Microbialites: Organosedimentary deposits of benthic microbial communities
1
1987
... 微生物岩是一种由底栖微生物群落捕获和黏结沉积物或原地形成矿物沉淀的有机沉积物[1 ] ,这种由微生物及其环境之间相互作用的石化产物占据了地球生命历史80%以上的化石记录.作为一种特征性的纹层状的微生物岩[2 ] ,叠层石已经从35亿年[3 ] 追溯到了37亿年[4 ] .Kalkowsky[2 ] 是第一个提出叠层石概念的学者,经过一个多世纪的探索,关于叠层石的解释有以下观点:① [2 ] 表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
Oolith und Stromatolith im norddeutschen Buntsandstein
3
1908
... 微生物岩是一种由底栖微生物群落捕获和黏结沉积物或原地形成矿物沉淀的有机沉积物[1 ] ,这种由微生物及其环境之间相互作用的石化产物占据了地球生命历史80%以上的化石记录.作为一种特征性的纹层状的微生物岩[2 ] ,叠层石已经从35亿年[3 ] 追溯到了37亿年[4 ] .Kalkowsky[2 ] 是第一个提出叠层石概念的学者,经过一个多世纪的探索,关于叠层石的解释有以下观点:① [2 ] 表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
... [2 ]是第一个提出叠层石概念的学者,经过一个多世纪的探索,关于叠层石的解释有以下观点:① [2 ] 表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
... [2 ]表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
Stromatolite reef from the Early Archaean Era of Australia
1
2006
... 微生物岩是一种由底栖微生物群落捕获和黏结沉积物或原地形成矿物沉淀的有机沉积物[1 ] ,这种由微生物及其环境之间相互作用的石化产物占据了地球生命历史80%以上的化石记录.作为一种特征性的纹层状的微生物岩[2 ] ,叠层石已经从35亿年[3 ] 追溯到了37亿年[4 ] .Kalkowsky[2 ] 是第一个提出叠层石概念的学者,经过一个多世纪的探索,关于叠层石的解释有以下观点:① [2 ] 表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures
1
2016
... 微生物岩是一种由底栖微生物群落捕获和黏结沉积物或原地形成矿物沉淀的有机沉积物[1 ] ,这种由微生物及其环境之间相互作用的石化产物占据了地球生命历史80%以上的化石记录.作为一种特征性的纹层状的微生物岩[2 ] ,叠层石已经从35亿年[3 ] 追溯到了37亿年[4 ] .Kalkowsky[2 ] 是第一个提出叠层石概念的学者,经过一个多世纪的探索,关于叠层石的解释有以下观点:① [2 ] 表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
Stromatolite morphogenesis: Progress and problems
1
1979
... 微生物岩是一种由底栖微生物群落捕获和黏结沉积物或原地形成矿物沉淀的有机沉积物[1 ] ,这种由微生物及其环境之间相互作用的石化产物占据了地球生命历史80%以上的化石记录.作为一种特征性的纹层状的微生物岩[2 ] ,叠层石已经从35亿年[3 ] 追溯到了37亿年[4 ] .Kalkowsky[2 ] 是第一个提出叠层石概念的学者,经过一个多世纪的探索,关于叠层石的解释有以下观点:① [2 ] 表示叠层石是微生物的生长和新陈代谢作用的产物,同时也是沉积物捕获、黏结和沉淀产生的有机沉积构造.② [5 ] 认为叠层石是“以一个受限制的界面为起点逐渐向上沉积,最终形成黏结的、石化的纹层状构造”.尽管这是一种涉及到生物、物理化学相互作用的过程,但可以达成共识的是,现代叠层石的形成主要依赖于微生物作用过程,所以现代叠层石是微生物岩的典型代表. ...
Discovery of large conical stromatolites in Lake Untersee, Antarctica
1
2011
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
Morphological record of oxygenic photosynthesis in conical stromatolites
1
2009
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
Formation and stability of oxygen‐rich bubbles that shape photosynthetic mats
1
2010
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
Biophysical basis for the geometry of conical stromatolites
1
2010
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
Influence of gas production and filament orientation on stromatolite microfabric
1
2012
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
Composition diversity of modern stromatolites: A key and window for further understanding of the formation of ancient stromatolites
1
2016
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
现代叠层石的多样化构成: 认识古代叠层石形成的关键和窗口
1
2016
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
Bahamas: Environmental setting, accretion mechanisms and role of euendoliths
7
2011
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... 在开放的海洋环境下,巴哈马Little Darby群岛的碳酸盐叠层石分布在由水流搬运碳酸盐砂颗粒的潮汐环境中;这些叠层石由丝状蓝细菌产生的胞外聚合物质(Extracellular Polymeric Substance)分泌物捕捉和黏结的鲕粒砂颗粒组成.Reid等[12 ] 在巴哈马地区进行了长期持续性实验,并取得了一些重要进展,涵盖了叠层石的形成机理、微生物席的石化、微生物新陈代谢作用和微生物群落的分布等多个方面[17 ] . ...
... Little Darby的叠层石位于潮下带地区,有些独立生长在充满碳酸盐砂颗粒的环境中,另一些则合并为大型生物礁.这些叠层石的形成机理包含2个方面:
① 位于叠层石顶部的裂须菌(
Schizothrix )生物席(
图1 a)捕获和黏结沉积物,并且随着叠层石的生长不断向上迁移,也有可能是对营养或化学成分的局限扩散的主动反应,从而形成穹窿状构造;
② 捕获或黏结由波浪和水流搬运来的碳酸盐砂颗粒,逐渐向上沉积形成毫米级别的纹层.
