2006
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Tracing nitrogen sources and cycling in catchments
1
1998
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Nitrogen fixation and nitrogen isotope abundances in zooplankton of the oligotrophic north Atlantic
2
2002
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
... ,42 ,43 ].因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Seasonal and depth-related changes in the source of sinking particles in the North Atlantic
2
1991
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
... ,43 ].因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Isotope constraints on particulate nitrogen source and dynamics in the upper water column of the oligotrophic South China Sea
2
2012
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
... [44 ].而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
. Examining N cycling in the northern South China Sea from N isotopic signals in nitrate and particulate phases
1
2017
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Stable nitrogen isotopic ratios of sinking particles and sediments from the northern Indian Ocean
4
2005
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
... ,46 ,47 ,48 ].此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
... 由于海洋和陆地有机物δ 15 N的差异,陆源物质的输入也会导致海洋沉积物δ 15 N的变化.典型的海洋沉积物δ 15 N与大洋溶解硝酸盐(平均δ 15 N为5‰)相近.而陆地植物通过固氮菌吸收大气N2 进行光合作用,其δ 15 N一般较轻,往往小于5‰.因此沉积物δ 15 N从陆架区向深海区呈现增加趋势,在南海从4.2‰~5‰增加到6‰[29 ] ,在北印度洋从小于4‰增加到大于11‰[46 ] ,北安达曼海从小于4‰增加到大于5‰ [59 ] .同时近几十年来随着人类活动的增加,大气中大幅增加的含氮化合物通过干湿沉降到地表,导致湖泊和海洋富营养化.因此干湿沉降中的氮不仅有闪电、土壤释放等自然来源还有化石燃料燃烧等人类来源,前者δ 15 N一般很低(<-20‰),而后者影响因素复杂,δ 15 N有较大变化范围(-23.3‰~25.6‰)[60 ] .外源氮可能会影响和改变海洋氮库的平衡,总体上占比为12%~25%,在开放大洋中更低[61 ] .但陆源输入的氮同位素很难被界定,包括河流到海岸带过程中存在的不同形态的氮化合物以及各种反应,因此目前仍缺乏对陆源输入的氮的同位素组成进行系统研究. ...
Sensitivity of δ15 N of nitrate, surface suspended and deep sinking particulate nitrogen to seasonal nitrate depletion in the Southern Ocean
1
2003
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
Stable carbon and nitrogen isotopic compositions of sinking particles in the northeast Japan Sea
1
2003
... 由于氮循环和海洋环境的的复杂性,不同的海洋及生物地球化学过程会造成不同海区稳定氮同位素在垂向上的差异(图2 ).沉积物捕获器是研究沉降颗粒δ 15 N变化、识别和定量各种氮源以及复杂的海洋过程的有效方法,一般沉降较慢的小颗粒被称为悬浮颗粒物(suspended particle),主要在水体上层,而沉降较快的大颗粒被称为沉降颗粒物(sinking particle),主要在深层水体[40 ] .在大多数海域,海洋上层颗粒有机质(Particle Organic Matter,POM)在下沉到海底的过程中δ 15 NPOM 的升高意味着矿化作用的加剧.但一般情况下沉积物中有机质的δ 15 N信号与上方输入的δ 15 NPOM 一致,成岩作用的影响并不会改变有机质δ 15 N反映的来源特征[16 ,41 ] .Montoya等[42 ] 曾报道北大西洋悬浮颗粒物δ 15 N低于沉降颗粒物和浮游动物的δ 15 N,并且随颗粒物粒径增加而增大.同时,在北大西洋随海水深度加深,由于颗粒物中轻同位素分子优先降解和/或溶解,沉降颗粒物δ 15 N增加(图2 d),随深度增加氨基酸的矿化也会导致沉降颗粒物δ 15 N增加[43 ] .而近年在南海北部的一项研究说明真光层底部沉降颗粒物的δ 15 N接近次表层硝酸盐δ 15 N(4‰~5‰)(图2 b),同时悬浮颗粒物、沉降颗粒物以及不同粒径的浮游动物有相似的δ 15 N,说明研究区域不同颗粒组分间的快速氮交换[44 ] .南海颗粒稳定氮同位素信号在向海底传递的过程中有显著变化,上层0~100 m的颗粒物平均δ 15 N(2.0‰~5.3‰)低于下层100~200 m的,表现出明显的年际变化,很可能存在垂直分馏或/和大气输入的15 N亏损的外源氮[45 ] ;2 000~3 000 m的深水δ 15 N低于真光层的颗粒物和次表层硝酸盐的δ 15 N,结合3 000 m以下颗粒氮通量的高值说明可能存在由陆架区侧向输入带来含有贫15 N的陆源颗粒或海洋动力学作用下具有较低δ 15 N特征的单细胞原核生物对南海深层水的贡献[44 ] .而在阿拉伯海,随水柱深度的增加沉降颗粒物δ 15 N减少(达2‰)[46 ] .可能与较深的捕获器中具有低δ 15 N特征的细菌富集或来自陆地的有机成分的贡献有关;而优先保存同位素偏负的颗粒物质或较浅的捕获器中同位素较高的“游泳生物”的贡献也不能忽略[6 ,46 ,47 ,48 ] .此外,食物链对生物的稳定氮同位素也有重要影响,食物链每增加一级就会使生物的δ 15 N正偏2.0‰~3.5‰,导致有浮游生物存在的颗粒物的δ 15 N显著增加[18 ,42 ,43 ] .因此鉴于海洋环境的时空复杂性和氮生物地球化学过程的敏感性,要结合具体的海洋环境判识实际氮循环过程及同位素分馏影响机制. ...
A review of nitrogen isotopic alteration in marine sediments
4
2012
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(
δ 15 N
bulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的
δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物
δ 15 N
bulk 和上覆海水下透光带的硝酸盐
δ 15 N-NO
3 - 有良好的相关性(
图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大
[25 ] .Galbraith等
[11 ] 汇聚全球现代海底样品
δ 15 N数据结果表明,现代海底沉积物的
δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物
δ 15 N高,固氮作用显著的海域则相反(
图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(
δ 15 N
bulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其
δ 15 N
bulk <5‰,主要位于固氮作用强的海域,因此
δ 15 N
bulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征
[50 ,51 ,52 ,53 ] .
