Determination of rare earth elements after pre-concentration using NOBIAS-chelate PA-1?resin: Method development and application in the San Francisco Bay plume
1
2014
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
Automated preconcentration of Fe,Zn,Cu,Ni,Cd,Pb,Co,and Mn in seawater with analysis using high-resolution sector field inductively-coupled plasma mass spectrometry
2017
Low blank preconcentration technique for the determination of lead,Copper,and Cadmium in small-volume seawater samples by Isotope Dilution ICPMS
1
1997
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
The biogeochemical cycles of trace metals in the oceans
2
2003
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
The distribution of dissolved copper in the Pacific
5
1977
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... 海水中铜的稳定同位素作为一个新的指标,可以进一步推断海洋中铜的源汇格局.通过比较现代海洋输入和输出的铜通量及其平均铜同位素值,发现了海水中典型铜汇的同位素值(约+0.3‰)相较于铜源的同位素值(约+0.6‰)偏轻.假设海洋铜循环处于稳态,通过同位素质量守恒计算出来的海洋体系中铜的收支是不平衡的[23 ] ,这意味着当代海洋存在至今未被确认的轻铜同位素的源[5 ,23 ,98 ] ,其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
Copper in surface waters south of New Zealand
2
1975
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
Trace element calibrations and profiles at the Geosecs Test Station in the northeast Pacific Ocean
1
1970
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
5
2003
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... [8 ].营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... [8 ].铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... 如图5 所示,铜的垂直分布模式介于营养型元素(表层因生物利用耗竭、于深层再生)和清除型元素(有强表层水来源但在整个水柱中清除)之间,其分布受到生物利用、清除和再生等过程的共同影响[109 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] . ...
GEOTRACES—An international study of the global marine biogeochemical cycles of trace elements and their isotopes
1
2007
... 半个世纪来,痕量金属一直是海洋生物地球化学研究的重点.痕量金属(铁、钴、镍、铜、锌、锰、镉、铅和稀土元素等)在海水中的浓度极低,通常在pmol/L~nmol/L范围内[1 ~3 ] .这些金属普遍具有生物学功能,是海洋生物酶系统的辅因子以及蛋白质的组成元素,适宜浓度的痕量金属能促进浮游植物的生长,提高海洋初级生产力,进而影响碳循环和氮循环过程[4 ] .20世纪70年代,随着地球化学海洋断面研究计划(Geochemical Ocean Sections Program,GEOSECS)的实行,痕量金属在海洋中的浓度、分布及化学行为得到了初步认识[5 ~7 ] .之后,伴随着仪器检测技术的发展,以及在海水采集、前处理和分析的全程中重视使用清洁技术,痕量金属的研究得到了进一步的拓展[8 ] ,痕量金属及其同位素的海洋生物地球化学国际合作研究(An International Study of the Marine Biogeochemical Cycles of Trace Elements and Their Isotopes,GEOTRACES)项目也在21世纪初应运而生.GEOTRACES项目是继20世纪70年代GEOSECS和90年代世界大洋环流实验(World Ocean Circulation Experiment,WOCE)等大型海洋调查计划之后,在海洋研究科学委员会(Scientific Committee for Oceanic Research,SCOR)框架下组织开展的最大型的国际合作计划.GEOTRACES旨在调查世界关键海域的痕量金属及其同位素以揭示物理输运、生物地球化学循环等重要的海洋学过程.该项目规范统一了海洋痕量金属的采样、前处理和分析测定流程;制作了标准参考物质以便不同实验室之间进行方法互校;并将航次数据全网共享.海水中痕量金属的研究也由此得到了飞跃式发展[9 ] . ...
Reduction of marine phytoplankton reproduction rates by copper and cadmium
5
1986
... 海水中的铜(Copper,Cu)因其生物可用性和毒性的双重性质[10 ] ,成为海洋生物地球化学研究的关键元素之一.铜是生物酶系统的重要金属辅基,参与了海洋生物的许多生理过程,可作为微量营养元素在真光层被初级生产者循环利用[11 ~16 ] .由于特殊金属结构性质,铜在海水中易与有机或无机配体发生络合反应,从而降低其毒性和清除速率[17 ] .过渡变价金属铜在海水中也具有氧化还原的特性,这使得铜可以指示缺氧海底沉积物中黑色页岩的氧化还原转化和有机物再矿化情况[17 ,18 ] ,铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
... 铜对海洋生物具有双重作用,铜是生命必需元素,但超出需求浓度范围的铜又具有毒性[10 ] .铜作为氧化还原过渡金属,其离子态能与蛋白质形成稳定的化合物,通过氧化还原反应转移电子[25 ] ,这些性质使得铜具有丰富的生物学作用,参与到海洋生物的生理过程,最终影响海洋生态系统.图1 归纳总结了海洋中铜参与的生物地球化学过程及其海洋生理生态功能. ...
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
... [10 ],有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Feedback Interactions between Trace Metal Nutrients and Phytoplankton in the Ocean
2
2012
... 海水中的铜(Copper,Cu)因其生物可用性和毒性的双重性质[10 ] ,成为海洋生物地球化学研究的关键元素之一.铜是生物酶系统的重要金属辅基,参与了海洋生物的许多生理过程,可作为微量营养元素在真光层被初级生产者循环利用[11 ~16 ] .由于特殊金属结构性质,铜在海水中易与有机或无机配体发生络合反应,从而降低其毒性和清除速率[17 ] .过渡变价金属铜在海水中也具有氧化还原的特性,这使得铜可以指示缺氧海底沉积物中黑色页岩的氧化还原转化和有机物再矿化情况[17 ,18 ] ,铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
The role of trace metals in photosynthetic electron transport in O2 -evolving organisms
1
1999
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
Copper-containing plastocyanin used for electron transport by an oceanic diatom
3
2006
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
... ,13 ]. ...
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Superoxide dismutase and stress tolerance
1
1992
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
Nitrosopumilus maritimus genome reveals unique mechanisms for nitrification and autotrophy in globally distributed marine crenarchaea
1
2010
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
The importance of siderophores in iron nutrition of heterotrophic marine bacteria
2
1999
... 海水中的铜(Copper,Cu)因其生物可用性和毒性的双重性质[10 ] ,成为海洋生物地球化学研究的关键元素之一.铜是生物酶系统的重要金属辅基,参与了海洋生物的许多生理过程,可作为微量营养元素在真光层被初级生产者循环利用[11 ~16 ] .由于特殊金属结构性质,铜在海水中易与有机或无机配体发生络合反应,从而降低其毒性和清除速率[17 ] .过渡变价金属铜在海水中也具有氧化还原的特性,这使得铜可以指示缺氧海底沉积物中黑色页岩的氧化还原转化和有机物再矿化情况[17 ,18 ] ,铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
The complexation of 'dissolved' Cu,Zn,Cd and Pb by soluble and colloidal organic matter in Narragansett Bay,RI
3
1998
... 海水中的铜(Copper,Cu)因其生物可用性和毒性的双重性质[10 ] ,成为海洋生物地球化学研究的关键元素之一.铜是生物酶系统的重要金属辅基,参与了海洋生物的许多生理过程,可作为微量营养元素在真光层被初级生产者循环利用[11 ~16 ] .由于特殊金属结构性质,铜在海水中易与有机或无机配体发生络合反应,从而降低其毒性和清除速率[17 ] .过渡变价金属铜在海水中也具有氧化还原的特性,这使得铜可以指示缺氧海底沉积物中黑色页岩的氧化还原转化和有机物再矿化情况[17 ,18 ] ,铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
... [17 ,18 ],铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
The behavior of trace elements in seawater,sedimentary pore water,and their incorporation into carbonate minerals: A review
1
2019
... 海水中的铜(Copper,Cu)因其生物可用性和毒性的双重性质[10 ] ,成为海洋生物地球化学研究的关键元素之一.铜是生物酶系统的重要金属辅基,参与了海洋生物的许多生理过程,可作为微量营养元素在真光层被初级生产者循环利用[11 ~16 ] .由于特殊金属结构性质,铜在海水中易与有机或无机配体发生络合反应,从而降低其毒性和清除速率[17 ] .过渡变价金属铜在海水中也具有氧化还原的特性,这使得铜可以指示缺氧海底沉积物中黑色页岩的氧化还原转化和有机物再矿化情况[17 ,18 ] ,铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
Cu isotopes in marine black shales record the Great Oxidation Event
1
2016
... 海水中的铜(Copper,Cu)因其生物可用性和毒性的双重性质[10 ] ,成为海洋生物地球化学研究的关键元素之一.铜是生物酶系统的重要金属辅基,参与了海洋生物的许多生理过程,可作为微量营养元素在真光层被初级生产者循环利用[11 ~16 ] .由于特殊金属结构性质,铜在海水中易与有机或无机配体发生络合反应,从而降低其毒性和清除速率[17 ] .过渡变价金属铜在海水中也具有氧化还原的特性,这使得铜可以指示缺氧海底沉积物中黑色页岩的氧化还原转化和有机物再矿化情况[17 ,18 ] ,铜同位素在氧化还原过程中的分馏也可以对大氧化事件等古海洋学过程进行指示[19 ] .因此,铜不仅作用于初级生产力从而对全球碳、氮循环产生影响,也能指示古海洋学过程,使得海水中的铜具有重要的研究意义. ...
Oceanographic distributions of cadmium,zinc,nickel,and copper in the North Pacific
1
1980
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
Dissolved Copper (dCu) biogeochemical cycling in the subarctic Northeast Pacific and a call for improving methodologies
2
2017
... 应用GEOTRACES IDP2017[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如图4 所示.如表1 和图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放[103 ] 和洋流输运[21 ,104 ] 等因素有关. ...