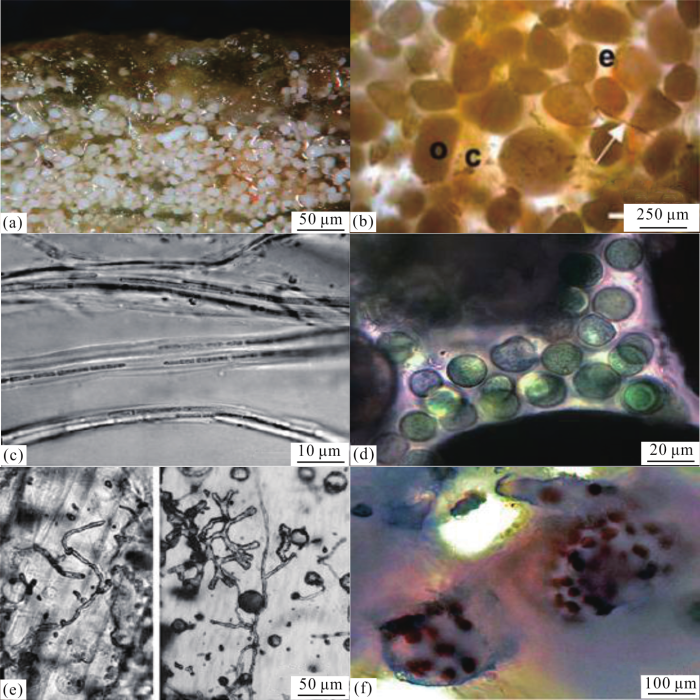
图1 巴哈马<strong> Little Darby </strong>群岛碳酸盐叠层石的微观特征<sup>[<xref ref-type="bibr" rid="R12">12</xref>]</sup> (a)位于叠层石表面的裂须菌微生物群落;(b)鲕粒砂颗粒(o),间隙充满细胞外聚合物分泌物(e)和蓝细菌(c和箭头所指);(c)主要的造席生物Schizothrix gebeleinii ,其特征是具有滑动的束状纤毛簇;(d)发育在砂颗粒间隙中的隐生菌(Aphanocapsa );(e)鲕粒砂颗粒中的微生物钻孔;(f)管状菌(Solentia ;一种真石内蓝细菌)的大规模爆发 ...
... (a)位于叠层石表面的裂须菌微生物群落;(b)鲕粒砂颗粒(o),间隙充满细胞外聚合物分泌物(e)和蓝细菌(c和箭头所指);(c)主要的造席生物
Schizothrix gebeleinii ,其特征是具有滑动的束状纤毛簇;(d)发育在砂颗粒间隙中的隐生菌(
Aphanocapsa );(e)鲕粒砂颗粒中的微生物钻孔;(f)管状菌(
Solentia ;一种真石内蓝细菌)的大规模爆发
Images showing microscopic features of carbonate stromatolites in Little Darby Cay of Bahamas<sup>[<xref ref-type="bibr" rid="R12">12</xref>]</sup> (a)Microbial community of Schizothrix on the surface of stromatolites; (b)Oolitic sand particles (o), interstitial filled with extracellular polymer secretions (e) and cyanobacteria (c,arrow); (c) Schizothrix gebeleinii , dominant microorganism in the mat, is characterized by presence of trichome bundled; (d)Aphanocapsa developing in the interstitial spaces of sand grains; (e)Microborings found in particles of oolitic sands; (f)Growth burst of the euendolithic cyanobacterium Solentia ...
... 主导叠层石的砂颗粒是由丝状蓝细菌产生的胞外聚合物质分泌物黏结而成(图1 b).其中,Schizothrix gebeleinii 作为一种典型的造席丝状蓝细菌, 它在细胞外聚合物的鞘中有滑动的束状纤毛簇(图1 c);另外的造席微生物有裂须菌(Schizothrix )、管状菌(Solentia )、螺旋菌(Spirulina )和较少的厌氧异养细菌等;裂须菌(Schizothrix )最重要的作用在于其内部的鞘被有机细菌降解,如硫酸盐还原细菌.另一种生物量产生在被黏结的鲕粒砂颗粒相互接触构成的空隙内,这个空间也适合球状蓝细菌的生长,例如隐生菌(Aphanocapsa )(图1 d).构成Little Darby群岛叠层石的第二个部分是黏结和捕捉到的鲕粒砂颗粒;这些特别的鲕粒砂颗粒具有活跃的微生物活动和光合作用,但被丝状蓝细菌产生的胞外聚合物质分泌物黏结后开始趋于稳定状态.当蓝细菌群落被融入到生物席时,砂颗粒就会被提前钻孔(图1 e),但仅有一小部分中生存着活跃的真石内菌;在砂颗粒被叠层体捕获并稳定后,真石内菌的生存环境发生了很大变化,从而促使管状菌(Solentia )大规模爆发[18 ,19 ] (图1 f),并且这个过程与生物席的石化过程基本一致[12 ,20 ] . ...
... Little Darby群岛叠层石的石化作用实际上是一个将叠层石的成因与其历史记录相联系的过程[12 ] .在叠层石的石化过程中,两个紧密耦合的组成部分在微生物席上驱使着碳酸盐的沉淀:① ②
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Cryptozoon and associate stromatolites from the Recent, Shark bay, Western Australia
3
1961
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... Logan[13 ] 在澳大利亚鲨鱼湾的Hamelin Pool中发现了第一批现代叠层石,其大小和形状都类似于寒武纪叠层石,因此,Hamelin Pool的叠层石一直是理解现代和古代叠层石体系的重要模型[21 ,22 ] .构成这些叠层石的骨架和胶结物主要依赖与微生物活动有关的微晶碳酸盐的沉淀作用[23 ] ,引发碳酸盐沉淀的机制可能与异养细菌活动(如硫酸盐还原反应)、光合作用CO2 吸收、有机质再矿化过程[24 ] 以及地下水注入等有关. ...
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Sedimentology and temporal distribution of microbial mats from Brejo do Espinho, Rio de Janeiro, Brazil
3
2012
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Microbial-mat sedimentology: A young branch from sedimentology
2
2011
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
微生物席沉积学:一个年轻的沉积学分支
2
2011
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Feature and nature of microbial-mat: Theoretical basis of microbial-mat sedimentology
2
2014
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
微生物席的特征和属性: 微生物席沉积学的理论基础
2
2014
... 现代叠层石多样性的构成,即泥质叠层石[6 ] 、硅质叠层石[7 ,8 ,9 ,10 ] 和碳酸盐叠层石[11 ] ,它们将成为探索寒武纪叠层石形成的一个锁眼,而不完全是开启寒武纪叠层石世界的一把钥匙.基于前人的研究成果,以下几个地区的碳酸盐叠层石研究已取得丰富的成果:① [12 ] ;② [13 ] ;③ [14 ] .虽然这些现代碳酸盐叠层石为了解寒武纪环境和叠层石的形成提供了一个潜在的窗口,同时为了解底栖微生物群落如何与环境相互作用提供了基础;但是,通过对本溪火连寨剖面寒武系张夏组中部的叠层石生物丘和现代碳酸盐叠层石的对比,发现两者在生长形态和沉积模式等方面有显著差异.所以,尽管这个潜在的窗口还不能完全窥探复杂多变的寒武纪叠层石世界,却为了解寒武纪叠层石的构建以及微生物碳酸盐岩工厂提供了丰富的思考途径[15 ,16 ] . ...