图3 全球不同大洋沉降颗粒(矩形)或硝酸盐(圆形)与表层沉积物的稳定氮同位素对比(据参考文献[<xref ref-type="bibr" rid="R49">49</xref>]修改) Stable nitrogen isotope comparison of sinking particles (rectangular) or nitrate (circular) with surface sediments from global oceans in different sites (modified after reference [<xref ref-type="bibr" rid="R49">49</xref>]) Fig. 3 ![]()
通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
... Stable nitrogen isotope comparison of sinking particles (rectangular) or nitrate (circular) with surface sediments from global oceans in different sites (modified after reference [
49 ])
Fig. 3 ![]()
通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
... 海洋沉积物受到复杂沉积环境的影响,如早期成岩作用,陆源输入和矿物组分等都会影响沉积物稳定氮同位素组成.海洋沉积速率或沉积物—水界面条件会随时间而变化,使得沉积物中稳定氮同位素随时间而存在成岩作用的富集,导致沉积物在早期埋藏阶段同位素的升高[49 ] ,早期成岩作用主要发生在表层10 cm左右,受有机质含量、氧含量以及底栖动物扰动等的影响,一般有机质含量越高成岩作用越显著,而底栖动物的扰动对矿化作用也有一定程度的促进作用.地中海作为世界上最古老的海域,在保存沉积物有机质(如交替的泥灰质和腐泥层)方面存在着显著的差异,在低有机碳的泥灰层具有高δ 15 Nbulk 而在有机质丰富的腐泥层δ 15 Nbulk 则较低.部分研究者认为该地区的δ 15 Nbulk 的原始数据已遭到了改变[57 ] ;而另有研究者认为大体上还是反映了原始的信号特征[58 ] .前者的主要依据是降解指数与δ 15 Nbulk 的线性关系,认为成岩作用导致的同位素蚀变提高了泥灰岩有机质δ 15 N,而腐泥层中有机质δ 15 Nbulk 较低是保存作用增强的结果[57 ] .后者主要是借助于δ 15 Nbulk 和绿素δ 15 N(δ 15 Nchlorin )的一致性证明两者的差异并不是由于后期改变而是表层海水生物地化条件的差异性造成的[58 ] ,因为δ 15 Nchlorin 主要反映了海洋浮游植物群落利用的初始海水营养盐δ 15 N的组成,受成岩作用的影响极小. ...
A global marine-fixed nitrogen isotopic budget: Implications for Holocene nitrogen cycling
1
2002
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(δ 15 Nbulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物δ 15 Nbulk 和上覆海水下透光带的硝酸盐δ 15 N-NO3 - 图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大[25 ] .Galbraith等[11 ] 汇聚全球现代海底样品δ 15 N数据结果表明,现代海底沉积物的δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物δ 15 N高,固氮作用显著的海域则相反(图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(δ 15 Nbulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其δ 15 Nbulk <5‰,主要位于固氮作用强的海域,因此δ 15 Nbulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征 [50 ,51 ,52 ,53 ] . ...
Isotopic constraints on glacial/interglacial changes in the oceanic nitrogen budget
1
2004
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(δ 15 Nbulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物δ 15 Nbulk 和上覆海水下透光带的硝酸盐δ 15 N-NO3 - 图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大[25 ] .Galbraith等[11 ] 汇聚全球现代海底样品δ 15 N数据结果表明,现代海底沉积物的δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物δ 15 N高,固氮作用显著的海域则相反(图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(δ 15 Nbulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其δ 15 Nbulk <5‰,主要位于固氮作用强的海域,因此δ 15 Nbulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征 [50 ,51 ,52 ,53 ] . ...
Nitrogen isotopic evidence for a poleward decrease in surface nitrate within the ice age Antarctic
1
2008
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(δ 15 Nbulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物δ 15 Nbulk 和上覆海水下透光带的硝酸盐δ 15 N-NO3 - 图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大[25 ] .Galbraith等[11 ] 汇聚全球现代海底样品δ 15 N数据结果表明,现代海底沉积物的δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物δ 15 N高,固氮作用显著的海域则相反(图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(δ 15 Nbulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其δ 15 Nbulk <5‰,主要位于固氮作用强的海域,因此δ 15 Nbulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征 [50 ,51 ,52 ,53 ] . ...
The polar ocean and glacial cycles in atmospheric CO2 concentration
1
2010
... 海洋表层沉积物是上层沉降颗粒物的长时间均匀混合而成,同样也可以反映上层海水营养盐变化情况,沉积物中总氮同位素(δ 15 Nbulk )常常作为海洋上层水体中氮循环替代指标.通过对悬浮颗粒物、沉降颗粒物和表面沉积物的δ 15 N的比较,在高输出生产力和/或有机物保存良好的区域(如陆架区域),海洋表层沉积物δ 15 Nbulk 和上覆海水下透光带的硝酸盐δ 15 N-NO3 - 图3 ),水体沉降过程和沉积后的各种作用对利用沉积物氮同位素示踪海洋上层水体的氮同位素影响不大[25 ] .Galbraith等[11 ] 汇聚全球现代海底样品δ 15 N数据结果表明,现代海底沉积物的δ 15 N与我们对水体中氮循环的认识预测值基本一致,反硝化作用强的海域的沉积物δ 15 N高,固氮作用显著的海域则相反(图3 ),区域1的研究站点的沉积物氮同位素高于海水平均硝酸盐氮同位素(δ 15 Nbulk >5‰),主要位于反硝化作用显著的海域,而区域2则相反,其δ 15 Nbulk <5‰,主要位于固氮作用强的海域,因此δ 15 Nbulk 基本可以反映海洋循环、生物泵和大尺度氮循环的变化特征 [50 ,51 ,52 ,53 ] . ...
Nitrogen cycling in the sediments of Santa Barbara basin and Eastern Subtropical North Pacific: Nitrogen isotopes, diagenesis and possible chemosymbiosis between two lithotrophs (Thioploca and Anammox)-“riding on a glider”
1
2006
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
Surface sediment distribution along the Chilean continental slope related to upwelling and productivity
1
2000
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
Nitrogen isotope and chlorin paleoproductivity records from the Northern South China Sea: remote vs. local forcing of millennial- and orbital-scale variability
1
2003
... 通常情况下,海洋沉积物中的生物降解作用会优先降解质量数轻的同位素,导致重同位素的富集,所以表层沉积物的δ 15 Nbulk 一般比δ 15 Nsink 高(如图3 所示,表层沉积物δ 15 N基本位于在1∶1等值线上侧,比相同站位的δ 15 N-NO3 - 图3 的1∶1等值线附近),比如在南海二者分别为5‰~6‰和2.7‰~3.6‰[29 ] ,北印度洋沉积物δ 15 N比捕获器颗粒物的高1.5‰~3.0‰[46 ] .在有机物贫乏、堆积速率低的深海沉积中这种降解效应更为显著,沉降颗粒物和表层沉积物的稳定氮同位素偏差显著增大,例如在南大洋等低有机氮通量区,海底沉积物记录的δ 15 N有可能大幅增加(3‰~6‰)[54 ] .但是由于难以直接识别缓慢的沉积物变化,用δ 15 Nbulk 阐明沉积有机氮库中的同位素转移的确切机制还存在一定不确定性.近年也有研究发现δ 15 Nbulk 的变化与水深有一定相关性,初始值在下沉和沉降的过程中遭到改变.水深可能是影响早期成岩作用造成同位素偏移的最强因素,与深度有关的海底暴露时间差异也可能是控制稳定氮同位素变化程度的另一因素[11 ] .而沉降为表层沉积物后,δ 15 Nbulk 在埋藏阶段又会受到最大的分馏影响,导致相同或近似站点悬浮颗粒和表层沉积物之间的差值有较大的范围(平均值为2.3‰±1.8‰).因此有学者认为δ 15 Nbulk 实际反映的是沉降有机质在下沉及埋藏过程中由于氮的去除或添加导致的次级同位素变化,拥有较明显的区域特征[49 ] .例如秘鲁—智利岸外27°~42°S的沉积物δ 15 Nbulk 离反硝化区的距离越远值越低[55 ] ,在相距不超过15 km的南海沉积物中40 ka以来稳定氮同位素变化很大,可能与沉积物中细颗粒物质的再分布有关[56 ] . ...