... 如
图5 所示,铜的垂直分布模式介于营养型元素(表层因生物利用耗竭、于深层再生)和清除型元素(有强表层水来源但在整个水柱中清除)之间,其分布受到生物利用、清除和再生等过程的共同影响
[109 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值
[8 ] .
图5 世界不同大洋的溶解态铜垂直剖面分布(数据来自参考文献[<xref ref-type="bibr" rid="R21">21</xref>,<xref ref-type="bibr" rid="R99">99</xref>,<xref ref-type="bibr" rid="R118">118</xref>]) <strong>The vertical profiles of dCu in different ocean </strong>(<strong>data from references </strong>[<strong>21</strong>,<strong>99</strong>,<strong>118</strong>]) Fig.5 ![]()
Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
The distribution of dissolved copper in the tropical-subtropical north Atlantic across the GEOTRACES GA03 transect
2
2015
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... 铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
The oceanic mass balance of copper and zinc isotopes,investigated by analysis of their inputs,and outputs to ferromanganese oxide sediments
15
2014
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... ,23 ]. ...
... 图3 总结了大洋铜的主要源汇过程及其通量和同位素组成特征.河流是大洋铜的主要来源,每年贡献0.60~6.70 Gmol的铜[23 ,24 ] ;大气沉降次之,每年贡献0.054~0.270 Gmol的铜[23 ,24 ] ;海底沉积物再矿化以每年约0.37 Gmol的通量补充海洋铜库[24 ] ;热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出[23 ,24 ] . ...
... [23 ,24 ];海底沉积物再矿化以每年约0.37 Gmol的通量补充海洋铜库[24 ] ;热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出[23 ,24 ] . ...
... [23 ,24 ]. ...
... 值(数据来自参考文献[
23 ,
24 ,
80 ~
87 ])
<strong>Schematic diagrams illustrating the global ocean isotopic mass balance of Cu </strong>(<strong>Data from references </strong>[<strong>23</strong>,<strong>24</strong>,<strong>80~87</strong>]) Fig.3 ![]()
铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
... 铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... [23 ]的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... 已有的知识通常认为海洋是铜的汇.在氧化水体中,铜会吸附在颗粒物表面或与之沉淀,从而沉降到海底,Little等[23 ] 估计沉降到氧化沉积物中的铜通量为0.49 Gmol/a.在还原海水中,铜会与硫化物发生沉淀作用,从水体中去除,例如在总硫化物含量达到100 μmol/L的黑海深处,铜和锌几乎从海水中分离,相应地在海底沉积物中富集[94 ] .然而,沉积物—海界面铜的生物地球化学过程复杂,源汇格局仍需要进一步研究[95 ] .例如:在低氧沉积物中,微生物介导的铁、锰氧化物的还原性溶解能释放与氧结合的铜[96 ] ,成为海水中铜的来源.在无氧沉积物中,细菌还原硫酸盐产生的硫化物会与铁形成铁硫化物,再进一步与铜共沉淀[97 ] . ...
... 海水中铜的稳定同位素作为一个新的指标,可以进一步推断海洋中铜的源汇格局.通过比较现代海洋输入和输出的铜通量及其平均铜同位素值,发现了海水中典型铜汇的同位素值(约+0.3‰)相较于铜源的同位素值(约+0.6‰)偏轻.假设海洋铜循环处于稳态,通过同位素质量守恒计算出来的海洋体系中铜的收支是不平衡的[23 ] ,这意味着当代海洋存在至今未被确认的轻铜同位素的源[5 ,23 ,98 ] ,其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
... ,23 ,98 ],其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
... [23 ]和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
... 应用GEOTRACES IDP2017
[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如
图4 所示.如
表1 和
图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放
[103 ] 和洋流输运
[21 ,104 ] 等因素有关.
图4 溶解态铜在世界大洋表层的现有数据分布(<strong>0~50 m</strong>)(数据来自参考文献[<xref ref-type="bibr" rid="R23">23</xref>, <xref ref-type="bibr" rid="R83">83</xref>,<xref ref-type="bibr" rid="R84">84</xref>, <xref ref-type="bibr" rid="R99">99</xref>~<xref ref-type="bibr" rid="R102">102</xref>]) The existing data of dissolved copper in the world surface ocean (0~50 m) (data from references [23, 83,84, 99~102]) Fig.4 ![]()
表1 不同海区、不同深度的海水溶解态铜浓度 ...
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
Insights into the major processes driving the global distribution of copper in the ocean from a global model
12
2019
... 海水中溶解态铜(dissolved Copper,dCu)的浓度普遍在0.5~5 nmol/L范围内,在远离其来源的开阔大洋表层海水,dCu浓度通常小于1 nmol/L[8 ,20 ~22 ] .海洋中铜的来源主要有河流输入、大气沉降、陆源沉积物和热液活动,其输出通常以颗粒沉降的形式从水体中去除[23 ,24 ] .铜在海洋中的垂直分布受到生物利用、清除和再生等过程的影响,呈现营养—清除型混合分布的特征[8 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.已有的研究估计铜在海洋表层的停留时间为2~50年[6 ] ,在深层水中的停留时间为5 000~5 400年[5 ,23 ] . ...
... 图3 总结了大洋铜的主要源汇过程及其通量和同位素组成特征.河流是大洋铜的主要来源,每年贡献0.60~6.70 Gmol的铜[23 ,24 ] ;大气沉降次之,每年贡献0.054~0.270 Gmol的铜[23 ,24 ] ;海底沉积物再矿化以每年约0.37 Gmol的通量补充海洋铜库[24 ] ;热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出[23 ,24 ] . ...
... ,24 ];海底沉积物再矿化以每年约0.37 Gmol的通量补充海洋铜库[24 ] ;热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出[23 ,24 ] . ...
... [24 ];热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出[23 ,24 ] . ...
... ,24 ]. ...
... ,
24 ,
80 ~
87 ])
<strong>Schematic diagrams illustrating the global ocean isotopic mass balance of Cu </strong>(<strong>Data from references </strong>[<strong>23</strong>,<strong>24</strong>,<strong>80~87</strong>]) Fig.3 ![]()
铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
... 铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... [24 ]利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... [24 ].Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... [24 ]的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... 海水中铜的稳定同位素作为一个新的指标,可以进一步推断海洋中铜的源汇格局.通过比较现代海洋输入和输出的铜通量及其平均铜同位素值,发现了海水中典型铜汇的同位素值(约+0.3‰)相较于铜源的同位素值(约+0.6‰)偏轻.假设海洋铜循环处于稳态,通过同位素质量守恒计算出来的海洋体系中铜的收支是不平衡的[23 ] ,这意味着当代海洋存在至今未被确认的轻铜同位素的源[5 ,23 ,98 ] ,其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
Regulated copper uptake in chlamydomonas reinhardtii in response to copper availability
2
1996
... 铜对海洋生物具有双重作用,铜是生命必需元素,但超出需求浓度范围的铜又具有毒性[10 ] .铜作为氧化还原过渡金属,其离子态能与蛋白质形成稳定的化合物,通过氧化还原反应转移电子[25 ] ,这些性质使得铜具有丰富的生物学作用,参与到海洋生物的生理过程,最终影响海洋生态系统.图1 归纳总结了海洋中铜参与的生物地球化学过程及其海洋生理生态功能. ...
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
The involvement of a multicopper oxidase in iron uptake by the green algae chlamydomonas reinhardtii
1
2002
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
Hydroxylamine as an intermediate in ammonia oxidation by globally abundant marine archaea
1
2013
... 海洋中铜是重要的生命元素.铜可作为海洋浮游植物体内酶的金属辅基,参与催化一系列氧化还原反应.光合自养生物所需的大部分铜都用于与光合作用有关的电子转运蛋白和酶[12 ] ,如硅藻利用含铜蛋白——质体蓝素,用于光合作用的电子转移[13 ] .含铜和铁的细胞色素氧化酶能协助线粒体输运电子,将O2 还原成H2 O[11 ] .含铜的超氧化物歧化酶(Superoxide Dismutase,SOD)能协助浮游植物清除在环境胁迫中产生的过多活性氧[14 ] .衣藻细胞膜表面的含多铜氧化酶与含多铜氧化酶(Multi-Cu Oxidases,MCOs)蛋白质序列相似,能将Fe(ΙΙ)氧化成Fe(ΙΙΙ),从而实现衣藻对铁的高亲和力吸收[26 ] .除此之外,铜还催化硝化反应的第一步——氨氧化反应:氨氧化细菌和古菌体内的氨单加氧铜酶将NH3 催化氧化成NH2 OH[15 ,27 ] .在反硝化反应中,一氧化二氮铜基还原酶能将N2 O催化还原成N2 [16 ] .浓度适宜的铜能对海洋生物的生理功能起到促进作用,在一定程度上提高海洋的初级生产力,从而影响全球的氮循环和碳循环[4 ,13 ] . ...
