Earth's climate: Past and future
1
2007
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
1
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
Atmospheric CO2 concentrations over the last glacial termination
2
2001
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... [3 ,4 ]、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
High-resolution carbon dioxide concentration record 650,000-800,000 years before present
2
2008
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... ,4 ]、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
PA4213
4
2011
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... [5 ,11 ,12 ]和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... [
5 ,
6 ,
11 ,
15 ,
17 ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Refining a pedogenic-carbonate CO2 paleobarometer to quantify a middle Miocene greenhouse spike
3
2009
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
6 ,
11 ,
15 ,
17 ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Significant transient, p CO2 , perturbation at the New Zealand Oligocene-Miocene transition recorded by fossil plant stomata
1
2019
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
The impact of miocene atmospheric carbon dioxide fluctuations on climate and the evolution of terrestrial ecosystems
1
2008
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
Stable isotopic results from paleosol carbonate in South Asia: Paleoenvironmental reconstructions and selective alteration
1
2009
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
Paleosol barometer indicates extreme fluctuations in atmospheric CO2 across the Cretaceous-Tertiary boundary
1
2002
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
Atmospheric carbon dioxide concentration across the mid-Pleistocene transition
3
2009
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
11 ,
15 ,
17 ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Atmospheric carbon dioxide over the past 60 million years
1
2000
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
The role of carbon dioxide during the onset of Antarctic glaciation
3
2011
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 对于p CO2 与全球气候变化之响应原因,Pagani等[65 ] 通对对比p CO2 重建与C4 植被扩张记录,认为造成始新世—渐新世p CO2 逐渐降低可能与南极冰盖扩张及C4 植被扩张有关.Pagani等[13 ] 通过对始新世到渐新世界线(E-O界线)附近的p CO2 的重建发现p CO2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是p CO2 的降低的结果,而晚中新世10 Ma左右p CO2 升高则是对东南极冰盖的扩张的响应[44 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时p CO2 的降低对应北半球冰川作用加强.Palmer等[14 ] 对29 ka以来的p CO2 进行重建,认为p CO2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的p CO2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的p CO2 存在高度一致性(图3 ). ...
Multi-proxy reconstruction of surface water p CO2 in the northern Arabian Sea since 29ka
3
2010
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... 对于p CO2 与全球气候变化之响应原因,Pagani等[65 ] 通对对比p CO2 重建与C4 植被扩张记录,认为造成始新世—渐新世p CO2 逐渐降低可能与南极冰盖扩张及C4 植被扩张有关.Pagani等[13 ] 通过对始新世到渐新世界线(E-O界线)附近的p CO2 的重建发现p CO2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是p CO2 的降低的结果,而晚中新世10 Ma左右p CO2 升高则是对东南极冰盖的扩张的响应[44 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时p CO2 的降低对应北半球冰川作用加强.Palmer等[14 ] 对29 ka以来的p CO2 进行重建,认为p CO2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的p CO2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的p CO2 存在高度一致性(图3 ). ...
High Earth-system climate sensitivity determined from Pliocene carbon dioxide concentrations
5
2010
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
15 ,
17 ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Transient Middle Eocene atmospheric CO? and temperature variations
3
2010
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
A 40-million-year history of atmospheric CO2
6
2013
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
... 其中δ 13 C C O 2 a q 2(aq) 的碳同位素组成,可利用同时代有孔虫壳体的碳同位素求得[45 ,62 ] .δ13 Corg 为浮游生物的碳同位素组成,可通过烯酮碳同位素δ13 C37:2 求得[17 ,45 ,62 ] : ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
17 ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
History of seawater carbonate chemistry, atmospheric CO2 , and ocean acidification
1
2012
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
The evolution of p CO2 , ice volume and climate during the middle Miocene
2
2012
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
... 烯酮碳同位素方法从颗石藻生理作用与同位素分馏关系研究入手、通过建立动力学模型解释碳同位素变化,对溶解二氧化碳进行重建,深海沉积物中烯酮物质的碳同位素受环境因素[19 ] 影响较小.本文旨在综述利用颗石藻长链烯酮碳同位素分馏(εp37:2 )重建p CO2 的方法,介绍该方法建立的过程、影响因素、碳浓缩机制及其在古p CO2 重建中的应用. ...
B/Ca in planktonic foraminifera as a proxy for surface seawater pH
1
2007
... 二氧化碳是重要的温室气体,吸收红外辐射引起温室效应造成全球变暖,影响全球气候[1 ] .工业革命以来大气二氧化碳分压(p CO2 )迅速升高,南极冰芯的气泡二氧化碳记录显示全球p CO2 从全新世初期的240×10-6 ~280×10-6 增加到现代的411×10-6 [2 ] ,达到了近800 ka以来的最高值[3 ,4 ] .但是过去p CO2 在长时间尺度波动范围极大,仅上新世就曾达到 400×10-6[5 ] ,因此了解过去大气二氧化碳的变化对了解气候敏感性和碳循环研究至关重要.目前对于p CO2 的了解主要依据对南极冰芯记录气泡组成的p CO2 测量[3 ,4 ] 、树叶化石的气孔指数[6 ,7 ,8 ] 、古土壤次生碳酸盐结核的碳同位素[9 ,10 ] 、有孔虫硼同位素[5 ,11 ,12 ] 和生标化合物(烯酮)的稳定同位素[13 ,14 ,15 ,16 ,17 ] 分析.但每个方法存在一定的适用性和局限性,如南极冰芯记录最长只能追溯到800 ka[18 ] ;树叶化石和古土壤次生碳酸盐结核保存在陆地上容易受到成岩作用或环境因素的影响;有孔虫的硼同位素和B/Ca值计算得到pH,需要结合碱度计算p CO2 [19 ,20 ] ,且碱度无法直接测得,需要盐度或者模型推算,误差较大;烯酮碳同位素可以直接求得水溶二氧化碳CO2(aq) 的浓度,且烯酮由颗石藻产生,来源稳定,颗石藻为单细胞藻类生理作用相比于有孔虫更为单一,因此误差更小.但是烯酮重建大气二氧化碳的方法中对生长速率替代性指标的构建还不够完善.重建大气二氧化碳的方法的建立和完善,对于我们了解p CO2 的变化机制及其与气候系统间的关系起到关键作用. ...