Diagenetic control of nitrogen isotope ratios in Holocene sapropels and recent sediments from the Eastern Mediterranean Sea
2
2010
... 海洋沉积物受到复杂沉积环境的影响,如早期成岩作用,陆源输入和矿物组分等都会影响沉积物稳定氮同位素组成.海洋沉积速率或沉积物—水界面条件会随时间而变化,使得沉积物中稳定氮同位素随时间而存在成岩作用的富集,导致沉积物在早期埋藏阶段同位素的升高[49 ] ,早期成岩作用主要发生在表层10 cm左右,受有机质含量、氧含量以及底栖动物扰动等的影响,一般有机质含量越高成岩作用越显著,而底栖动物的扰动对矿化作用也有一定程度的促进作用.地中海作为世界上最古老的海域,在保存沉积物有机质(如交替的泥灰质和腐泥层)方面存在着显著的差异,在低有机碳的泥灰层具有高δ 15 Nbulk 而在有机质丰富的腐泥层δ 15 Nbulk 则较低.部分研究者认为该地区的δ 15 Nbulk 的原始数据已遭到了改变[57 ] ;而另有研究者认为大体上还是反映了原始的信号特征[58 ] .前者的主要依据是降解指数与δ 15 Nbulk 的线性关系,认为成岩作用导致的同位素蚀变提高了泥灰岩有机质δ 15 N,而腐泥层中有机质δ 15 Nbulk 较低是保存作用增强的结果[57 ] .后者主要是借助于δ 15 Nbulk 和绿素δ 15 N(δ 15 Nchlorin )的一致性证明两者的差异并不是由于后期改变而是表层海水生物地化条件的差异性造成的[58 ] ,因为δ 15 Nchlorin 主要反映了海洋浮游植物群落利用的初始海水营养盐δ 15 N的组成,受成岩作用的影响极小. ...
... [57 ].后者主要是借助于δ 15 Nbulk 和绿素δ 15 N(δ 15 Nchlorin )的一致性证明两者的差异并不是由于后期改变而是表层海水生物地化条件的差异性造成的[58 ] ,因为δ 15 Nchlorin 主要反映了海洋浮游植物群落利用的初始海水营养盐δ 15 N的组成,受成岩作用的影响极小. ...
Evidence from chlorin nitrogen isotopes for alternating nutrient regimes in the Eastern Mediterranean Sea
2
2010
... 海洋沉积物受到复杂沉积环境的影响,如早期成岩作用,陆源输入和矿物组分等都会影响沉积物稳定氮同位素组成.海洋沉积速率或沉积物—水界面条件会随时间而变化,使得沉积物中稳定氮同位素随时间而存在成岩作用的富集,导致沉积物在早期埋藏阶段同位素的升高[49 ] ,早期成岩作用主要发生在表层10 cm左右,受有机质含量、氧含量以及底栖动物扰动等的影响,一般有机质含量越高成岩作用越显著,而底栖动物的扰动对矿化作用也有一定程度的促进作用.地中海作为世界上最古老的海域,在保存沉积物有机质(如交替的泥灰质和腐泥层)方面存在着显著的差异,在低有机碳的泥灰层具有高δ 15 Nbulk 而在有机质丰富的腐泥层δ 15 Nbulk 则较低.部分研究者认为该地区的δ 15 Nbulk 的原始数据已遭到了改变[57 ] ;而另有研究者认为大体上还是反映了原始的信号特征[58 ] .前者的主要依据是降解指数与δ 15 Nbulk 的线性关系,认为成岩作用导致的同位素蚀变提高了泥灰岩有机质δ 15 N,而腐泥层中有机质δ 15 Nbulk 较低是保存作用增强的结果[57 ] .后者主要是借助于δ 15 Nbulk 和绿素δ 15 N(δ 15 Nchlorin )的一致性证明两者的差异并不是由于后期改变而是表层海水生物地化条件的差异性造成的[58 ] ,因为δ 15 Nchlorin 主要反映了海洋浮游植物群落利用的初始海水营养盐δ 15 N的组成,受成岩作用的影响极小. ...
... [58 ],因为δ 15 Nchlorin 主要反映了海洋浮游植物群落利用的初始海水营养盐δ 15 N的组成,受成岩作用的影响极小. ...
Distribution and sources of organic carbon, nitrogen and their isotopic signatures in sediments from the Ayeyarwady (Irrawaddy) continental shelf, northern Andaman Sea
1
2008
... 由于海洋和陆地有机物δ 15 N的差异,陆源物质的输入也会导致海洋沉积物δ 15 N的变化.典型的海洋沉积物δ 15 N与大洋溶解硝酸盐(平均δ 15 N为5‰)相近.而陆地植物通过固氮菌吸收大气N2 进行光合作用,其δ 15 N一般较轻,往往小于5‰.因此沉积物δ 15 N从陆架区向深海区呈现增加趋势,在南海从4.2‰~5‰增加到6‰[29 ] ,在北印度洋从小于4‰增加到大于11‰[46 ] ,北安达曼海从小于4‰增加到大于5‰ [59 ] .同时近几十年来随着人类活动的增加,大气中大幅增加的含氮化合物通过干湿沉降到地表,导致湖泊和海洋富营养化.因此干湿沉降中的氮不仅有闪电、土壤释放等自然来源还有化石燃料燃烧等人类来源,前者δ 15 N一般很低(<-20‰),而后者影响因素复杂,δ 15 N有较大变化范围(-23.3‰~25.6‰)[60 ] .外源氮可能会影响和改变海洋氮库的平衡,总体上占比为12%~25%,在开放大洋中更低[61 ] .但陆源输入的氮同位素很难被界定,包括河流到海岸带过程中存在的不同形态的氮化合物以及各种反应,因此目前仍缺乏对陆源输入的氮的同位素组成进行系统研究. ...