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
The microbial communities of the modern marine stromatolites at Highborn Cay, Bahamas
1
2009
... 在开放的海洋环境下,巴哈马Little Darby群岛的碳酸盐叠层石分布在由水流搬运碳酸盐砂颗粒的潮汐环境中;这些叠层石由丝状蓝细菌产生的胞外聚合物质(Extracellular Polymeric Substance)分泌物捕捉和黏结的鲕粒砂颗粒组成.Reid等[12 ] 在巴哈马地区进行了长期持续性实验,并取得了一些重要进展,涵盖了叠层石的形成机理、微生物席的石化、微生物新陈代谢作用和微生物群落的分布等多个方面[17 ] . ...
Detection and enumeration of microbial cells within highly porous calcareous reef sands
1
2006
... 主导叠层石的砂颗粒是由丝状蓝细菌产生的胞外聚合物质分泌物黏结而成(图1 b).其中,Schizothrix gebeleinii 作为一种典型的造席丝状蓝细菌, 它在细胞外聚合物的鞘中有滑动的束状纤毛簇(图1 c);另外的造席微生物有裂须菌(Schizothrix )、管状菌(Solentia )、螺旋菌(Spirulina )和较少的厌氧异养细菌等;裂须菌(Schizothrix )最重要的作用在于其内部的鞘被有机细菌降解,如硫酸盐还原细菌.另一种生物量产生在被黏结的鲕粒砂颗粒相互接触构成的空隙内,这个空间也适合球状蓝细菌的生长,例如隐生菌(Aphanocapsa )(图1 d).构成Little Darby群岛叠层石的第二个部分是黏结和捕捉到的鲕粒砂颗粒;这些特别的鲕粒砂颗粒具有活跃的微生物活动和光合作用,但被丝状蓝细菌产生的胞外聚合物质分泌物黏结后开始趋于稳定状态.当蓝细菌群落被融入到生物席时,砂颗粒就会被提前钻孔(图1 e),但仅有一小部分中生存着活跃的真石内菌;在砂颗粒被叠层体捕获并稳定后,真石内菌的生存环境发生了很大变化,从而促使管状菌(Solentia )大规模爆发[18 ,19 ] (图1 f),并且这个过程与生物席的石化过程基本一致[12 ,20 ] . ...
On microbiocorrosion
1
2011
... 主导叠层石的砂颗粒是由丝状蓝细菌产生的胞外聚合物质分泌物黏结而成(图1 b).其中,Schizothrix gebeleinii 作为一种典型的造席丝状蓝细菌, 它在细胞外聚合物的鞘中有滑动的束状纤毛簇(图1 c);另外的造席微生物有裂须菌(Schizothrix )、管状菌(Solentia )、螺旋菌(Spirulina )和较少的厌氧异养细菌等;裂须菌(Schizothrix )最重要的作用在于其内部的鞘被有机细菌降解,如硫酸盐还原细菌.另一种生物量产生在被黏结的鲕粒砂颗粒相互接触构成的空隙内,这个空间也适合球状蓝细菌的生长,例如隐生菌(Aphanocapsa )(图1 d).构成Little Darby群岛叠层石的第二个部分是黏结和捕捉到的鲕粒砂颗粒;这些特别的鲕粒砂颗粒具有活跃的微生物活动和光合作用,但被丝状蓝细菌产生的胞外聚合物质分泌物黏结后开始趋于稳定状态.当蓝细菌群落被融入到生物席时,砂颗粒就会被提前钻孔(图1 e),但仅有一小部分中生存着活跃的真石内菌;在砂颗粒被叠层体捕获并稳定后,真石内菌的生存环境发生了很大变化,从而促使管状菌(Solentia )大规模爆发[18 ,19 ] (图1 f),并且这个过程与生物席的石化过程基本一致[12 ,20 ] . ...
Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms
1
2000
... 主导叠层石的砂颗粒是由丝状蓝细菌产生的胞外聚合物质分泌物黏结而成(图1 b).其中,Schizothrix gebeleinii 作为一种典型的造席丝状蓝细菌, 它在细胞外聚合物的鞘中有滑动的束状纤毛簇(图1 c);另外的造席微生物有裂须菌(Schizothrix )、管状菌(Solentia )、螺旋菌(Spirulina )和较少的厌氧异养细菌等;裂须菌(Schizothrix )最重要的作用在于其内部的鞘被有机细菌降解,如硫酸盐还原细菌.另一种生物量产生在被黏结的鲕粒砂颗粒相互接触构成的空隙内,这个空间也适合球状蓝细菌的生长,例如隐生菌(Aphanocapsa )(图1 d).构成Little Darby群岛叠层石的第二个部分是黏结和捕捉到的鲕粒砂颗粒;这些特别的鲕粒砂颗粒具有活跃的微生物活动和光合作用,但被丝状蓝细菌产生的胞外聚合物质分泌物黏结后开始趋于稳定状态.当蓝细菌群落被融入到生物席时,砂颗粒就会被提前钻孔(图1 e),但仅有一小部分中生存着活跃的真石内菌;在砂颗粒被叠层体捕获并稳定后,真石内菌的生存环境发生了很大变化,从而促使管状菌(Solentia )大规模爆发[18 ,19 ] (图1 f),并且这个过程与生物席的石化过程基本一致[12 ,20 ] . ...
Modern algal stromatolites at Hamelin Pool, a hypersaline barred basin in Shark Bay, Western Australia
3
1976
... Logan[13 ] 在澳大利亚鲨鱼湾的Hamelin Pool中发现了第一批现代叠层石,其大小和形状都类似于寒武纪叠层石,因此,Hamelin Pool的叠层石一直是理解现代和古代叠层石体系的重要模型[21 ,22 ] .构成这些叠层石的骨架和胶结物主要依赖与微生物活动有关的微晶碳酸盐的沉淀作用[23 ] ,引发碳酸盐沉淀的机制可能与异养细菌活动(如硫酸盐还原反应)、光合作用CO2 吸收、有机质再矿化过程[24 ] 以及地下水注入等有关. ...
... 环境压力通常被认为是驱动宏观形态学的因素,而微观形态学特征则归因于生物作用[25 ,26 ] ;因此,在整个叠层石历史可观测的时间尺度记录中,解释沉积环境和叠层石的生物学贡献变得尤为重要[27 ] .水位变化和底层陆架地貌[23 ,26 ] 可能是导致Hamelin Pool的叠层石分带的驱动因素;潮间带上部分布着未石化的微生物席,潮间带下部和潮下带地区分布石化程度较好的微生物席,依据宏观形态可划分为脓疱状、光滑状和胶粒状微生物席[21 ] ,它们都代表着多样化的微生物群落. ...