Copper requirements of the ammonia-oxidizing archaeon Nitrosopumilus maritimus SCM1 and implications for nitrification in the marine environment
1
2013
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
Mechanism of toxicity of ionic copper and copper complexes to algae
1
1987
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
Physiological responses of coastal phytoplankton (Visakhapatnam,SW Bay of Bengal,India) to experimental copper addition
2
2017
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Coastal strains of marine Synechococcus species exhibit increased tolerance to copper shock and a distinctive transcriptional response relative to those of open-ocean strains
1
2009
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
Copper and photosystem II: A controversial relationship
1
1995
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
Copper complexation in the Northeast Pacific
2
1988
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
... van den Berg[71 ] 在1984年首次提出使用竞争配体交换—阴极溶出伏安法(CLE-CSV)精确测定铜配体的条件稳定常数.根据条件稳定常数,铜的有机配体可分为2类:一类是浓度较低的强配体L1 (log10 K΄Cu-L1 =12~14,[CuL1 ]=1.8~13 nmol/L),另一类是浓度较高的弱配体L2 (log10 K΄Cu-L2 =9~12,[CuL2 ]=8~30 nmol/L[33 ,72 ,73 ] ).图2 显示了铜及其配体在北太平洋东部一个站位的垂直剖面.总铜和自由铜离子表现出了表层亏损而深层富集的营养盐分布的模式;铜配体则在表层有个极大值,随深度加深后迅速降低随后在深层海水富集.近表层海水中较高的配体浓度对应着较低的自由铜离子浓度,一方面与生物利用表层海水的总溶解铜离子有关,另一方面可能是配体络合自由铜离子造成其浓度降低.Moffett等[53 ] 认为在200~300 m水深以下,强有机配体的络合作用的重要性有所减弱,因为配体在300 m水深以下都维持相对较高的浓度,甚至在3 000 m以下,Cu还能被有机配体强烈络合.铜的强有机配体L1 浓度与叶绿素浓度的垂直分布存在一致性[53 ] ,推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
Interactions among Cu2+ ,Zn2+ ,and Mn2+ in controlling cellular Mn,Zn,and growth rate in the coastal alga Chlamydomonas
2
1998
... 超出细胞需求的铜会对海洋浮游植物产生毒害作用.高浓度铜能催化活性氧的形成[28 ] ,还会与-SH结合或与O 2 - [29 ] .Cu2+ 会取代卟啉环中的Mg2+ ,破坏叶绿素的分子结构[30 ] .高浓度的铜会降低浮游植物与光合作用相关的基因转录水平[31 ] ,干扰色素和脂质的合成,影响叶绿体超微结构,从而对海洋光合效率产生负面影响[32 ] .铜还会在酶的金属辅基结合位点[33 ] 或者在细胞内的代谢部位[34 ] 对其他金属造成竞争性抑制.高浓度铜会影响浮游植物的光合作用,对营养盐等必需元素的吸收,脂质、色素和酶的生物合成等生理过程,并最终产生相应的生态学效应. ...
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Dynamics of phytoplankton community structure in the South China Sea in response to the East Asian aerosol input
1
2011
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Reduction and transport of organically bound iron by Thalassiosira oceanica (Bacillariophyceae)
2
2001
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
... [36 ]和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Utilization of iron bound to strong organic ligands by plankton communities in the subarctic Pacific Ocean
1
1999
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Copper‐dependent iron transport in coastal and oceanic diatoms
2
2006
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
... [38 ].此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
A general kinetic model for iron acquisition by eukaryotic phytoplankton
1
2005
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Synthesis of iron fertilization experiments: From the iron age in the age of Enlightenment
1
2005
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Interactive effects of external manganese,the toxic metals copper and zinc,and light in controlling cellular manganese and growth in a coastal diatom
1
1998
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
The interaction between zinc deficiency and copper toxicity as it affects the silicic acid uptake mechanisms in Thalassiosira pseudonana1
1
1981
... 海洋中铜会与其他元素发生协同和拮抗等相互作用.在铁限制海域,铜能替代铁执行部分生理功能:例如硅藻等浮游植物使用含铜的质体蓝素来替代含铁的细胞色素C6 氧化酶[13 ] ;用Cu-SOD来替代Fe-SOD[35 ] .在铁限制的条件下,浮游植物通过上调高亲和力铁吸收系统(High-Affinity Fe Transport System,HAFeTS),获得足够的铁以满足其生长需要[36 ,37 ] .而HAFeTS系统则由含多铜铁氧化酶[38 ] 、Fe(ΙΙΙ)还原酶[36 ] 和Fe(ΙΙ)通透酶[39 ] 组成.对硅藻T. pseudonana 的研究表明,在铁限制下硅藻细胞对含多铜铁氧化酶的转录水平显著提高,而加铜组MCO的转录水平显著高于减铜组[38 ] .此类铜的协同作用可以通过增加浮游植物对铜的吸收,以减少浮游植物对铁的需求或增强对铁的吸收能力,以维持浮游植物正常的生理过程.众所周知,铁会限制占世界海洋25%面积的高营养盐、低叶绿素(High Nitrate Low Chlorophyll,HNLC)海区的浮游植物生长[40 ] ,铜与铁的协同作用在HNLC海区的生态效应还有待进一步深入研究.铜也会影响海洋浮游植物对其他金属的吸收,如高浓度的Cu(ΙΙ)会抑制衣藻细胞和硅藻细胞对海水锰的吸收以及减缓对锌的吸收速率[34 ,41 ] .铜和锌的交互作用影响硅藻对硅的吸收,研究发现硅藻T. pseudonana 中铜和硅酸的吸收活性位点存在不可逆的竞争关系,而锌可以协助参与硅酸的吸收,锌的存在可以缓解铜对硅酸盐吸收的抑制作用[42 ] .其他金属以及氮、磷等营养盐水平也会影响铜毒性的表达,例如铁的共富集作用能缓解铜对浮游植物的毒性[30 ] .铜可以影响浮游植物对其他金属的吸收,同时铁也能缓解铜的毒性,这些过程反映了铜与其他金属在浮游植物生理代谢过程中的交互作用. ...
Copper uptake kinetics in diverse marine phytoplankton
2
2006
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
... [43 ,48 ];近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
Responses of diverse heterotrophic bacteria to elevated copper concentrations
2
1994
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
... [44 ,47 ].浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
Production of phytochelatins and glutathione by marine phytoplankton in response to metal stress
1
2006
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
Production of strong,extracellular Cu chelators by marine cyanobacteria in response to Cu stress
2
1996
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Export of cadmium and phytochelatin by the marine diatom Thalassiosira weissflogii
1
1996
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
Sensitivity of marine microalgae to copper: The effect of biotic factors on copper adsorption and toxicity
1
2007
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
Copper requirements for iron acquisition and growth of coastal and oceanic diatoms
1
2005
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
The natural selection of the chemical elements
1
1997
... 浮游植物也进化出一些应对铜毒性的解毒机制:一是通过减少细胞膜表面的活性铜吸收位点,或者浮游植物可以通过增加细胞体积以降低细胞表面积比,以降低对铜的吸收速率[43 ] ;二是通过释放胞外有机配体与铜络合以减少海水环境中的自由铜离子的浓度[44 ~46 ] ;三是通过外排机制移除细胞内形成的铜络合物[44 ,47 ] .浮游植物对海水中铜的耐受性存在物种差异,例如,聚球藻对铜浓度最敏感,颗石藻和甲藻次之,硅藻最不敏感,更能够耐受海洋环境中铜浓度的提高[10 ] .浮游植物通过细胞膜的特殊转运配体吸收铜离子,这一吸收过程受到光和温度等环境因子的影响[25 ] .生物和环境因素中铜的浓度相互作用,决定了浮游植物的群落组成.例如,体型较大的浮游植物由于低的细胞表面积比而更能抵抗铜的毒性,可以在铜浓度较高的海区生存[43 ,48 ] ;近岸生活的浮游生物物种往往比大洋物种更耐受铜毒性[49 ] ,这是因为藻类在较高铜浓度的近岸海水中进化出更强的耐铜机制[50 ] . ...