Atmospheric reactive nitrogen cycle and stable nitrogen isotope processes:Progresses and perspectives
1
2019
... 由于海洋和陆地有机物δ 15 N的差异,陆源物质的输入也会导致海洋沉积物δ 15 N的变化.典型的海洋沉积物δ 15 N与大洋溶解硝酸盐(平均δ 15 N为5‰)相近.而陆地植物通过固氮菌吸收大气N2 进行光合作用,其δ 15 N一般较轻,往往小于5‰.因此沉积物δ 15 N从陆架区向深海区呈现增加趋势,在南海从4.2‰~5‰增加到6‰[29 ] ,在北印度洋从小于4‰增加到大于11‰[46 ] ,北安达曼海从小于4‰增加到大于5‰ [59 ] .同时近几十年来随着人类活动的增加,大气中大幅增加的含氮化合物通过干湿沉降到地表,导致湖泊和海洋富营养化.因此干湿沉降中的氮不仅有闪电、土壤释放等自然来源还有化石燃料燃烧等人类来源,前者δ 15 N一般很低(<-20‰),而后者影响因素复杂,δ 15 N有较大变化范围(-23.3‰~25.6‰)[60 ] .外源氮可能会影响和改变海洋氮库的平衡,总体上占比为12%~25%,在开放大洋中更低[61 ] .但陆源输入的氮同位素很难被界定,包括河流到海岸带过程中存在的不同形态的氮化合物以及各种反应,因此目前仍缺乏对陆源输入的氮的同位素组成进行系统研究. ...
大气氧化态活性氮循环与稳定同位素过程: 问题与展望
1
2019
... 由于海洋和陆地有机物δ 15 N的差异,陆源物质的输入也会导致海洋沉积物δ 15 N的变化.典型的海洋沉积物δ 15 N与大洋溶解硝酸盐(平均δ 15 N为5‰)相近.而陆地植物通过固氮菌吸收大气N2 进行光合作用,其δ 15 N一般较轻,往往小于5‰.因此沉积物δ 15 N从陆架区向深海区呈现增加趋势,在南海从4.2‰~5‰增加到6‰[29 ] ,在北印度洋从小于4‰增加到大于11‰[46 ] ,北安达曼海从小于4‰增加到大于5‰ [59 ] .同时近几十年来随着人类活动的增加,大气中大幅增加的含氮化合物通过干湿沉降到地表,导致湖泊和海洋富营养化.因此干湿沉降中的氮不仅有闪电、土壤释放等自然来源还有化石燃料燃烧等人类来源,前者δ 15 N一般很低(<-20‰),而后者影响因素复杂,δ 15 N有较大变化范围(-23.3‰~25.6‰)[60 ] .外源氮可能会影响和改变海洋氮库的平衡,总体上占比为12%~25%,在开放大洋中更低[61 ] .但陆源输入的氮同位素很难被界定,包括河流到海岸带过程中存在的不同形态的氮化合物以及各种反应,因此目前仍缺乏对陆源输入的氮的同位素组成进行系统研究. ...
Nitrogen cycles: Past, present, and future
1
2004
... 由于海洋和陆地有机物δ 15 N的差异,陆源物质的输入也会导致海洋沉积物δ 15 N的变化.典型的海洋沉积物δ 15 N与大洋溶解硝酸盐(平均δ 15 N为5‰)相近.而陆地植物通过固氮菌吸收大气N2 进行光合作用,其δ 15 N一般较轻,往往小于5‰.因此沉积物δ 15 N从陆架区向深海区呈现增加趋势,在南海从4.2‰~5‰增加到6‰[29 ] ,在北印度洋从小于4‰增加到大于11‰[46 ] ,北安达曼海从小于4‰增加到大于5‰ [59 ] .同时近几十年来随着人类活动的增加,大气中大幅增加的含氮化合物通过干湿沉降到地表,导致湖泊和海洋富营养化.因此干湿沉降中的氮不仅有闪电、土壤释放等自然来源还有化石燃料燃烧等人类来源,前者δ 15 N一般很低(<-20‰),而后者影响因素复杂,δ 15 N有较大变化范围(-23.3‰~25.6‰)[60 ] .外源氮可能会影响和改变海洋氮库的平衡,总体上占比为12%~25%,在开放大洋中更低[61 ] .但陆源输入的氮同位素很难被界定,包括河流到海岸带过程中存在的不同形态的氮化合物以及各种反应,因此目前仍缺乏对陆源输入的氮的同位素组成进行系统研究. ...
Early diagenesis of organic matter from sediments of the eastern subtropical Atlantic: Evidence from stable nitrogen and carbon isotopes
1
2001
... 沉积物中的无机组分对于稳定氮同位素的研究也有很大的影响,无机氮是沉积物总氮中的重要组成部分,含量从40%(亚热带大西洋沉积物中)[62 ] 到约60%不等(北冰洋、亚北极西部太平洋,中国南海沉积物中)[17 ,63 ,64 ] .高无机氮组分会导致沉积物稳定氮同位素呈现完全不同的特征,造成C/N和δ 15 Nbulk 偏移,使用KOH-KOBr处理去除有机氮间接获得有机氮数据或者直接酸处理去除无机氮的影响[65 ] .快速沉积环境中的大量沉积物基质可能含有许多不稳定碳以及与矿物相关的无机氮,特别是高矿物质但低有机物的环境,物种差异及其同位素组成对沉积物稳定氮同位素分析都有重要影响. ...
Nitrogen and carbon isotopic composition of marine and terrestrial organic matter in Arctic Ocean sediments: Implications for nutrient utilization and organic matter composition
1
2001
... 沉积物中的无机组分对于稳定氮同位素的研究也有很大的影响,无机氮是沉积物总氮中的重要组成部分,含量从40%(亚热带大西洋沉积物中)[62 ] 到约60%不等(北冰洋、亚北极西部太平洋,中国南海沉积物中)[17 ,63 ,64 ] .高无机氮组分会导致沉积物稳定氮同位素呈现完全不同的特征,造成C/N和δ 15 Nbulk 偏移,使用KOH-KOBr处理去除有机氮间接获得有机氮数据或者直接酸处理去除无机氮的影响[65 ] .快速沉积环境中的大量沉积物基质可能含有许多不稳定碳以及与矿物相关的无机氮,特别是高矿物质但低有机物的环境,物种差异及其同位素组成对沉积物稳定氮同位素分析都有重要影响. ...
Sedimentary inorganic nitrogen and its isotope ratio in the western subarctic Pacific over the last 145 kyr
1
2009
... 沉积物中的无机组分对于稳定氮同位素的研究也有很大的影响,无机氮是沉积物总氮中的重要组成部分,含量从40%(亚热带大西洋沉积物中)[62 ] 到约60%不等(北冰洋、亚北极西部太平洋,中国南海沉积物中)[17 ,63 ,64 ] .高无机氮组分会导致沉积物稳定氮同位素呈现完全不同的特征,造成C/N和δ 15 Nbulk 偏移,使用KOH-KOBr处理去除有机氮间接获得有机氮数据或者直接酸处理去除无机氮的影响[65 ] .快速沉积环境中的大量沉积物基质可能含有许多不稳定碳以及与矿物相关的无机氮,特别是高矿物质但低有机物的环境,物种差异及其同位素组成对沉积物稳定氮同位素分析都有重要影响. ...