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
Shark Bay stromatolites: Microfabrics and reinterpretation of origins
1
2003
... Logan[13 ] 在澳大利亚鲨鱼湾的Hamelin Pool中发现了第一批现代叠层石,其大小和形状都类似于寒武纪叠层石,因此,Hamelin Pool的叠层石一直是理解现代和古代叠层石体系的重要模型[21 ,22 ] .构成这些叠层石的骨架和胶结物主要依赖与微生物活动有关的微晶碳酸盐的沉淀作用[23 ] ,引发碳酸盐沉淀的机制可能与异养细菌活动(如硫酸盐还原反应)、光合作用CO2 吸收、有机质再矿化过程[24 ] 以及地下水注入等有关. ...
New multi-scale perspectives on the stromatolites of Shark Bay, Western Australia
5
2016
... Logan[13 ] 在澳大利亚鲨鱼湾的Hamelin Pool中发现了第一批现代叠层石,其大小和形状都类似于寒武纪叠层石,因此,Hamelin Pool的叠层石一直是理解现代和古代叠层石体系的重要模型[21 ,22 ] .构成这些叠层石的骨架和胶结物主要依赖与微生物活动有关的微晶碳酸盐的沉淀作用[23 ] ,引发碳酸盐沉淀的机制可能与异养细菌活动(如硫酸盐还原反应)、光合作用CO2 吸收、有机质再矿化过程[24 ] 以及地下水注入等有关. ...
... 环境压力通常被认为是驱动宏观形态学的因素,而微观形态学特征则归因于生物作用[25 ,26 ] ;因此,在整个叠层石历史可观测的时间尺度记录中,解释沉积环境和叠层石的生物学贡献变得尤为重要[27 ] .水位变化和底层陆架地貌[23 ,26 ] 可能是导致Hamelin Pool的叠层石分带的驱动因素;潮间带上部分布着未石化的微生物席,潮间带下部和潮下带地区分布石化程度较好的微生物席,依据宏观形态可划分为脓疱状、光滑状和胶粒状微生物席[21 ] ,它们都代表着多样化的微生物群落. ...
... 在潮间带上部分布着未石化的光滑状和脓疱状生物席(
图2 a,b).其中,光滑状生物席具有良好的纹层,以丝状蓝细菌为主导,如微鞘菌(
Microcoleus )(
图2 c)和裂须菌(
Schizothrix ).而脓疱状生物席无纹层结构,主要由嵌在胞外聚合物中较小的球状蓝细菌组成(
图2 d).
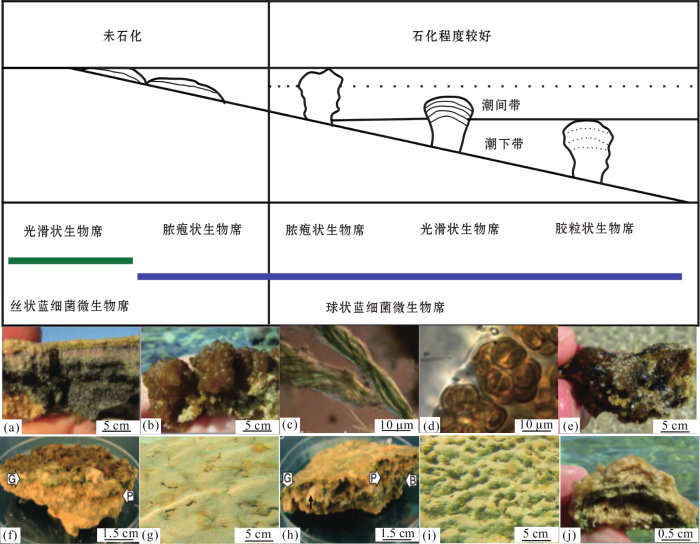
图2 <strong>Hamelin Pool</strong>的叠层石<sup>[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R28">28</xref>]</sup> (a),(b)分别为未石化的光滑状和脓胞状生物席构成的叠层石;(c)微鞘菌(Microcoleus );(d)石囊藻(Entophysalis );(e)石化的脓胞状生物席构成的叠层石表面;(f)脓疱状生物席表面;(g)石化的光滑状生物席构成的叠层石表面;(h)光滑状生物席表面,(f)和(h)中箭头G表示蓝细菌微生物群落组成的绿色层,箭头P代表紫色硫细菌组成的粉色层,箭头B代表硫酸盐还原细菌组成的黑色层;(i)石化的胶粒状生物席构成的叠层石表面;(j)叠层石的穹状构造 ...
... (a),(b)分别为未石化的光滑状和脓胞状生物席构成的叠层石;(c)微鞘菌(
Microcoleus );(d)石囊藻(
Entophysalis );(e)石化的脓胞状生物席构成的叠层石表面;(f)脓疱状生物席表面;(g)石化的光滑状生物席构成的叠层石表面;(h)光滑状生物席表面,(f)和(h)中箭头G表示蓝细菌微生物群落组成的绿色层,箭头P代表紫色硫细菌组成的粉色层,箭头B代表硫酸盐还原细菌组成的黑色层;(i)石化的胶粒状生物席构成的叠层石表面;(j)叠层石的穹状构造
Stromatolites from Hamelin Pool<sup>[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R28">28</xref>]</sup> (a),(b)They are stromatolites composed of smooth and pustular mats which are non-lithifying;(c)Microcoleus ; (d)Entophysalis ;(e)The surface of stromatolites composed of lithified pustular mats; (f) Surface of pustular mats; (g)The surface of stromatolites composed of lithified smooth mats; (h)Surface of smooth mats, arrow G represents the green layer composed of cyanobacteria microbial community, arrow P represents the pink layer composed of purple sulfur bacteria, and arrow B represents the black layer composed of sulfate reducing bacteria;(i)The surface of stromatolites composed of lithified colloform mats; (j)A domed structure of stromatolites ...