Measurement of copper(I) in surface waters of the subtropical Atlantic and Gulf of Mexico
1
1988
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Interactive influences of bioactive trace metals on biological production in oceanic waters
1
1991
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Cu complexation by organic ligands in the sub-arctic NW Pacific and Bering Sea
5
2007
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
... van den Berg[71 ] 在1984年首次提出使用竞争配体交换—阴极溶出伏安法(CLE-CSV)精确测定铜配体的条件稳定常数.根据条件稳定常数,铜的有机配体可分为2类:一类是浓度较低的强配体L1 (log10 K΄Cu-L1 =12~14,[CuL1 ]=1.8~13 nmol/L),另一类是浓度较高的弱配体L2 (log10 K΄Cu-L2 =9~12,[CuL2 ]=8~30 nmol/L[33 ,72 ,73 ] ).图2 显示了铜及其配体在北太平洋东部一个站位的垂直剖面.总铜和自由铜离子表现出了表层亏损而深层富集的营养盐分布的模式;铜配体则在表层有个极大值,随深度加深后迅速降低随后在深层海水富集.近表层海水中较高的配体浓度对应着较低的自由铜离子浓度,一方面与生物利用表层海水的总溶解铜离子有关,另一方面可能是配体络合自由铜离子造成其浓度降低.Moffett等[53 ] 认为在200~300 m水深以下,强有机配体的络合作用的重要性有所减弱,因为配体在300 m水深以下都维持相对较高的浓度,甚至在3 000 m以下,Cu还能被有机配体强烈络合.铜的强有机配体L1 浓度与叶绿素浓度的垂直分布存在一致性[53 ] ,推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
... [53 ],推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
... )铜配体、总铜离子和自由铜离子浓度剖面(据参考文献[
53 ]修改)
<strong>Profiles of ligand</strong>,<strong>total dissolved Cu and free Cu<sup>2+</sup> for station in the North Pacific </strong>(<strong>47°01′N</strong>,<strong> 170°30′W</strong>,<strong>modified after reference </strong>[<xref ref-type="bibr" rid="R53">53</xref>]) Fig.2 ![]()
海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
... [
53 ])
Fig.2 ![]()
海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
A review of total dissolved copper and its chemical speciation in San Francisco Bay,California
2
2007
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Evaluation of the CuCl+ stability constant and molar absorptivity in aqueous media
1
1983
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Chemical speciation of environmentally significant metals with inorganic ligands - Part 2: The Cu2+ -OH- ,Cl- ,CO3 2 - 4 2 - 4 3 -
1
2007
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Chemistry of superoxide radical in seawater: Reactions with organic Cu complexes
1
2000
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Chemistry of the Superoxide Radical (O2- ) in Seawater: Reactions with Inorganic Copper Complexes
2
1998
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
... [58 ].Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Oxidation of copper(I) in seawater at nanomolar levels
1
2009
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Oxidation kinetics of Cu(I) in seawater: Implications for its existence in the marine environment
1
1983
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Evidence for strong copper(I) complexation by organic ligands in seawater
1
1998
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Paired dissolved and particulate phase Cu isotope distributions in the South Atlantic
1
2018
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Size distribution of particulate trace elements in the U.S. GEOTRACES Eastern Pacific Zonal Transect (GP16)
1
2018
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Exposing the distributions and elemental associations of scavenged particulate phases in the ocean using basin-scale multi-element data sets
1
2019
... 铜在海洋中表现出不同的形态.在海水中,铜的价态表现为Cu(Ι)和Cu(ΙΙ),常见的形态有溶解态、络合态和颗粒态.在当代氧化的海洋水体,铜主要以氧化态的Cu(II)存在,通常占总铜的90%~95%[51 ] .有机络合是海洋中铜的主要络合形式,铜在海洋中的有机络合程度最高值一般出现在海洋生产力最大值附近[52 ] .在上层海洋,铜与有机物络合比例高达99%[53 ] .在显著受到人类活动影响的近岸水体,铜与有机物络合的比例可高达99.9%以上[54 ] .海水中的铜离子也可与无机配体络合,其中Cu(Ι)以氯离子络合为主[55 ] ,而Cu(ΙΙ)则以碳酸盐络合为主[56 ] .Cu(I)可通过光化学过程和海底沉积物释放等过程产生.光化学作用产生的超氧化物HO2 或O 2 - [57 ,58 ] ,有研究报道了阳光照射的表层海水中Cu(I)占总铜的比例可高达25%[58 ] .Cu(I)通常以C u C l 2 - [59 ] ,氯离子的络合作用能使Cu(I)保持稳定[60 ] ,谷胱甘肽和半胱氨酸等硫醇也能络合Cu(I)稳定存在[61 ] .开阔大洋颗粒态的铜的含量通常为10~500 pmol/kg,垂向分布与溶解态类似表现为表层亏损并在深层富集[62 ,63 ] ,通常认为海水中的铁氧化物和氢氧化物水合物是控制颗粒态铜含量的主控因素[64 ] . ...
Testing the underlying chemical principles of the Biotic Ligand Model (BLM) to marine copper systems: Measuring copper speciation using fluorescence quenching
1
2018
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
Metal speciation in natural waters with emphasis on reduced sulfur groups as strong metal binding sites
1
2002
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
Copper-induced release of complexing ligands similar to thiols by Emiliania huxleyi in seawater cultures
1
1999
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
Methanobactin,a copper-acquisition compound from methane-oxidizing bacteria
1
2004
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
Primary- and secondary-structural analysis of a unique prokaryotic metallothionein from a Synechococcus sp. cyanobacterium
1
1988
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
Novel speciation analysis of copper in river water: Observation of soluble anionic copper-ligand complexes
2
2016
... 海水中铜的有机配体主要来自浮游植物的代谢产物和陆源物质[65 ] .常见的天然有机物含有许多铜的结合位点,如氨基、羧基、酚基和硫基等官能团[66 ] .浮游植物产生铜配体的类型体现着物种差异性.例如,在球石藻产生的配体中,硫醇占大部分[67 ] ,而甲烷氧化细菌Methylosinus trichosporium OB3b能产生类似于铁载体的胞外铜转运分子——甲烷氧化菌素[68 ] .海洋中的真核和原核微生物都能产生富含半胱氨酸的类金属硫蛋白来络合胞内有毒金属[69 ] ,培养实验还从真核浮游植物中分离出谷胱甘肽和植物螯合素等有机配体[45 ] .这些有机配体的存在不仅可以将游离铜离子活性维持在海洋生物可接受范围内,同时限制下沉颗粒物对生物活性金属铜的清除[17 ] .在河流和河口区域,铜能与陆源物质发生络合反应,包括了腐殖酸、富里酸等天然有机质以及与人类活动释放有机物质(如EDTA等)[70 ] . ...
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Determination of the complexing capacity and conditional stability constants of complexes of copper(II) with natural organic ligands in seawater by cathodic stripping voltammetry of copper-catechol complex ions
1
1984
... van den Berg[71 ] 在1984年首次提出使用竞争配体交换—阴极溶出伏安法(CLE-CSV)精确测定铜配体的条件稳定常数.根据条件稳定常数,铜的有机配体可分为2类:一类是浓度较低的强配体L1 (log10 K΄Cu-L1 =12~14,[CuL1 ]=1.8~13 nmol/L),另一类是浓度较高的弱配体L2 (log10 K΄Cu-L2 =9~12,[CuL2 ]=8~30 nmol/L[33 ,72 ,73 ] ).图2 显示了铜及其配体在北太平洋东部一个站位的垂直剖面.总铜和自由铜离子表现出了表层亏损而深层富集的营养盐分布的模式;铜配体则在表层有个极大值,随深度加深后迅速降低随后在深层海水富集.近表层海水中较高的配体浓度对应着较低的自由铜离子浓度,一方面与生物利用表层海水的总溶解铜离子有关,另一方面可能是配体络合自由铜离子造成其浓度降低.Moffett等[53 ] 认为在200~300 m水深以下,强有机配体的络合作用的重要性有所减弱,因为配体在300 m水深以下都维持相对较高的浓度,甚至在3 000 m以下,Cu还能被有机配体强烈络合.铜的强有机配体L1 浓度与叶绿素浓度的垂直分布存在一致性[53 ] ,推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
Organic complexation and its control of the dissolved concentrations of copper and zinc in the Scheldt estuary
1
1987
... van den Berg[71 ] 在1984年首次提出使用竞争配体交换—阴极溶出伏安法(CLE-CSV)精确测定铜配体的条件稳定常数.根据条件稳定常数,铜的有机配体可分为2类:一类是浓度较低的强配体L1 (log10 K΄Cu-L1 =12~14,[CuL1 ]=1.8~13 nmol/L),另一类是浓度较高的弱配体L2 (log10 K΄Cu-L2 =9~12,[CuL2 ]=8~30 nmol/L[33 ,72 ,73 ] ).图2 显示了铜及其配体在北太平洋东部一个站位的垂直剖面.总铜和自由铜离子表现出了表层亏损而深层富集的营养盐分布的模式;铜配体则在表层有个极大值,随深度加深后迅速降低随后在深层海水富集.近表层海水中较高的配体浓度对应着较低的自由铜离子浓度,一方面与生物利用表层海水的总溶解铜离子有关,另一方面可能是配体络合自由铜离子造成其浓度降低.Moffett等[53 ] 认为在200~300 m水深以下,强有机配体的络合作用的重要性有所减弱,因为配体在300 m水深以下都维持相对较高的浓度,甚至在3 000 m以下,Cu还能被有机配体强烈络合.铜的强有机配体L1 浓度与叶绿素浓度的垂直分布存在一致性[53 ] ,推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
Speciation of dissolved copper and nickel in South San Francisco Bay: A multi-method approach
2
1994
... van den Berg[71 ] 在1984年首次提出使用竞争配体交换—阴极溶出伏安法(CLE-CSV)精确测定铜配体的条件稳定常数.根据条件稳定常数,铜的有机配体可分为2类:一类是浓度较低的强配体L1 (log10 K΄Cu-L1 =12~14,[CuL1 ]=1.8~13 nmol/L),另一类是浓度较高的弱配体L2 (log10 K΄Cu-L2 =9~12,[CuL2 ]=8~30 nmol/L[33 ,72 ,73 ] ).图2 显示了铜及其配体在北太平洋东部一个站位的垂直剖面.总铜和自由铜离子表现出了表层亏损而深层富集的营养盐分布的模式;铜配体则在表层有个极大值,随深度加深后迅速降低随后在深层海水富集.近表层海水中较高的配体浓度对应着较低的自由铜离子浓度,一方面与生物利用表层海水的总溶解铜离子有关,另一方面可能是配体络合自由铜离子造成其浓度降低.Moffett等[53 ] 认为在200~300 m水深以下,强有机配体的络合作用的重要性有所减弱,因为配体在300 m水深以下都维持相对较高的浓度,甚至在3 000 m以下,Cu还能被有机配体强烈络合.铜的强有机配体L1 浓度与叶绿素浓度的垂直分布存在一致性[53 ] ,推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Distribution of copper-complexing ligands in Canadian Arctic waters as determined by immobilized copper(II)-ion affinity chromatography
1
2019
... van den Berg[71 ] 在1984年首次提出使用竞争配体交换—阴极溶出伏安法(CLE-CSV)精确测定铜配体的条件稳定常数.根据条件稳定常数,铜的有机配体可分为2类:一类是浓度较低的强配体L1 (log10 K΄Cu-L1 =12~14,[CuL1 ]=1.8~13 nmol/L),另一类是浓度较高的弱配体L2 (log10 K΄Cu-L2 =9~12,[CuL2 ]=8~30 nmol/L[33 ,72 ,73 ] ).图2 显示了铜及其配体在北太平洋东部一个站位的垂直剖面.总铜和自由铜离子表现出了表层亏损而深层富集的营养盐分布的模式;铜配体则在表层有个极大值,随深度加深后迅速降低随后在深层海水富集.近表层海水中较高的配体浓度对应着较低的自由铜离子浓度,一方面与生物利用表层海水的总溶解铜离子有关,另一方面可能是配体络合自由铜离子造成其浓度降低.Moffett等[53 ] 认为在200~300 m水深以下,强有机配体的络合作用的重要性有所减弱,因为配体在300 m水深以下都维持相对较高的浓度,甚至在3 000 m以下,Cu还能被有机配体强烈络合.铜的强有机配体L1 浓度与叶绿素浓度的垂直分布存在一致性[53 ] ,推测叶绿素最大层中存在的浮游植物和蓝藻可能是产生铜配体的主要来源[74 ] . ...