Isotopic composition and speciation of sedimentary nitrogen and carbon in the Okinawa Trough over the past 30 ka
1
2015
... 沉积物中的无机组分对于稳定氮同位素的研究也有很大的影响,无机氮是沉积物总氮中的重要组成部分,含量从40%(亚热带大西洋沉积物中)[62 ] 到约60%不等(北冰洋、亚北极西部太平洋,中国南海沉积物中)[17 ,63 ,64 ] .高无机氮组分会导致沉积物稳定氮同位素呈现完全不同的特征,造成C/N和δ 15 Nbulk 偏移,使用KOH-KOBr处理去除有机氮间接获得有机氮数据或者直接酸处理去除无机氮的影响[65 ] .快速沉积环境中的大量沉积物基质可能含有许多不稳定碳以及与矿物相关的无机氮,特别是高矿物质但低有机物的环境,物种差异及其同位素组成对沉积物稳定氮同位素分析都有重要影响. ...
Reduced nitrogen fixation in the glacial ocean inferred from changes in marine nitrogen and phosphorus inventories
1
2002
... 沉积物中有机物稳定氮同位素变化同样是了解地质历史上海洋氮循环变化的有效手段,虽然其作为海洋氮动力学记录的保真度仍有疑问,但通过结合其他指标对不同沉积状态记录进行对比可以更好地约束环境参数,在重建古海洋古气候方面取得了显著的进展.事实上,迄今为止许多δ 15 N记录已显示了不同空间和时间尺度上的相互关联,证明δ 15 N记录与全球气候变化的紧密联系[13 ,66 ,67 ] . ...
The effect of millennial-scale changes in Arabian Sea denitrification on atmospheric CO2
2
2002
... 沉积物中有机物稳定氮同位素变化同样是了解地质历史上海洋氮循环变化的有效手段,虽然其作为海洋氮动力学记录的保真度仍有疑问,但通过结合其他指标对不同沉积状态记录进行对比可以更好地约束环境参数,在重建古海洋古气候方面取得了显著的进展.事实上,迄今为止许多δ 15 N记录已显示了不同空间和时间尺度上的相互关联,证明δ 15 N记录与全球气候变化的紧密联系[13 ,66 ,67 ] . ...
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Glacial-interglacial variability in denitrification in the world's oceans: Causes and consequences
1
2000
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Detailed sedimentary N isotope records from Cariaco Basin for Terminations I and V: Local and global implications
8
2007
... 据参考文献[69 ]修改:智利海盆(黑)[70 ] ,阿拉伯海(蓝)[71 ] ;卡里亚科海盆(红)[69 ] ...
... [69 ] ...
... Modified after reference [69 ]: Chile basin (Black)[70 ] , Arabian Sea (Blue)[71 ] ; Cariaco Basin (Red)[69 ] ...
... [69 ] ...
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
... [69 ](图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
... [69 ,79 ].Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物
δ 15 N
bulk 几乎没有变化(<0.5%),如北大西洋
[84 ] 、南印度洋
[85 ] 、苏禄海
[30 ] 和南中国海
[86 ,87 ] (
图5 ).现代开放大洋中同样缺乏间冰期
δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast
[86 ] 将南海不同区域
δ 15 N
bulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低
[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素
[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的
δ 15 N-NO
3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(
δ 15 N正偏)和氮固定作用(
δ 15 N负偏)导致的同位素平衡相关
[86 ,92 ] .Eugster等
[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素.
图5 不同类型海域<strong>50 ka</strong>沉积物稳定氮同位素记录对比(据参考文献[<xref ref-type="bibr" rid="R69">69</xref>,<xref ref-type="bibr" rid="R88">88</xref>]修改) 卡里亚科海盆(蓝色)沉积物δ 15 Nbulk [79 ] ;阿拉伯海(棕红)沉积物δ 15 Nbulk [19 ] ;南海沉积物δ 15 Nbulk (黑色)[86 ] 和FB-δ 15 N(绿色)[89 ] ;阴影部分为间冰期,数字为海洋同位素阶段 ...
Melting of the Patagonian Ice Sheet and deglacial perturbations of the nitrogen cycle in the eastern South Pacific
3
2006
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
... 据参考文献[69 ]修改:智利海盆(黑)[70 ] ,阿拉伯海(蓝)[71 ] ;卡里亚科海盆(红)[69 ] ...
... Modified after reference [69 ]: Chile basin (Black)[70 ] , Arabian Sea (Blue)[71 ] ; Cariaco Basin (Red)[69 ] ...
Variations in tropical convection as an amplifier of global climate change at the millennial scale
3
2005
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
... 据参考文献[69 ]修改:智利海盆(黑)[70 ] ,阿拉伯海(蓝)[71 ] ;卡里亚科海盆(红)[69 ] ...
... Modified after reference [69 ]: Chile basin (Black)[70 ] , Arabian Sea (Blue)[71 ] ; Cariaco Basin (Red)[69 ] ...