... 在上潮间带至中潮间带区域,脓胞状生物席构成了柱状叠层石;这些叠层石具有由蓝细菌组成的无清晰纹层的暗色表面层(图2 e,f),这种蓝细菌基于其形态判断为石囊藻属,而构成脓胞生物席主要的蓝细菌是双歧菌(Scytonema ).值得注意的是,脓疱状生物席内富集了大量的光合基因,说明光合作用可能是该生物席驱动沉淀的主要代谢方式.在低潮间带至浅潮下带区域,光滑状生物席构成了较小的柱状叠层石;这些叠层石具有发育较好的纹层和光滑的表面层[28 ] (图2 g),并且由典型的球状蓝细菌主导,例如隐杆菌(Aphanothece )和隐球菌(Aphanocapsa ).表层之下有一组表征蓝细菌的深粉色和暗黑色斑状物(图2 h),深粉色是由于硫化物氧化过程形成,而暗黑色是因为硫酸盐还原反应的发生,还存在一些绿色层,是由蓝细菌的产氧光合作用形成.潮下带区域的叠层石具有层状结构并且表面呈卷曲状[28 ] (图2 i),单个的穹隆状叠层石(图2 j)是由胶粒状生物席构成.综上所述,潮间带下部和潮下带地区构成叠层石的微生物席中球状蓝细菌明显富集[23 ] .其次,在脓疱状和光滑状生物席中发生着多样化的新陈代谢活动:① ② ③ [26 ] .总之,无论从哪个角度看,Hamelin Pool都是一个壮观的实验室.在微生物群落多样性和叠层石生长模式等方面,Hamelin Pool的现代叠层石为了解寒武纪环境提供了一个潜在的窗口,为重建古环境和了解底栖微生物群落如何与这些环境相互作用奠定了基础. ...
Processes of carbonate precipitation in modern microbial mats
1
2009
... Logan[13 ] 在澳大利亚鲨鱼湾的Hamelin Pool中发现了第一批现代叠层石,其大小和形状都类似于寒武纪叠层石,因此,Hamelin Pool的叠层石一直是理解现代和古代叠层石体系的重要模型[21 ,22 ] .构成这些叠层石的骨架和胶结物主要依赖与微生物活动有关的微晶碳酸盐的沉淀作用[23 ] ,引发碳酸盐沉淀的机制可能与异养细菌活动(如硫酸盐还原反应)、光合作用CO2 吸收、有机质再矿化过程[24 ] 以及地下水注入等有关. ...
Living dendrolitic microbial mats in Hamelin Pool, Shark Bay, Western Australia
2
2018
... 环境压力通常被认为是驱动宏观形态学的因素,而微观形态学特征则归因于生物作用[25 ,26 ] ;因此,在整个叠层石历史可观测的时间尺度记录中,解释沉积环境和叠层石的生物学贡献变得尤为重要[27 ] .水位变化和底层陆架地貌[23 ,26 ] 可能是导致Hamelin Pool的叠层石分带的驱动因素;潮间带上部分布着未石化的微生物席,潮间带下部和潮下带地区分布石化程度较好的微生物席,依据宏观形态可划分为脓疱状、光滑状和胶粒状微生物席[21 ] ,它们都代表着多样化的微生物群落. ...
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
Stromatolite provinces of Hamelin Pool: Physiographic controls on stromatolites and associated lithofacies
4
2019
... 环境压力通常被认为是驱动宏观形态学的因素,而微观形态学特征则归因于生物作用[25 ,26 ] ;因此,在整个叠层石历史可观测的时间尺度记录中,解释沉积环境和叠层石的生物学贡献变得尤为重要[27 ] .水位变化和底层陆架地貌[23 ,26 ] 可能是导致Hamelin Pool的叠层石分带的驱动因素;潮间带上部分布着未石化的微生物席,潮间带下部和潮下带地区分布石化程度较好的微生物席,依据宏观形态可划分为脓疱状、光滑状和胶粒状微生物席[21 ] ,它们都代表着多样化的微生物群落. ...
... ,26 ]可能是导致Hamelin Pool的叠层石分带的驱动因素;潮间带上部分布着未石化的微生物席,潮间带下部和潮下带地区分布石化程度较好的微生物席,依据宏观形态可划分为脓疱状、光滑状和胶粒状微生物席[21 ] ,它们都代表着多样化的微生物群落. ...
... 在上潮间带至中潮间带区域,脓胞状生物席构成了柱状叠层石;这些叠层石具有由蓝细菌组成的无清晰纹层的暗色表面层(图2 e,f),这种蓝细菌基于其形态判断为石囊藻属,而构成脓胞生物席主要的蓝细菌是双歧菌(Scytonema ).值得注意的是,脓疱状生物席内富集了大量的光合基因,说明光合作用可能是该生物席驱动沉淀的主要代谢方式.在低潮间带至浅潮下带区域,光滑状生物席构成了较小的柱状叠层石;这些叠层石具有发育较好的纹层和光滑的表面层[28 ] (图2 g),并且由典型的球状蓝细菌主导,例如隐杆菌(Aphanothece )和隐球菌(Aphanocapsa ).表层之下有一组表征蓝细菌的深粉色和暗黑色斑状物(图2 h),深粉色是由于硫化物氧化过程形成,而暗黑色是因为硫酸盐还原反应的发生,还存在一些绿色层,是由蓝细菌的产氧光合作用形成.潮下带区域的叠层石具有层状结构并且表面呈卷曲状[28 ] (图2 i),单个的穹隆状叠层石(图2 j)是由胶粒状生物席构成.综上所述,潮间带下部和潮下带地区构成叠层石的微生物席中球状蓝细菌明显富集[23 ] .其次,在脓疱状和光滑状生物席中发生着多样化的新陈代谢活动:① ② ③ [26 ] .总之,无论从哪个角度看,Hamelin Pool都是一个壮观的实验室.在微生物群落多样性和叠层石生长模式等方面,Hamelin Pool的现代叠层石为了解寒武纪环境提供了一个潜在的窗口,为重建古环境和了解底栖微生物群落如何与这些环境相互作用奠定了基础. ...
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
The vulnerability of microbial ecosystems in a changing climate: Potential impact in Shark Bay
1
2019
... 环境压力通常被认为是驱动宏观形态学的因素,而微观形态学特征则归因于生物作用[25 ,26 ] ;因此,在整个叠层石历史可观测的时间尺度记录中,解释沉积环境和叠层石的生物学贡献变得尤为重要[27 ] .水位变化和底层陆架地貌[23 ,26 ] 可能是导致Hamelin Pool的叠层石分带的驱动因素;潮间带上部分布着未石化的微生物席,潮间带下部和潮下带地区分布石化程度较好的微生物席,依据宏观形态可划分为脓疱状、光滑状和胶粒状微生物席[21 ] ,它们都代表着多样化的微生物群落. ...