Processes regulating cellular metal accumulation and physiological effects: Phytoplankton as model systems
1
1998
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Uptake of lipophilic organic Cu,Cd,and Pb complexes in the coastal diatom Thalassiosira weissflogii
1
1994
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Production of extracellular Cu complexing ligands by eucaryotic phytoplankton in response to Cu stress
1
2000
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
The effects of Cu and Fe availability on the growth and Cu∶C ratios of marine diatoms
1
2008
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
Effect of ocean acidification on the speciation of metals in seawater
1
2009
... 海洋中铜的形态影响了铜的生物可利用性.自由铜离子可直接被吸收利用[10 ,75 ] ,低分子量的亲脂性铜和无机态铜较易被吸收[54 ,76 ] ,而高分子量有机质(如腐殖质)结合的铜、胶体态和颗粒态铜的生物可利用性则较低[73 ] .铜的毒性主要由游离二价铜离子浓度主导,当Cu(ΙΙ)浓度达到10-12 nmol/L,就会对浮游植物产生毒性[10 ] ,有机配体与铜离子的络合作用能减轻由游离铜离子引起的抑制藻类生长的毒害作用.研究表明,对铜毒性越敏感的浮游植物,往往产生与铜络合能力越强的配体[77 ] .例如,铜暴露培养实验结果表明,对铜毒性高度敏感的聚球藻会释放与L1 强度相近的胞外配体[46 ] .然而,过度的铜络合对海洋生态系统也会产生负面效应.例如:过度络合会导致铜毒性的降低进而导致水体中藻类的爆发[70 ] ;过度络合会抑制含多铜二价铁氧化酶的合成,降低高亲和铁吸收系统的所需铁的可利用性,从而影响浮游植物对铁的吸收[78 ] .综上所述,海洋中铜的形态多样且各自具有不同的生物有效性,在全球气候变化背景下,增加了铜对海洋生态未来影响趋势预测的难度.例如:日益严重的海洋酸化现象可能会增加海洋中游离铜离子的浓度,对初级生产力造成不良影响;但海洋酸化也可能增加溶解态铁浓度,从而刺激初级生产力[79 ] . ...
A global assessment of copper,zinc,and lead isotopes in mineral dust sources and aerosols
2
2020
...
图3 总结了大洋铜的主要源汇过程及其通量和同位素组成特征.河流是大洋铜的主要来源,每年贡献0.60~6.70 Gmol的铜
[23 ,24 ] ;大气沉降次之,每年贡献0.054~0.270 Gmol的铜
[23 ,24 ] ;海底沉积物再矿化以每年约0.37 Gmol的通量补充海洋铜库
[24 ] ;热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出
[23 ,24 ] .
图3 全球海洋铜质量平衡示意图及源汇物质的<strong><i>δ</i><sup>65</sup>Cu</strong>值(数据来自参考文献[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R80">80</xref>~<xref ref-type="bibr" rid="R87">87</xref>]) <strong>Schematic diagrams illustrating the global ocean isotopic mass balance of Cu </strong>(<strong>Data from references </strong>[<strong>23</strong>,<strong>24</strong>,<strong>80~87</strong>]) Fig.3 ![]()
铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
Isotopic constraints on biogeochemical cycling of copper in the ocean
2014
Copper isotope signatures in modern marine sediments
1
2017
... 海水中铜的稳定同位素作为一个新的指标,可以进一步推断海洋中铜的源汇格局.通过比较现代海洋输入和输出的铜通量及其平均铜同位素值,发现了海水中典型铜汇的同位素值(约+0.3‰)相较于铜源的同位素值(约+0.6‰)偏轻.假设海洋铜循环处于稳态,通过同位素质量守恒计算出来的海洋体系中铜的收支是不平衡的[23 ] ,这意味着当代海洋存在至今未被确认的轻铜同位素的源[5 ,23 ,98 ] ,其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
Determination of the copper isotope composition of seawater revisited: A case study from the Mediterranean Sea
2
2019
... 应用GEOTRACES IDP2017
[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如
图4 所示.如
表1 和
图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放
[103 ] 和洋流输运
[21 ,104 ] 等因素有关.
图4 溶解态铜在世界大洋表层的现有数据分布(<strong>0~50 m</strong>)(数据来自参考文献[<xref ref-type="bibr" rid="R23">23</xref>, <xref ref-type="bibr" rid="R83">83</xref>,<xref ref-type="bibr" rid="R84">84</xref>, <xref ref-type="bibr" rid="R99">99</xref>~<xref ref-type="bibr" rid="R102">102</xref>]) The existing data of dissolved copper in the world surface ocean (0~50 m) (data from references [23, 83,84, 99~102]) Fig.4 ![]()
表1 不同海区、不同深度的海水溶解态铜浓度 ...
... Dissolved copper concentration in different oceans and depth
Table 1 水体类型 区域 铜浓度/(nmol/kg) n 参考文献 表层水体:开阔大洋 北太平洋 1.72±0.82 16 [99 ] 北大西洋 0.88±0.28 20 [99 ] 印度洋 1.46±0.64 10 [99 ] 南太平洋 0.53±0.26 135 [99 ] 南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ]
海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
Dissolved copper isotope biogeochemistry in the Tasman Sea,SW Pacific Ocean
1
2014
... 应用GEOTRACES IDP2017
[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如
图4 所示.如
表1 和
图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放
[103 ] 和洋流输运
[21 ,104 ] 等因素有关.
图4 溶解态铜在世界大洋表层的现有数据分布(<strong>0~50 m</strong>)(数据来自参考文献[<xref ref-type="bibr" rid="R23">23</xref>, <xref ref-type="bibr" rid="R83">83</xref>,<xref ref-type="bibr" rid="R84">84</xref>, <xref ref-type="bibr" rid="R99">99</xref>~<xref ref-type="bibr" rid="R102">102</xref>]) The existing data of dissolved copper in the world surface ocean (0~50 m) (data from references [23, 83,84, 99~102]) Fig.4 ![]()
表1 不同海区、不同深度的海水溶解态铜浓度 ...
The stable isotope geochemistry of copper and zinc
2004
Determination of natural Cu-isotope variation by plasma-source mass spectrometry: Implications for use as geochemical tracers
1
2000
... 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
Sources of particulate Ni and Cu in the water column of the northern South China Sea: Evidence from elemental and isotope ratios in aerosols and sinking particles
2
2020
...
图3 总结了大洋铜的主要源汇过程及其通量和同位素组成特征.河流是大洋铜的主要来源,每年贡献0.60~6.70 Gmol的铜
[23 ,24 ] ;大气沉降次之,每年贡献0.054~0.270 Gmol的铜
[23 ,24 ] ;海底沉积物再矿化以每年约0.37 Gmol的通量补充海洋铜库
[24 ] ;热液口的输入和陆源沉积物等来源也逐步被认为对海洋铜库有贡献,但其通量仍需进一步研究.铜通常以颗粒沉降的形式从水体中去除,每年以0.49~0.54 Gmol的通量输出
[23 ,24 ] .
图3 全球海洋铜质量平衡示意图及源汇物质的<strong><i>δ</i><sup>65</sup>Cu</strong>值(数据来自参考文献[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R80">80</xref>~<xref ref-type="bibr" rid="R87">87</xref>]) <strong>Schematic diagrams illustrating the global ocean isotopic mass balance of Cu </strong>(<strong>Data from references </strong>[<strong>23</strong>,<strong>24</strong>,<strong>80~87</strong>]) Fig.3 ![]()
铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
... 海水中铜的稳定同位素作为一个新的指标,可以进一步推断海洋中铜的源汇格局.通过比较现代海洋输入和输出的铜通量及其平均铜同位素值,发现了海水中典型铜汇的同位素值(约+0.3‰)相较于铜源的同位素值(约+0.6‰)偏轻.假设海洋铜循环处于稳态,通过同位素质量守恒计算出来的海洋体系中铜的收支是不平衡的[23 ] ,这意味着当代海洋存在至今未被确认的轻铜同位素的源[5 ,23 ,98 ] ,其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
The mechanism of iron removal in estuaries
1
1977
... 铜本身的海洋生物地球化学过程复杂,陆—海、海—气等界面的交互作用也很复杂,这导致目前对大洋中铜源汇格局的估算仍存在着不确定性.陆—海界面的河口区域,铜的行为在河海交界处受盐度驱动的絮凝、混凝和颗粒沉降等过程的控制[88 ] ,这增加了估算河流中的铜入海通量的难度.例如,Roshan等[22 ] 发现北美沿岸至北大西洋环流的表层水表现出dCu浓度与盐度较好的线性关系(R 2 =0.93),表明了该海域表层水主要受控于河流输入,且铜的清除速率低于水体横向混合速率.Richon等[24 ] 通过河流的铜同位素组成,利用模型估计全球河流铜通量为6.7 Gmol/a,是Little等[23 ] 估计的(0.6~0.8 Gmol/a)10倍左右. ...