Nitrogen isotope variations in Santa Barbara Basin sediments: Implications for denitrification in the eastern tropical North Pacific during the last 50,000 years
1
2000
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Intermittent existence of a southern Californian upwelling cell during submillennial climate change of the last 60 kyr
1
2004
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Comment on sedimentary phosphorus record from the Oman margin: New evidence of high productivity during glacial periods by F. Tamburini
1
2004
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Southern Ocean control on the extent of denitrification in the southeast Pacific over the last 70 ka
2
2007
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
... [75 ].此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Increase in water column denitrification during the deglaciation controlled by oxygen demand in the eastern equatorial Pacific
1
2010
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Oxygen minimum zone expansion in the eastern tropical North Pacific during deglaciation
1
2006
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Consistent relationship between global climate and surface nitrate utilization in the western subarctic Pacific throughout the last 500 ka
1
2008
... 利用沉积物稳定氮同位素重建历史时期的海洋硝酸盐浓度变化最初来自对阿拉伯海和东赤道太平洋的研究[68 ] ,这些典型的反硝化作用强的海区拥有明显的OMZ,强烈的水柱反硝化作用造成显著的同位素分馏,简单使用沉积物δ 15 Nbulk 即可反演海水温跃层反硝化作用程度和/或活动变化,因此为全球大洋氮循环的地质学研究提供了丰富的信息.阿拉伯海、东赤道太平洋高分辨率的δ 15 N记录不仅显示出良好的一致性而且显著响应北大西洋气候快速变化事件,证明了高δ 15 N时期温跃层低氧区的扩大[32 ] ,具体表现为冰期δ 15 N降低,水柱反硝化作用减弱;间冰期则相反(图4 ),其中阿拉伯海的反硝化作用记录具有明显的千年尺度循环,DO(Dansgaard-Oeschger)旋回中δ 15 N变化幅度达3‰~4‰[67 ] ,新仙女木(Younger Dryas,YD)事件时期快速变化幅度达2‰,该区域反硝化作用的高低极可能与南亚季风的强弱变化有关[71 ] .南加州岸外上升流区和东北热带太平洋的圣芭芭拉(Santa Barbara)海盆也有类似千年尺度变化记录[72 ,73 ] ,只是对YD事件和DO旋回的响应程度相对较弱.在冰期循环模式类似的模式下,在研究中也发现与上述冰消期(15~10 ka)温跃层反硝化加强过程不一致的区域,如智利、秘鲁和南墨西哥岸外的研究显示19 ka时δ 15 N突然升高,幅度达4.5‰,说明反硝化的突然增强,随后冰消期阶段呈现逐渐下降趋势[70 ,74 ] (图4 ),而研究显示南半球反硝化主要与亚南极模态水和南极气候事件密切相关,而不是之前认为的当地初级生产力和南大洋通风变化[32 ,75 ,76 ,77 ] .这些研究显示了南北半球稳定氮同位素演变存在非同步性,南半球的反硝化作用记录超前于北半球[75 ] .此外,亚北极太平洋3个深水站位500 ka以来的δ 15 N呈现不同的记录,该区域东部末次冰期以来的沉积物氮同位素组成与北美西部边缘的δ 15 N分布具有一致性,说明东北太平洋δ 15 N-NO3 - [78 ] .以上研究说明海洋环境的复杂、多变造成的区域差异性特征可能会导致截然不同的地质学记录,因此,在研究局部氮循环时需要特别关注当地的水文特征. ...
Glacial/interglacial variations in production and nitrogen fixation in the Cariaco Basin during the last 580 kyr
3
1998
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
... 卡里亚科海盆(蓝色)沉积物δ 15 Nbulk [79 ] ;阿拉伯海(棕红)沉积物δ 15 Nbulk [19 ] ;南海沉积物δ 15 Nbulk (黑色)[86 ] 和FB-δ 15 N(绿色)[89 ] ;阴影部分为间冰期,数字为海洋同位素阶段 ...
... Cariaco basin (blue) sediments δ 15 Nbulk [79 ] ; Arabian sea (brown red) sediments δ 15 Nbulk [19 ] The South China Sea sediments δ 15 Nbulk (black)[86 ] and FB-δ 15 N (green)[89 ] . The shaded part is the interglacial period, and the figure is the Marine Isotope Stage(MIS) ...
Deglacial nitrogen isotope changes in the Gulf of Mexico: Evidence from bulk sedimentary and foraminifera-bound nitrogen in Orca Basin sediments
2
2011
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
... 为了更好地研究固氮海域的古海洋学特征,科学家使用浮游有孔虫FB-δ 15 N等指标协助反演氮同位素演变特征.Ren等[88 ] 的最新研究发现南海860 ka有孔虫FB-δ 15 N记录与海平面升降同步变化,与底栖反硝化作用的强弱密切相关(图5 ),表明海平面变化是冰期间冰期氮固定作用变化的主要驱动因素,同时还驱动着大陆边缘底栖反硝化作用的冰期循环.冰期海平面大幅度下降,海洋环境缩小后沉积物向上层海水释放的营养物质相应降低,此时沉积物反硝化作用对去除海洋中生物可利用氮的贡献可达一半或更多,因此南海可能倾向于底栖脱氮作用和氮固定作用之间的耦合.墨西哥湾早前的研究结果与南海相似,利用FB-δ 15 N结合所观察到的C/N值的关系校正后,沉积物δ 15 Nbulk 从冰期到全新世δ 15 N大幅减少(接近2.5倍),氮固定作用大幅增加[80 ] .综上来看,沉积物δ 15 Nbulk 记录在固氮作用为主的海域出现误差的可能性较大.同时复杂的外源输入也增加了δ 15 Nbulk 解释的难度.在更多时候由于海源和陆源物质的混合,反硝化作用和固氮作用的相互作用,沉积物δ 15 Nbulk 在冰期间冰期循环中将缺乏显著的变化.因此在未来的研究中需要注意多指标的结合利用,才能更准确地解释海洋氮固定作用变化的多重因素. ...
Nutrient conditions in the subpolar North Atlantic during the last glacial period reconstructed from foraminifera-bound nitrogen isotopes
1
2013
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
Biological productivity during sapropel S5 formation in the Eastern Mediterranean Sea: Evidence from stable isotopes of nitrogen and carbon
1
2001
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
N2 -fixing cyanobacteria supplied nutrient N for Cretaceous oceanic anoxic events
1
2004
... 海洋中被固定的氮为全球浮游植物生产力提供了能源,海洋固定氮库的变化被认为是大气二氧化碳浓度冰期旋回变化的原因之一[69 ] .对于缺乏典型的发生反硝化作用而固氮作用强盛的海区而言,地质历史时期稳定氮同位素的变化主要认为是氮固定作用的变化.海洋固氮受多种因素的影响,地质记录中氮固定作用的强弱可以反演历史海水温度、铁的输入以及磷的有效性等.关于冰期间冰期固氮作用变化最典型的记录来自于大西洋卡里亚科海盆,间冰期稳定氮同位素显著低于冰期,沉积物δ 15 N从LGM到全新世降幅约达3‰[69 ] (图4 ),固氮作用显著增强.6~3 ka时固氮量的增强与非洲向热带北大西洋和加勒比海地区的沙尘输送的铁的增加相吻合,但也有可能与当地以及全球反硝化变化有关[69 ,79 ] .Meckler等[80 ] 也认为末次冰期期间风尘输入的Fe的增多并不是造成固氮作用增强主要因素,冰期至冰消期δ 15 N的降低是大西洋氮固定对于冰消期全球大洋反硝化作用降低的响应.利用加勒比海沉积物中浮游生物稳定氮同位素对过去160 ka以来北大西洋固氮作用变化的重建结果存在23 ka的周期变化,无法用温度的升降、含铁粉尘的供给或水柱反硝化变化来解释,主要是对轨道驱动的赤道大西洋上升流变化将过剩磷输入热带北大西洋表面的响应.同时固氮量在MIS4和MIS6两个冰期期间减少,这个时期北大西洋深水变浅成为北大西洋中层水,将现代从南大洋进入的富含磷的中深层水隔离[81 ] .东地中海也是一个对固氮作用敏感的地区,夏季固氮生物对生物输出量的贡献可达30%甚至更多,在暖期低N∶P大西洋水的注入促进了该地区的固氮作用,导致沉积物δ 15 N降低,富含有机物的腐泥层中δ 15 N减少3‰,固氮作用显著增强[82 ] .白垩纪沉积物δ 15 N的低值也主要与固氮作用相关,该时期固氮蓝藻的勃发为白垩纪海洋缺氧事件提供了生物生长所必需的氮营养[83 ] .苏禄海83 ka的沉积物记录中关于冰期固氮作用增强的原因也倾向于区域洋流变化特征的解释,固氮作用在MIS3和MIS2期显著增强,利用质量平衡模型得出这两个阶段该地区固氮作用显著增加是季风增强驱动海水混合引起的表层水磷酸盐累积所致[30 ] . ...