Comparative metagenomics provides insight into the ecosystem functioning of the Shark Bay stromatolites, Western Australia
4
2018
... 在潮间带上部分布着未石化的光滑状和脓疱状生物席(
图2 a,b).其中,光滑状生物席具有良好的纹层,以丝状蓝细菌为主导,如微鞘菌(
Microcoleus )(
图2 c)和裂须菌(
Schizothrix ).而脓疱状生物席无纹层结构,主要由嵌在胞外聚合物中较小的球状蓝细菌组成(
图2 d).
图2 <strong>Hamelin Pool</strong>的叠层石<sup>[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R28">28</xref>]</sup> (a),(b)分别为未石化的光滑状和脓胞状生物席构成的叠层石;(c)微鞘菌(Microcoleus );(d)石囊藻(Entophysalis );(e)石化的脓胞状生物席构成的叠层石表面;(f)脓疱状生物席表面;(g)石化的光滑状生物席构成的叠层石表面;(h)光滑状生物席表面,(f)和(h)中箭头G表示蓝细菌微生物群落组成的绿色层,箭头P代表紫色硫细菌组成的粉色层,箭头B代表硫酸盐还原细菌组成的黑色层;(i)石化的胶粒状生物席构成的叠层石表面;(j)叠层石的穹状构造 ...
... (a),(b)分别为未石化的光滑状和脓胞状生物席构成的叠层石;(c)微鞘菌(
Microcoleus );(d)石囊藻(
Entophysalis );(e)石化的脓胞状生物席构成的叠层石表面;(f)脓疱状生物席表面;(g)石化的光滑状生物席构成的叠层石表面;(h)光滑状生物席表面,(f)和(h)中箭头G表示蓝细菌微生物群落组成的绿色层,箭头P代表紫色硫细菌组成的粉色层,箭头B代表硫酸盐还原细菌组成的黑色层;(i)石化的胶粒状生物席构成的叠层石表面;(j)叠层石的穹状构造
Stromatolites from Hamelin Pool<sup>[<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R28">28</xref>]</sup> (a),(b)They are stromatolites composed of smooth and pustular mats which are non-lithifying;(c)Microcoleus ; (d)Entophysalis ;(e)The surface of stromatolites composed of lithified pustular mats; (f) Surface of pustular mats; (g)The surface of stromatolites composed of lithified smooth mats; (h)Surface of smooth mats, arrow G represents the green layer composed of cyanobacteria microbial community, arrow P represents the pink layer composed of purple sulfur bacteria, and arrow B represents the black layer composed of sulfate reducing bacteria;(i)The surface of stromatolites composed of lithified colloform mats; (j)A domed structure of stromatolites ...
... 在上潮间带至中潮间带区域,脓胞状生物席构成了柱状叠层石;这些叠层石具有由蓝细菌组成的无清晰纹层的暗色表面层(图2 e,f),这种蓝细菌基于其形态判断为石囊藻属,而构成脓胞生物席主要的蓝细菌是双歧菌(Scytonema ).值得注意的是,脓疱状生物席内富集了大量的光合基因,说明光合作用可能是该生物席驱动沉淀的主要代谢方式.在低潮间带至浅潮下带区域,光滑状生物席构成了较小的柱状叠层石;这些叠层石具有发育较好的纹层和光滑的表面层[28 ] (图2 g),并且由典型的球状蓝细菌主导,例如隐杆菌(Aphanothece )和隐球菌(Aphanocapsa ).表层之下有一组表征蓝细菌的深粉色和暗黑色斑状物(图2 h),深粉色是由于硫化物氧化过程形成,而暗黑色是因为硫酸盐还原反应的发生,还存在一些绿色层,是由蓝细菌的产氧光合作用形成.潮下带区域的叠层石具有层状结构并且表面呈卷曲状[28 ] (图2 i),单个的穹隆状叠层石(图2 j)是由胶粒状生物席构成.综上所述,潮间带下部和潮下带地区构成叠层石的微生物席中球状蓝细菌明显富集[23 ] .其次,在脓疱状和光滑状生物席中发生着多样化的新陈代谢活动:① ② ③ [26 ] .总之,无论从哪个角度看,Hamelin Pool都是一个壮观的实验室.在微生物群落多样性和叠层石生长模式等方面,Hamelin Pool的现代叠层石为了解寒武纪环境提供了一个潜在的窗口,为重建古环境和了解底栖微生物群落如何与这些环境相互作用奠定了基础. ...
... [28 ](图2 i),单个的穹隆状叠层石(图2 j)是由胶粒状生物席构成.综上所述,潮间带下部和潮下带地区构成叠层石的微生物席中球状蓝细菌明显富集[23 ] .其次,在脓疱状和光滑状生物席中发生着多样化的新陈代谢活动:① ② ③ [26 ] .总之,无论从哪个角度看,Hamelin Pool都是一个壮观的实验室.在微生物群落多样性和叠层石生长模式等方面,Hamelin Pool的现代叠层石为了解寒武纪环境提供了一个潜在的窗口,为重建古环境和了解底栖微生物群落如何与这些环境相互作用奠定了基础. ...
Lithifying microbial mats in Lagoa Vermelha, Brazil: Modern precambrian relics?
5
2006
... 通过对巴西东南部Lagoa Vermelha潟湖的微生物群落和矿物形成过程的综合分析,从而更详细地解释了高镁方解石和白云石形成的化学条件[29 ] .Lagoa Vermelha的叠层石只存在于极端的温度、盐度和营养物质条件下或水动力有利条件下矿物饱和的环境中,因为这种环境限制了生物体的发展,同时也增强了对微生物席的保护[29 ] . ...
... [29 ]. ...
... Lagoa Vermelha高盐潟湖的特点是较低的沉淀/蒸发比以及丰富的碳酸盐沉淀
[30 ] ;
图3 a所示的生物席约3 cm厚,由碳酸盐构成的白色纹层和非石化的有机质层相互交替组成.非石化的有机质层从上至下显示出绿色、棕色和红色的微生物群落分层;顶部的绿色层由嗜盐的蓝细菌如螺旋菌(
Spirulina )、黏球菌(
Gloeocapsa )和微鞘菌(
Microcoleus )组成;位于绿色层之下的棕色层内栖息着部分丝状蓝细菌,如螺旋菌和微鞘菌;大量的硫化物氧化细菌在蓝细菌底层处被发现,由储存在细胞内的硫元素所识别
[31 ] (
图3 b).因此,Lagoa Vermelha的微生物群落自上而下分层为绿色的光合细菌层,其次是寄居异养细菌的棕色层,紧接着是含有紫色硫细菌的红色层,以及带有硫酸盐还原菌的底部灰色层(
图3 a).