Factors controlling the solubility of aerosol trace metals in the atmosphere and on mixing into seawater
1
1995
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
Global iron connections between desert dust,ocean biogeochemistry,and climate
1
2005
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
Toxicity of atmospheric aerosols on marine phytoplankton
2
2009
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
... [91 ],使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
Dissolution and solubility of trace metals from natural and anthropogenic aerosol particulate matter
1
2005
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
On the fractional solubility of copper in marine aerosols: Toxicity of aeolian copper revisited
1
2010
... 在海—气界面,通过干、湿沉降进入海洋的铜,其溶解度受控于pH主导的颗粒—水界面的吸附—解吸过程[89 ] .铜的人为来源和天然来源差异会影响其在海洋表面分布的均匀性,例如北半球径流较多、人口密集,因此从河流和大气接收到外部输入铜会比南半球多[24 ] .Little等[23 ] 基于Jickells等[90 ] 的数据,其中大气尘埃沉降通量为450 Mt/a,以地壳平均铜含量(28×10-6 )代表大气尘埃铜含量,以天然气溶胶中铜的溶解度(27%)代表大气尘埃中铜的溶解度,最后估算出大气尘埃铜通量为0.054 Gmol/a.而Richon等[24 ] 利用PISCES模型模拟了全球气溶胶铜沉降通量,辅以Paytan等[91 ] 估算的海洋气溶胶(天然与人为气溶胶)铜的溶解度为40%,最终估算出人为气溶胶铜通量为0.08 Gmol/a,天然气溶胶铜通量为0.19 Gmol/a[24 ] .Richon等[24 ] 的估算结果比Little等[23 ] 的高出500%,二者的差异主要由气溶胶通量和铜的溶解度因选取区域和气溶胶性质不同而造成的.由于现有观测数据、模拟参数、气候条件以及沙漠沙尘和人为来源粒子等条件上的差异[91 ] ,使得不同全球沉降模型的估算结果存在一定差异.而铜的溶解度因气溶胶类型不同而存在较大差异,在天然气溶胶中,铜的溶解度为1%~27%,而铜在人为气溶胶中的溶解度可高达75%~98%[92 ,93 ] .模型估算和溶解度的差异是估算大气沉降铜输送的误差来源.图3 b列出了部分区域的气溶胶铜同位素的组成,δ 65 Cu整体上分布在0~0.4‰,铜同位素主要受控于气溶胶颗粒的粒径大小,与气溶胶类型的相关性不明显,一般小颗粒和大颗粒的铜同位素偏重,而中等颗粒的铜同位素偏轻[80 ] . ...
Geochemistry and origin of the Holocene sapropel in the Black Sea
1
1990
... 已有的知识通常认为海洋是铜的汇.在氧化水体中,铜会吸附在颗粒物表面或与之沉淀,从而沉降到海底,Little等[23 ] 估计沉降到氧化沉积物中的铜通量为0.49 Gmol/a.在还原海水中,铜会与硫化物发生沉淀作用,从水体中去除,例如在总硫化物含量达到100 μmol/L的黑海深处,铜和锌几乎从海水中分离,相应地在海底沉积物中富集[94 ] .然而,沉积物—海界面铜的生物地球化学过程复杂,源汇格局仍需要进一步研究[95 ] .例如:在低氧沉积物中,微生物介导的铁、锰氧化物的还原性溶解能释放与氧结合的铜[96 ] ,成为海水中铜的来源.在无氧沉积物中,细菌还原硫酸盐产生的硫化物会与铁形成铁硫化物,再进一步与铜共沉淀[97 ] . ...
Geochemistry of recent oxic and anoxic marine sediments: Implications for the geological record
1
1993
... 已有的知识通常认为海洋是铜的汇.在氧化水体中,铜会吸附在颗粒物表面或与之沉淀,从而沉降到海底,Little等[23 ] 估计沉降到氧化沉积物中的铜通量为0.49 Gmol/a.在还原海水中,铜会与硫化物发生沉淀作用,从水体中去除,例如在总硫化物含量达到100 μmol/L的黑海深处,铜和锌几乎从海水中分离,相应地在海底沉积物中富集[94 ] .然而,沉积物—海界面铜的生物地球化学过程复杂,源汇格局仍需要进一步研究[95 ] .例如:在低氧沉积物中,微生物介导的铁、锰氧化物的还原性溶解能释放与氧结合的铜[96 ] ,成为海水中铜的来源.在无氧沉积物中,细菌还原硫酸盐产生的硫化物会与铁形成铁硫化物,再进一步与铜共沉淀[97 ] . ...
The adsorption of Cu,Mn and Zn by iron oxyhydroxide in model estuarine solutions
1
1982
... 已有的知识通常认为海洋是铜的汇.在氧化水体中,铜会吸附在颗粒物表面或与之沉淀,从而沉降到海底,Little等[23 ] 估计沉降到氧化沉积物中的铜通量为0.49 Gmol/a.在还原海水中,铜会与硫化物发生沉淀作用,从水体中去除,例如在总硫化物含量达到100 μmol/L的黑海深处,铜和锌几乎从海水中分离,相应地在海底沉积物中富集[94 ] .然而,沉积物—海界面铜的生物地球化学过程复杂,源汇格局仍需要进一步研究[95 ] .例如:在低氧沉积物中,微生物介导的铁、锰氧化物的还原性溶解能释放与氧结合的铜[96 ] ,成为海水中铜的来源.在无氧沉积物中,细菌还原硫酸盐产生的硫化物会与铁形成铁硫化物,再进一步与铜共沉淀[97 ] . ...
Adsorption and coprecipitation of divalent metals with mackinawite (FeS)
1
1993
... 已有的知识通常认为海洋是铜的汇.在氧化水体中,铜会吸附在颗粒物表面或与之沉淀,从而沉降到海底,Little等[23 ] 估计沉降到氧化沉积物中的铜通量为0.49 Gmol/a.在还原海水中,铜会与硫化物发生沉淀作用,从水体中去除,例如在总硫化物含量达到100 μmol/L的黑海深处,铜和锌几乎从海水中分离,相应地在海底沉积物中富集[94 ] .然而,沉积物—海界面铜的生物地球化学过程复杂,源汇格局仍需要进一步研究[95 ] .例如:在低氧沉积物中,微生物介导的铁、锰氧化物的还原性溶解能释放与氧结合的铜[96 ] ,成为海水中铜的来源.在无氧沉积物中,细菌还原硫酸盐产生的硫化物会与铁形成铁硫化物,再进一步与铜共沉淀[97 ] . ...
The copper isotope geochemistry of rivers and the oceans
1
2008
... 海水中铜的稳定同位素作为一个新的指标,可以进一步推断海洋中铜的源汇格局.通过比较现代海洋输入和输出的铜通量及其平均铜同位素值,发现了海水中典型铜汇的同位素值(约+0.3‰)相较于铜源的同位素值(约+0.6‰)偏轻.假设海洋铜循环处于稳态,通过同位素质量守恒计算出来的海洋体系中铜的收支是不平衡的[23 ] ,这意味着当代海洋存在至今未被确认的轻铜同位素的源[5 ,23 ,98 ] ,其同位素接近于岩石组成(约为0)(图3 ).Takano等[87 ] 测得的大气尘埃的铜同位素组成为0,并通过箱式模型估算气溶胶铜通量为0.96 Gmol/a,这比Little等[23 ] 和Richon等[24 ] 的估算结果大了1~2个数量级,与河流输入的通量相当,因此可以作为缺失铜源的可能之一,填补目前大洋铜同位素的不平衡.Little等[82 ] 通过低氧还原沉积环境的δ 65 Cu值,估算缺失铜源的通量为0.28 Gmol/a,并猜测可能是热液输入或陆架衍生颗粒物的部分溶解.随着未来全球铜库同位素数据的完善,海洋源汇格局可得到进一步完善. ...
The GEOTRACES Intermediate Data Product 2017
14
2018
... 应用GEOTRACES IDP2017[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如图4 所示.如表1 和图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放[103 ] 和洋流输运[21 ,104 ] 等因素有关. ...
... ,
99 ~
102 ])
The existing data of dissolved copper in the world surface ocean (0~50 m) (data from references [23, 83,84, 99~102]) Fig.4 ![]()
表1 不同海区、不同深度的海水溶解态铜浓度 ...
... Dissolved copper concentration in different oceans and depth
Table 1 水体类型 区域 铜浓度/(nmol/kg) n 参考文献 表层水体:开阔大洋 北太平洋 1.72±0.82 16 [99 ] 北大西洋 0.88±0.28 20 [99 ] 印度洋 1.46±0.64 10 [99 ] 南太平洋 0.53±0.26 135 [99 ] 南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ]
海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
印度洋 1.46±0.64 10 [99 ] 南太平洋 0.53±0.26 135 [99 ] 南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
南太平洋 0.53±0.26 135 [99 ] 南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
南极底层水 1.13±0.69 697 [99 ] 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... [
99 ]
海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... 如
图5 所示,铜的垂直分布模式介于营养型元素(表层因生物利用耗竭、于深层再生)和清除型元素(有强表层水来源但在整个水柱中清除)之间,其分布受到生物利用、清除和再生等过程的共同影响
[109 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值
[8 ] .