Sources of fine-sized organic matter in North Atlantic Heinrich Layers: δ13 C and δ15 N tracers
1
2002
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
Contribution of Southern Ocean surface-water stratification to low atmospheric CO2 concentrations during the last glacial period
1
1997
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
Unchanged nitrogen isotopic composition of organic matter in the South China Sea during the last climatic cycle: Global implications
5
2000
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
... [86 ]将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
... [86 ,92 ].Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
... 卡里亚科海盆(蓝色)沉积物δ 15 Nbulk [79 ] ;阿拉伯海(棕红)沉积物δ 15 Nbulk [19 ] ;南海沉积物δ 15 Nbulk (黑色)[86 ] 和FB-δ 15 N(绿色)[89 ] ;阴影部分为间冰期,数字为海洋同位素阶段 ...
... Cariaco basin (blue) sediments δ 15 Nbulk [79 ] ; Arabian sea (brown red) sediments δ 15 Nbulk [19 ] The South China Sea sediments δ 15 Nbulk (black)[86 ] and FB-δ 15 N (green)[89 ] . The shaded part is the interglacial period, and the figure is the Marine Isotope Stage(MIS) ...
Investigating the history of East Asian monsoon and climate during the last glacial-interglacial period (0-140,000 years): Mineralogy and geochemistry of ODP Sites 1143 and 1144, South China Sea
1
2003
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
Impact of glacial/interglacial sea level change on the ocean nitrogen cycle
2
2017
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物
δ 15 N
bulk 几乎没有变化(<0.5%),如北大西洋
[84 ] 、南印度洋
[85 ] 、苏禄海
[30 ] 和南中国海
[86 ,87 ] (
图5 ).现代开放大洋中同样缺乏间冰期
δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast
[86 ] 将南海不同区域
δ 15 N
bulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低
[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素
[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的
δ 15 N-NO
3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(
δ 15 N正偏)和氮固定作用(
δ 15 N负偏)导致的同位素平衡相关
[86 ,92 ] .Eugster等
[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素.
图5 不同类型海域<strong>50 ka</strong>沉积物稳定氮同位素记录对比(据参考文献[<xref ref-type="bibr" rid="R69">69</xref>,<xref ref-type="bibr" rid="R88">88</xref>]修改) 卡里亚科海盆(蓝色)沉积物δ 15 Nbulk [79 ] ;阿拉伯海(棕红)沉积物δ 15 Nbulk [19 ] ;南海沉积物δ 15 Nbulk (黑色)[86 ] 和FB-δ 15 N(绿色)[89 ] ;阴影部分为间冰期,数字为海洋同位素阶段 ...
... 为了更好地研究固氮海域的古海洋学特征,科学家使用浮游有孔虫FB-δ 15 N等指标协助反演氮同位素演变特征.Ren等[88 ] 的最新研究发现南海860 ka有孔虫FB-δ 15 N记录与海平面升降同步变化,与底栖反硝化作用的强弱密切相关(图5 ),表明海平面变化是冰期间冰期氮固定作用变化的主要驱动因素,同时还驱动着大陆边缘底栖反硝化作用的冰期循环.冰期海平面大幅度下降,海洋环境缩小后沉积物向上层海水释放的营养物质相应降低,此时沉积物反硝化作用对去除海洋中生物可利用氮的贡献可达一半或更多,因此南海可能倾向于底栖脱氮作用和氮固定作用之间的耦合.墨西哥湾早前的研究结果与南海相似,利用FB-δ 15 N结合所观察到的C/N值的关系校正后,沉积物δ 15 Nbulk 从冰期到全新世δ 15 N大幅减少(接近2.5倍),氮固定作用大幅增加[80 ] .综上来看,沉积物δ 15 Nbulk 记录在固氮作用为主的海域出现误差的可能性较大.同时复杂的外源输入也增加了δ 15 Nbulk 解释的难度.在更多时候由于海源和陆源物质的混合,反硝化作用和固氮作用的相互作用,沉积物δ 15 Nbulk 在冰期间冰期循环中将缺乏显著的变化.因此在未来的研究中需要注意多指标的结合利用,才能更准确地解释海洋氮固定作用变化的多重因素. ...
Data-based estimates of suboxia, denitrification, and N2 O production in the ocean and their sensitivities to dissolved O2
3
2012
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
... 卡里亚科海盆(蓝色)沉积物δ 15 Nbulk [79 ] ;阿拉伯海(棕红)沉积物δ 15 Nbulk [19 ] ;南海沉积物δ 15 Nbulk (黑色)[86 ] 和FB-δ 15 N(绿色)[89 ] ;阴影部分为间冰期,数字为海洋同位素阶段 ...
... Cariaco basin (blue) sediments δ 15 Nbulk [79 ] ; Arabian sea (brown red) sediments δ 15 Nbulk [19 ] The South China Sea sediments δ 15 Nbulk (black)[86 ] and FB-δ 15 N (green)[89 ] . The shaded part is the interglacial period, and the figure is the Marine Isotope Stage(MIS) ...
On the sedimentological origin of down-core variations of bulk sedimentary nitrogen isotope ratios
1
2005
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
Rapid flooding of the Sunda Shelf: A late glacial sea level record
1
2000
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...