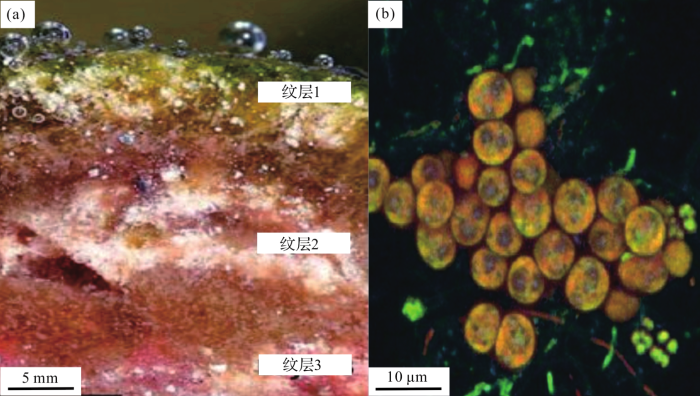
图3 巴西<strong> Lagoa Vermelha </strong>现代碳酸盐叠层石的总体特征<sup>[<xref ref-type="bibr" rid="R29">29</xref>]</sup> (a)微生物席形成的叠层石的横切面,表现为已石化的碳酸盐沉淀物构成的白色纹层和未石化的有机质层的交替,纹层1为蓝细菌主导的绿色层,纹层2为异养细菌主导的棕色层,纹层3为紫色硫细菌主导的红色层;(b)由较底层的细菌组成的样本,这一层显示出微红色,是紫色硫细菌的象征 ...
... (a)微生物席形成的叠层石的横切面,表现为已石化的碳酸盐沉淀物构成的白色纹层和未石化的有机质层的交替,纹层1为蓝细菌主导的绿色层,纹层2为异养细菌主导的棕色层,纹层3为紫色硫细菌主导的红色层;(b)由较底层的细菌组成的样本,这一层显示出微红色,是紫色硫细菌的象征
Images showing general features of modern carbonate stromatolites in Lagoa Vermelha, Brazil<sup>[<xref ref-type="bibr" rid="R29">29</xref>]</sup> (a)The cross section of stromatolites formed by the microbial mat shows alternating layers of white laminae composed of fossilized carbonate sediments with layers of unfossilized organic matter. Layer 1 is the green layer dominated by cyanobacteria, layer 2 is the brown layer dominated by heterotrophic bacteria, and layer 3 is the red layer dominated by purple sulfur bacteria; (b) A sample of bacteria from the lower layer, this layer shows a reddish tint, the symbol of purple sulfur bacteria ...
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
Hypersalinity: Global distribution, causes, and present and future effects on the Biota of Estuaries and Lagoons
2
2019
... Lagoa Vermelha高盐潟湖的特点是较低的沉淀/蒸发比以及丰富的碳酸盐沉淀[30 ] ;图3 a所示的生物席约3 cm厚,由碳酸盐构成的白色纹层和非石化的有机质层相互交替组成.非石化的有机质层从上至下显示出绿色、棕色和红色的微生物群落分层;顶部的绿色层由嗜盐的蓝细菌如螺旋菌(Spirulina )、黏球菌(Gloeocapsa )和微鞘菌(Microcoleus )组成;位于绿色层之下的棕色层内栖息着部分丝状蓝细菌,如螺旋菌和微鞘菌;大量的硫化物氧化细菌在蓝细菌底层处被发现,由储存在细胞内的硫元素所识别[31 ] (图3 b).因此,Lagoa Vermelha的微生物群落自上而下分层为绿色的光合细菌层,其次是寄居异养细菌的棕色层,紧接着是含有紫色硫细菌的红色层,以及带有硫酸盐还原菌的底部灰色层(图3 a). ...
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
Western Australia
1
1990
... Lagoa Vermelha高盐潟湖的特点是较低的沉淀/蒸发比以及丰富的碳酸盐沉淀[30 ] ;图3 a所示的生物席约3 cm厚,由碳酸盐构成的白色纹层和非石化的有机质层相互交替组成.非石化的有机质层从上至下显示出绿色、棕色和红色的微生物群落分层;顶部的绿色层由嗜盐的蓝细菌如螺旋菌(Spirulina )、黏球菌(Gloeocapsa )和微鞘菌(Microcoleus )组成;位于绿色层之下的棕色层内栖息着部分丝状蓝细菌,如螺旋菌和微鞘菌;大量的硫化物氧化细菌在蓝细菌底层处被发现,由储存在细胞内的硫元素所识别[31 ] (图3 b).因此,Lagoa Vermelha的微生物群落自上而下分层为绿色的光合细菌层,其次是寄居异养细菌的棕色层,紧接着是含有紫色硫细菌的红色层,以及带有硫酸盐还原菌的底部灰色层(图3 a). ...
Diatoms from the most hypersaline lagoon in Brazil: Vermelha lagoon
2
2019
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
... [32 ,33 ].随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
Role of sulfide oxidation in dolomitization: Sediment and pore-water geochemistry of a modern hypersaline lagoon system
1
2004
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
Formation of stromatolite lamina at the interface of oxygenic-anoxygenic photosynthesis
2
2018
... Lagoa Vermelha高盐环境中强烈的微生物活性是形成白云石等自生矿物的必要化学条件[29 ,32 ] ,产生白云石的过程发生在沉积物中含氧—无氧界面附近.在沉积物的缺氧区域,硫酸盐还原反应和盐度控制着白云石的沉淀[14 ,21 ,30 ] ;在含氧—无氧边界处,硫化物氧化以及由此导致的pH值下降则有利于白云石的产生[32 ,33 ] .随着生物席向上生长,碳酸盐薄层逐渐叠加并沉淀在含氧—无氧界面上,这表明含氧和无氧光合作用产物的垂直结构可能促成了叠层石中纹层的形成[34 ] . ...