图5 世界不同大洋的溶解态铜垂直剖面分布(数据来自参考文献[<xref ref-type="bibr" rid="R21">21</xref>,<xref ref-type="bibr" rid="R99">99</xref>,<xref ref-type="bibr" rid="R118">118</xref>]) <strong>The vertical profiles of dCu in different ocean </strong>(<strong>data from references </strong>[<strong>21</strong>,<strong>99</strong>,<strong>118</strong>]) Fig.5 ![]()
Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Distribution of dissolved and particulate metals in Antarctic sea ice
2011
Sources,solubility,and dry deposition of aerosol trace elements over the East China Sea
2010
Mixing of Asian mineral dust with anthropogenic pollutants over East Asia: A model case study of a super-duststorm in March 2010
1
2012
... 应用GEOTRACES IDP2017
[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如
图4 所示.如
表1 和
图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放
[103 ] 和洋流输运
[21 ,104 ] 等因素有关.
图4 溶解态铜在世界大洋表层的现有数据分布(<strong>0~50 m</strong>)(数据来自参考文献[<xref ref-type="bibr" rid="R23">23</xref>, <xref ref-type="bibr" rid="R83">83</xref>,<xref ref-type="bibr" rid="R84">84</xref>, <xref ref-type="bibr" rid="R99">99</xref>~<xref ref-type="bibr" rid="R102">102</xref>]) The existing data of dissolved copper in the world surface ocean (0~50 m) (data from references [23, 83,84, 99~102]) Fig.4 ![]()
表1 不同海区、不同深度的海水溶解态铜浓度 ...
Cu speciation and cyanobacterial distribution in harbors subject to anthropogenic Cu inputs
1
1997
... 应用GEOTRACES IDP2017[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如图4 所示.如表1 和图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放[103 ] 和洋流输运[21 ,104 ] 等因素有关. ...
Seasonal variation and sources of dissolved trace metals in Maó Harbour,Minorca Island
1
2016
... 应用GEOTRACES IDP2017[99 ] 数据库和前人文献数据绘制的全球海洋的表层海水dCu浓度分布如图4 所示.如表1 和图4 所示,溶解态铜在表层海水中的浓度普遍在0.5~5.0 nmol/kg范围内.全球分布上表现为,近岸dCu浓度远高于开阔大洋,在渤海湾等边缘海可达10~20 nmol/kg,而开阔大洋表层海水的dCu浓度普遍低于1 nmol/kg.dCu在近岸的浓度较高,这与径流和气溶胶等陆源输入、沿海区域的人类物质排放[103 ] 和洋流输运[21 ,104 ] 等因素有关. ...
Dissolved trace metal distributions and Cu speciation in the southern Bohai Sea,China
2
2015
... Dissolved copper concentration in different oceans and depth
Table 1 水体类型 区域 铜浓度/(nmol/kg) n 参考文献 表层水体:开阔大洋 北太平洋 1.72±0.82 16 [99 ] 北大西洋 0.88±0.28 20 [99 ] 印度洋 1.46±0.64 10 [99 ] 南太平洋 0.53±0.26 135 [99 ] 南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ]
海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
Three-dimensional modeling of Cu and Pb distributions in Boston Harbor,Massachusetts and Cape Cod Bays
2
2010
... Dissolved copper concentration in different oceans and depth
Table 1 水体类型 区域 铜浓度/(nmol/kg) n 参考文献 表层水体:开阔大洋 北太平洋 1.72±0.82 16 [99 ] 北大西洋 0.88±0.28 20 [99 ] 印度洋 1.46±0.64 10 [99 ] 南太平洋 0.53±0.26 135 [99 ] 南大西洋 0.82±0.34 27 [99 ] 表层水体:边缘海 黄渤海 14.04±8.04 25 [105 ] 地中海 2.06±1.29 11 [83 ] 墨西哥湾 5.53±1.11 6 [106 ] 深层水体 北大西洋深层水 1.94±0.43 163 [99 ] 北太平洋深层水 3.15±0.60 113 [99 ] 印度洋深层水 3.60±1.03 53 [99 ] 南太平洋 3.54±0.67 402 [99 ] 南大西洋 2.43±0.55 121 [99 ] 南极底层水 1.13±0.69 697 [99 ]
海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
... 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
Winter and summer distributions of Copper,Zinc and Nickel along the International GEOTRACES Section GIPY05: Insights into deep winter mixing
1
2019
... 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
Evaluating the impact of atmospheric deposition on dissolved trace-metals in the Gulf of Aqaba,Red Sea
1
2011
... 海盆尺度铜的浓度差异,如表1 所示.dCu浓度在北太平洋表层水最高,为(1.72±0.82) nmol/kg(n =16),随后印度洋表层水为(1.46±0.64) nmol/kg(n =10),北大西洋为(0.88±0.28) nmol/kg(n =20),均高于南太平洋的(0.53±0.26) nmol/kg(n =135)和南大西洋的(0.82±0.34) nmol/kg(n =27).北半球表层海水的dCu浓度普遍高于南半球,可能是因为南半球的人类活动较少,风化作用较弱,同时接收河流输送也较少.在北半球,除了河流输送,造成各海区表层水dCu浓度差异的原因,除了河流输送外,可能主要是接收大气沉降来源的差异.铜在人类活动影响气溶胶中的溶解度大于在矿物气溶胶中的溶解度,例如亚洲尘埃事件时期沉降在东海的气溶胶中有高达50%的可溶解态铜[105 ] ,而撒哈拉沙漠气溶胶中沉降在马尾藻海和百慕大海域的铜只有1%~7%的溶解度[86 ] .北太平洋表层水dCu浓度最高,这可能是受到东亚地区受人类活动影响程度较高的气溶胶铜输送的影响[106 ] .阿拉伯海和孟加拉湾受陆地工业污染排放和撒哈拉沙漠沙尘的影响,dCu浓度也相对较高.铜在海水中的表层分布还受到浮游生物活动、混合层深度和上升流等海洋过程季节性波动的影响.例如:浮游植物的吸收作用和混合层深度的变化会导致南大洋冬季表层dCu浓度较高[107 ] ;层化以及撒哈拉沙漠的沙尘沉降[108 ] 会导致红海亚喀巴湾表层海水铜的富集. ...
Controls of trace metals in seawater
1
2003
... 如图5 所示,铜的垂直分布模式介于营养型元素(表层因生物利用耗竭、于深层再生)和清除型元素(有强表层水来源但在整个水柱中清除)之间,其分布受到生物利用、清除和再生等过程的共同影响[109 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值[8 ] . ...
The distribution of dissolved copper in the eastern Atlantic Ocean
1
1978
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Cd,Zn,Ni and Cu in the Indian Ocean
1
1992
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Seasonal metal remobilization in the sediments of Great Bay,New Hampshire
1
1984
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Copper distribution and speciation across the International GEOTRACES Section GA03
1
2015
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Impact of hydrothermalism on the ocean iron cycle
1
2016
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
A modeling assessment of the role of reversible scavenging in controlling oceanic dissolved Cu and Zn distributions
1
2013
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Biological uptake and reversible scavenging of zinc in the global ocean
1
2018
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Distribution of thorium isotopes between dissolved and particulate forms in the deep sea
1
1982
... Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
Distribution of copper-complexing ligands in Canadian Arctic waters as determined by immobilized copper(II)-ion affinity chromatography
1
2019
... 如
图5 所示,铜的垂直分布模式介于营养型元素(表层因生物利用耗竭、于深层再生)和清除型元素(有强表层水来源但在整个水柱中清除)之间,其分布受到生物利用、清除和再生等过程的共同影响
[109 ] .营养盐类型分布的铜剖面主要出现在开阔大洋中,铜在表层水体被消耗或清除而在深层水体再生;清除型分布主要出现在接收大气沉降较多的水体,铜在表层水体中体现出高值
[8 ] .
图5 世界不同大洋的溶解态铜垂直剖面分布(数据来自参考文献[<xref ref-type="bibr" rid="R21">21</xref>,<xref ref-type="bibr" rid="R99">99</xref>,<xref ref-type="bibr" rid="R118">118</xref>]) <strong>The vertical profiles of dCu in different ocean </strong>(<strong>data from references </strong>[<strong>21</strong>,<strong>99</strong>,<strong>118</strong>]) Fig.5 ![]()
Boyle等[5 ] 在1997年首次发表了GEOSECS项目中部分太平洋站位的dCu垂直剖面,结果显示dCu浓度从1 000 m处到深层水体增加地非常缓慢,这表明太平洋的中深层海水可能只发生了轻微的再矿化作用以补充水体中的铜.而非洲西北沿岸上升流地区的深层水中出现了dCu浓度升高现象[110 ] ,在高生产力的阿拉伯海也观察到显著的底层水dCu最大值[111 ] .前人对中深层海水dCu浓度分布差异性做出了不同的解释:一是沉积物或热液来源的铜自下而上补充到中深层水[112 ] ;二是高颗粒物通量自上而下输入铜[113 ] .但热液输入具有明显的区域性[114 ] ,沉积物类型和颗粒物通量也存在海区差异,这些因素都不能单独控制铜的垂直分布.Little等[115 ] 提出了铜在颗粒物上存在可逆清除的假设,即表层海水中浓度较高的颗粒物会导致相对较强的对铜的清除作用,随着水深增加,颗粒物浓度降低会进一步促进颗粒相铜的释放[116 ] .可逆清除模型在不考虑边界(海底)、侧向传输和垂向物质输送的情况下能为铜的垂直线性剖面提供较为合理的理论解释[117 ] .总而言之,颗粒物的外部来源、内部来源和再生清除等相对作用,以及铜与配体的有机络合作用共同控制着铜的垂直剖面分布. ...