Nitrogen in past marine environments
1
2008
... 与上述地区的岩芯记录显示出明显的冰期—间冰期稳定氮同位素的变化相比,有些地区的沉积物δ 15 Nbulk 几乎没有变化(<0.5%),如北大西洋[84 ] 、南印度洋[85 ] 、苏禄海[30 ] 和南中国海[86 ,87 ] (图5 ).现代开放大洋中同样缺乏间冰期δ 15 N低于冰期的记录,可能是因为固氮作用引起的氮同位素分馏程度小,被反硝化作用的巨大分馏信号压制,导致沉积物缺乏明显的固氮信号.Kienast[86 ] 将南海不同区域δ 15 Nbulk 较小的变化解释为底栖反硝化作用影响.底栖反硝化作用一般发生在大陆架近岸沉积物中,因为大陆架水深较浅,无论是营养的释放或是上层有机质的沉降都比较快速,因此沉积物中的有机物质通量高,细菌分解作用导致的氧含量低共同促进了底栖反硝化作用的发生.冰期时海平面降低,海洋面积的缩小会降低大陆架浅层循环的速率,底栖反硝化作用也随着大陆架区域范围的缩小而有所降低[89 ,90 ] .对于底栖反硝化作用来说,底物的变化和海平面的升降是很重要的驱动因素[91 ] .底栖反硝化作用在很大程度上与LGM和全新世之间的水柱反硝化作用一致,因为只有这两个反硝化过程保持相对固定的比率才能确保在远离强固氮作用和水柱反硝化作用的区域中有相对恒定的δ 15 N-NO3 - . 这些海域的沉积记录中稳定氮同位素缺乏明显变化可能也与区域的反硝化作用(δ 15 N正偏)和氮固定作用(δ 15 N负偏)导致的同位素平衡相关[86 ,92 ] .Eugster等[10 ] 通过模型模拟冰消期氮循环发现氮固定作用与反硝化作用的不平衡导致从冰期到间冰期海洋氮储库下降了16%~50%.氮库减少在很大程度上是由于在冰期初始期固氮率的暂时大幅度下降造成的,虽然模型结果支持铁在控制固氮方面的重要性,但也驳斥了之前认为LGM过程中高固氮率的假设.与冰川变化速率相比,整个过渡时期水体反硝化速率提高了10%~100%,而全球底栖生物反硝化速率变化为-1%~58%.与以往认为的LGM-冰消期初期海洋氮库的变化是由水柱反硝化的变化驱动的观点不同,固氮作用可能是这些区域海洋氮收支变化首要的也是最重要的影响因素. ...




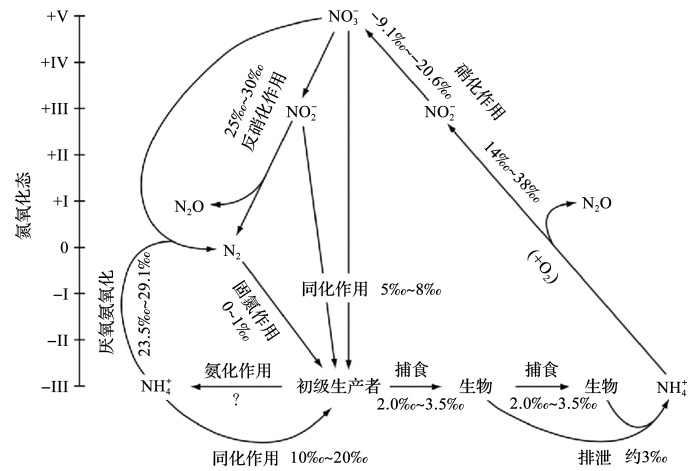
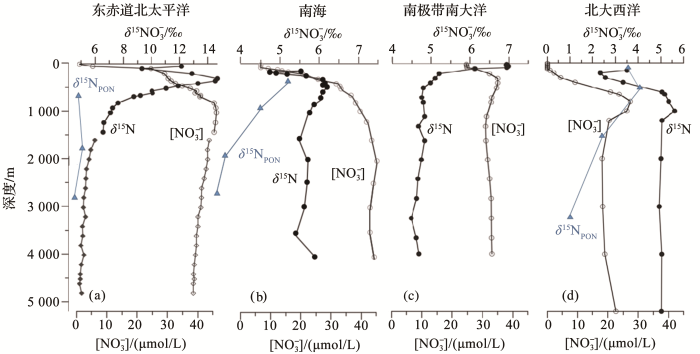
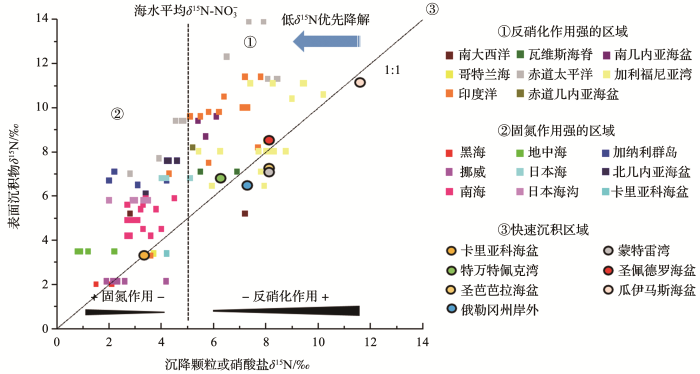
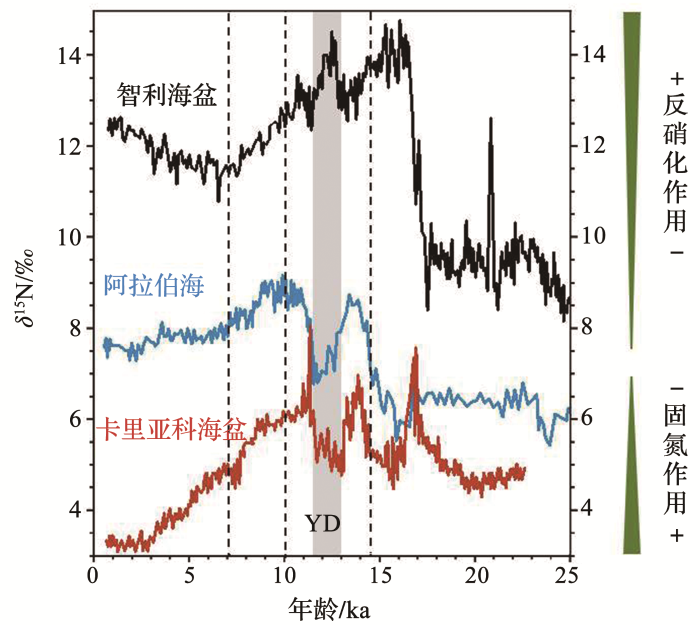
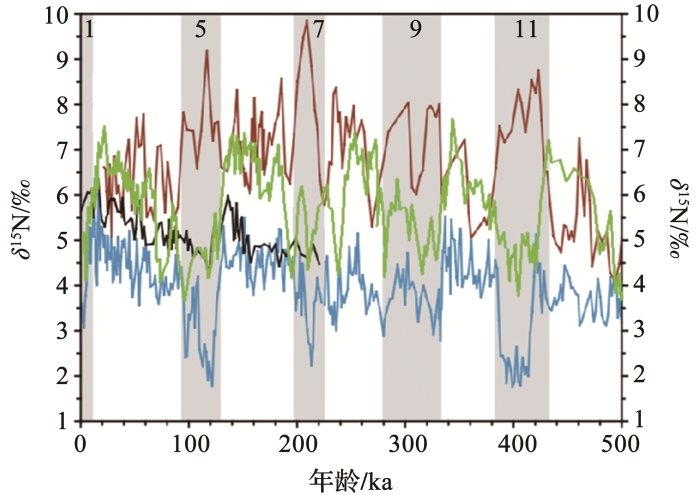

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}