... 微生物席上紧密相连的碳酸盐岩纹层可类比于叠层石的纹层构造,并且没有发现明显的被捕获的碳酸盐颗粒存在,所以碳酸盐的沉淀可能导致了叠层石纹层的形成.主要通过微生物的新陈代谢过程促使Lagoa Vermelha生物席中的碳酸盐析出,例如呼吸作用促使方解石在生物席表面层析出,而硫化物氧化和硫酸盐还原反应诱发了成岩过程,促进了高镁方解石和钙白云石的产生(图3 a纹层1).如图3 a叠层石纹层所示,微生物席系统的典型特征是从表层到深层逐渐多样化的生物活动[34 ,35 ] ,分别为:① ② ③ ④
The role of purple sulphur bacteria in carbonate precipitation of modern and possibly early Precambrian stromatolites
1
2011
... 微生物席上紧密相连的碳酸盐岩纹层可类比于叠层石的纹层构造,并且没有发现明显的被捕获的碳酸盐颗粒存在,所以碳酸盐的沉淀可能导致了叠层石纹层的形成.主要通过微生物的新陈代谢过程促使Lagoa Vermelha生物席中的碳酸盐析出,例如呼吸作用促使方解石在生物席表面层析出,而硫化物氧化和硫酸盐还原反应诱发了成岩过程,促进了高镁方解石和钙白云石的产生(图3 a纹层1).如图3 a叠层石纹层所示,微生物席系统的典型特征是从表层到深层逐渐多样化的生物活动[34 ,35 ] ,分别为:① ② ③ ④
Proterozoic stromatolites: The first marine evolutionary biota
1
1999
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Microbial signatures in peritidal siliciclastic sediments,a catalogue
1
2000
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
From the matground structure to the primary sedimentary structure of a fifth category: Significant concepts on sedimentology
1
2006
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
从席底构造到第 5 类原生沉积构造
1
2006
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Overview of advances in studies of primary sedimentary structures formed by microbes
1
2007
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
微生物形成的原生沉积构造研究进展综述
1
2007
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
Are cyanobacterial mats precursors of stromatolites?
2
2011
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
... 作为地球上最为古老的“宏观化石”[40 ] ,现代碳酸盐叠层石的构建扩大了我们对于古代生态系统中微生物新陈代谢活动和叠层石形成过程的理解.但还需要更深入地对一些细节性的问题进行探究,如蓝细菌生物席是否与谜一般的前寒武纪叠层石的形态学先导有关,以及鲨鱼湾叠层石中的3种经典微生物群落是否会长期稳定存在等.到目前为止,我们仅仅接触到了碳酸盐叠层石的冰山一角,最大的挑战是通过对现代碳酸盐叠层石的研究,穿越成岩作用“过滤器”,更好地应用于对古代叠层石的解释中. ...
Sulfate reducing bacteria in microbial mats: Changing paradigms, new discoveries
1
2006
... 已经达成共识的是,现代微生物席的建造主要依赖于蓝细菌[15 ,16 ,36 ,37 ,38 ,39 ] .值得强调的是,蓝细菌、硫氧化细菌和甲烷菌等均是现代微生物席重要的功能构成者[40 ,41 ] .通过对Little Darby群岛的粗粒叠层石[12 ] 、Hamelin Pool中的细粒叠层石[13 ] 以及Lagoa Vermelha潟湖中的细粒叠层石[14 ] 研究的总结,可以为我们提供一个探究寒武纪叠层石的潜在窗口,但是这个窗口还不能完全窥探复杂多变的寒武纪叠层石世界,因为古代叠层石与现代叠层石在生长形态和沉积环境等方面还存在明显差异. ...
The meaning of stromatolites
1
2013
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
Mesoproterozoic carbon dioxide levels inferred from calcified cyanobacteria
1
2007
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
Calcified cyanobacteria
2
1991
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
... [44 ]相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms
1
2000
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
The nature of stromatolites: 3,500 million years of history and a century of research
1
2011
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...
Microstructure and cyanobacterial composition of microbial mats from the High Arctic
1
2015
... 辽宁省本溪火连寨剖面张夏组中部鲕粒滩相灰岩中的叠层石生物丘,发育在III级层序中代表强迫型海退沉积的顶部;以高能鲕粒滩相背景和大型柱状叠层石为特征,从而与生长在潮下带环境中的巴哈马Little Darby群岛的穹隆状叠层石形成对比,说明现代碳酸盐叠层石的发育环境和产出形态与寒武纪叠层石存在明显差异,同时这也为环境控制微生物岩形态提供了依据[26 ] .这些柱状体单层厚度5~10 m,图4 a所示的细密向上凸起的纹层组成说明了叠层石生长的趋光性特征[42 ] ;单个柱体宽约10 cm,相互平行不向外分叉,表现为典型的高能背景下的泥晶相叠层石的特征.① 图4 b~d);这些内部由方解石充填的细管状丝状体长度约为0.2 mm,直径约20 μm,壁厚均匀.每根单独的丝状体都被清晰的微晶壁分割开,微晶壁均匀的厚度和成分表明了一个浸渍过程,可能代表蓝细菌钙化鞘的降解残余物[43 ,44 ,45 ,46 ] ,因此可以归为简单的葛万菌(Girvanella )化石;这些丝状体构成的一个相互缠绕的网络状结构与现代的织线菌(Plectonema )[44 ] 相似,没有缠绕状结构的丝状体与现代的第谷菌(Tychonema )[47 ] 相似.丝状的葛万菌(Girvanella )化石组合在潮间带、潮上带均有分布,但是存在较大的形态差异:相互缠绕的网络状丝状葛万菌(图4 b,c)分布在潮上带的小型柱状叠层石中,说明在较弱的水体环境下,丝状体受水动力环境影响较小,从而能够自由无规律地缠绕式生长.平铺式的片状分布葛万菌化石(图4 d)构成了潮间带上部的叠层石,反映了在一定动荡的水体环境下,沉积物供给速率加快,丝状蓝细菌的生长受到影响,整体表现出较粗的不明显的丝状结构.② Hedstroemla )(图4 e)出现在潮上带和潮间带叠层石中,每个基座菌菌落之间以松散的形式存在,说明微生物席受到较强的水流作用,很难完整保存下来.因此,叠层石的生长环境,尤其是水流条件是叠层石中钙化蓝细菌组合保存和分布的重要影响因素.火连寨剖面张夏组中构成柱状叠层石的丝状葛万菌(Girvanella )(图4 b~d)和球状基座菌(Hedstroemla )(图4 e)的分布模式完全不同于鲨鱼湾丝状和球状蓝细菌微生物席的分布模式,进一步说明不同微生物群落和环境条件导致了不同的构造形态[25 ] ,尽管Hamelin Pool中典型的叠层石沉积模式为古代早期复杂的生态系统提供了现代模型,但是现代叠层石的生长模式要更为复杂,还不能完全运用到古代叠层石的研究中. ...






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}