What controls dissolved iron concentrations in the world ocean?
1
1997
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
Sources and fluxes of atmospheric trace elements to the Gulf of Aqaba,Red Sea
1
2008
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
Marine scavenging: Trace metal adsorption by interfacial sediment from MANOP Site H
1
1984
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
Scavenging residence times of trace metals and surface chemistry of sinking particles in the deep ocean
1
1981
... 在图5 中,太平洋dCu垂直剖面的浓度范围为1.66~3.38 nmol/kg,印度洋的dCu垂直浓度范围为1~5.49 nmol/kg,大西洋的dCu垂直浓度范围为0.89~2.52 nmol/kg,南大洋的dCu垂直浓度范围为1.01~3.05 nmol/kg.结合表1 ,可以看出,北太平洋深层水和印度洋深层水中的dCu浓度高于北大西洋和南大西洋,这是因为随着温盐环流路径的前进,水团年龄增大,再生积累的dCu越来越多[119 ] .铜在海洋中的停留时间因深度和供应来源不同而存在差异.如果只基于河流供应估算,铜在表层海洋的停留时间为2~50年[6 ] ,在深层海洋的停留时间为5 000~5 400年[5 ,23 ] .只基于大气沉降供应估算,铜在亚喀巴海湾表层50 m海水的停留时间有32年[120 ] .而基于沉积物供应的估算表明,铜的停留时间相对于清除作用较短,有650年左右[121 ,122 ] .在大洋铜源汇格局尚未明确的情况下,对表层和深层海水铜停留时间的估算仍需要进一步对大洋铜生物地球化学进行研究. ...
海洋环境中铁的来源、微生物作用过程及生态效应
1
2019
... (1)海洋中铜的生理功能.例如:海水中铜配体、铜价态对铜生物有效性的影响,浮游植物和微生物对铜的加工、吸收和转化作用,浮游生物的利用对铜同位素的分馏效应等方面.应用现代组学技术(宏基因组、宏转录组、宏蛋白组和宏代谢组)和单细胞测序技术深入研究海洋微生物对铜的代谢过程与分子机制[123 ] . ...
海洋环境中铁的来源、微生物作用过程及生态效应
1
2019
... (1)海洋中铜的生理功能.例如:海水中铜配体、铜价态对铜生物有效性的影响,浮游植物和微生物对铜的加工、吸收和转化作用,浮游生物的利用对铜同位素的分馏效应等方面.应用现代组学技术(宏基因组、宏转录组、宏蛋白组和宏代谢组)和单细胞测序技术深入研究海洋微生物对铜的代谢过程与分子机制[123 ] . ...
全球业务化海洋预报进展与展望
1
2018
... (5)利用铜数据库发展海洋模式及全球耦合预报.加强卫星、浮标、潜标等对铜的立体观测系统建设,将海水铜的生物地球化学行为纳入全球海洋预报系统,提高痕量金属的海洋模式分辨率,优化海洋物理过程参数化方案,最终实现全球海洋—大气耦合预报[124 ] . ...
全球业务化海洋预报进展与展望
1
2018
... (5)利用铜数据库发展海洋模式及全球耦合预报.加强卫星、浮标、潜标等对铜的立体观测系统建设,将海水铜的生物地球化学行为纳入全球海洋预报系统,提高痕量金属的海洋模式分辨率,优化海洋物理过程参数化方案,最终实现全球海洋—大气耦合预报[124 ] . ...




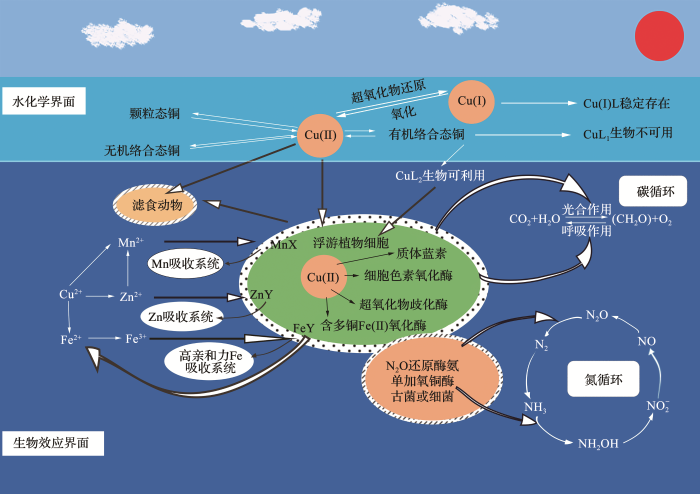
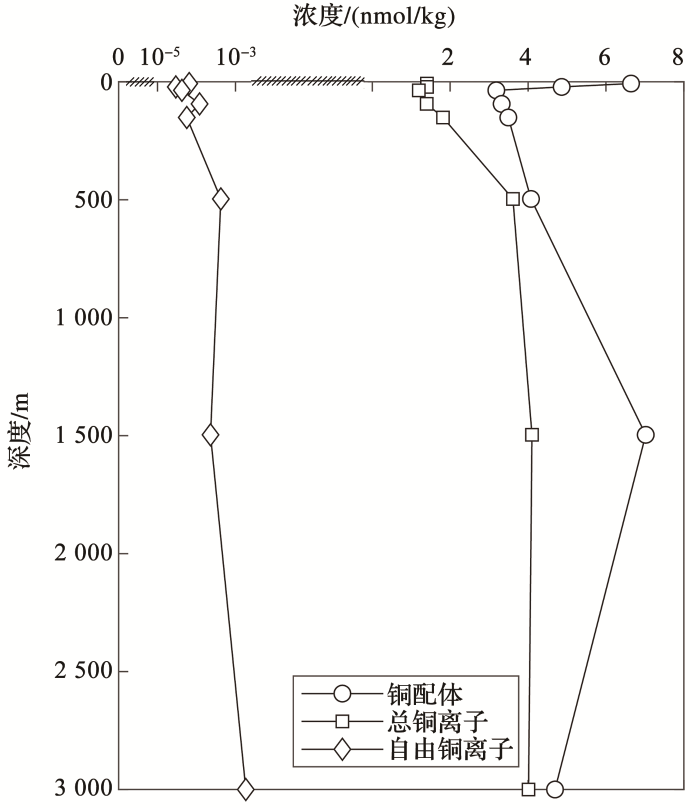
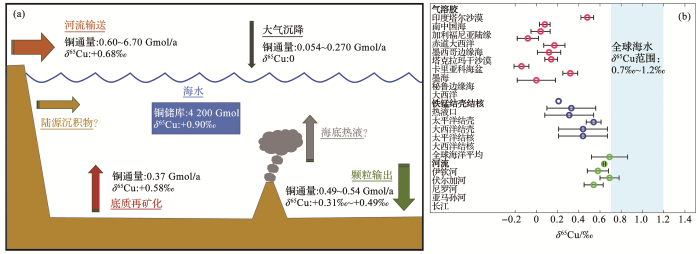
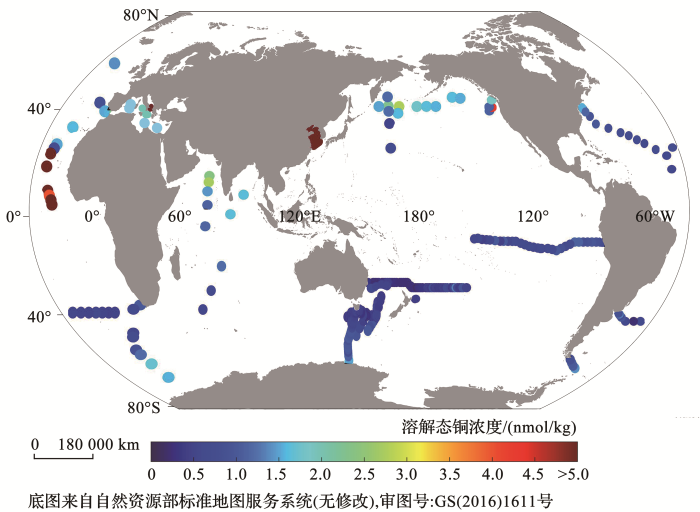
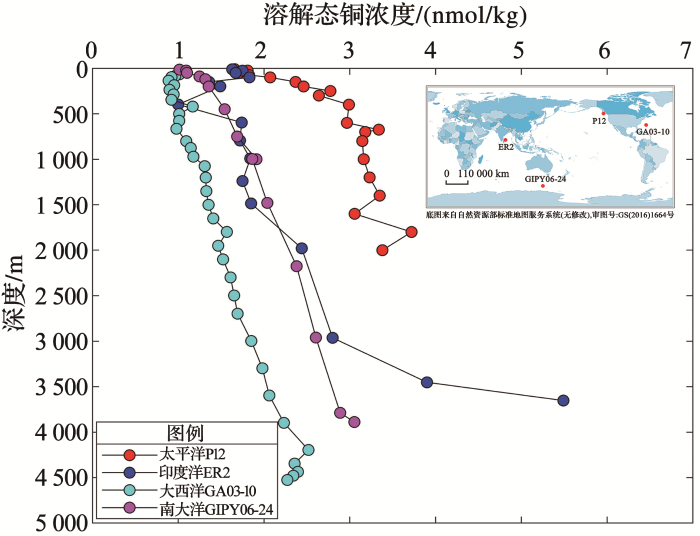

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}