Texas:Texas A & M University
1
1978
... 长链烯酮(long-chain alkenones)为一系列37~39个碳原子组成的碳链,且还有带2~4个碳双键的不饱和甲基或乙基酮的脂肪族化合物.长链烯酮在DSDP(Deep Sea Drilling Project)40航次的中新世到更新世的沉积物中首次被发现[21 ] ,随后发现其存在于各个大洋沉积物中.长链烯酮在沉积物中含量丰富,在全球范围内均可长时间保存[22 ,23 ,24 ] .对长链烯酮相对于其他化合物可以长时间保存的原因有几种猜测,首先其化学键的链长和不饱和键所在的位置固定,不饱和键的生物结构较为少见,使得其在沉积物中长时间保存不被细菌破坏;其次其双键结构使烯酮的生物降解作用比其他不饱和脂类更慢;最后其长链分子使得烯酮在水溶液中不易溶解,从而使其不易被微生物降解,同时烯酮可能被生物以某种形式压缩在一起使其不易被降解[23 ] . ...
Does growth rate affect ketone unsaturation and intracellular carbon isotopic variability inEmiliania huxleyi?
3
1998
... 长链烯酮(long-chain alkenones)为一系列37~39个碳原子组成的碳链,且还有带2~4个碳双键的不饱和甲基或乙基酮的脂肪族化合物.长链烯酮在DSDP(Deep Sea Drilling Project)40航次的中新世到更新世的沉积物中首次被发现[21 ] ,随后发现其存在于各个大洋沉积物中.长链烯酮在沉积物中含量丰富,在全球范围内均可长时间保存[22 ,23 ,24 ] .对长链烯酮相对于其他化合物可以长时间保存的原因有几种猜测,首先其化学键的链长和不饱和键所在的位置固定,不饱和键的生物结构较为少见,使得其在沉积物中长时间保存不被细菌破坏;其次其双键结构使烯酮的生物降解作用比其他不饱和脂类更慢;最后其长链分子使得烯酮在水溶液中不易溶解,从而使其不易被微生物降解,同时烯酮可能被生物以某种形式压缩在一起使其不易被降解[23 ] . ...
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Applications of biomarkers for delineating marine paleoclimatic fluctuations during the Pleistocene
2
1993
... 长链烯酮(long-chain alkenones)为一系列37~39个碳原子组成的碳链,且还有带2~4个碳双键的不饱和甲基或乙基酮的脂肪族化合物.长链烯酮在DSDP(Deep Sea Drilling Project)40航次的中新世到更新世的沉积物中首次被发现[21 ] ,随后发现其存在于各个大洋沉积物中.长链烯酮在沉积物中含量丰富,在全球范围内均可长时间保存[22 ,23 ,24 ] .对长链烯酮相对于其他化合物可以长时间保存的原因有几种猜测,首先其化学键的链长和不饱和键所在的位置固定,不饱和键的生物结构较为少见,使得其在沉积物中长时间保存不被细菌破坏;其次其双键结构使烯酮的生物降解作用比其他不饱和脂类更慢;最后其长链分子使得烯酮在水溶液中不易溶解,从而使其不易被微生物降解,同时烯酮可能被生物以某种形式压缩在一起使其不易被降解[23 ] . ...
... [23 ]. ...
Consistent fractionation of 13C in nature and in the laboratory: Growth-rate effects in some haptophyte algae
10
1997
... 长链烯酮(long-chain alkenones)为一系列37~39个碳原子组成的碳链,且还有带2~4个碳双键的不饱和甲基或乙基酮的脂肪族化合物.长链烯酮在DSDP(Deep Sea Drilling Project)40航次的中新世到更新世的沉积物中首次被发现[21 ] ,随后发现其存在于各个大洋沉积物中.长链烯酮在沉积物中含量丰富,在全球范围内均可长时间保存[22 ,23 ,24 ] .对长链烯酮相对于其他化合物可以长时间保存的原因有几种猜测,首先其化学键的链长和不饱和键所在的位置固定,不饱和键的生物结构较为少见,使得其在沉积物中长时间保存不被细菌破坏;其次其双键结构使烯酮的生物降解作用比其他不饱和脂类更慢;最后其长链分子使得烯酮在水溶液中不易溶解,从而使其不易被微生物降解,同时烯酮可能被生物以某种形式压缩在一起使其不易被降解[23 ] . ...
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 式中:b 为光合作用过程中所有影响总碳同位素分馏的生理作用值,包括生长速率和细胞体表面积比值[24 ,56 ,62 ] ;εf 为光合作用中的碳同位素分馏[50 ,51 ] . ...
... b 值的经验公式计算最早由Bidigare等[24 ] 提出,根据不同地区的现场观察和实验室连续培养发现b 值与磷酸盐浓度[PO4 3 - 4 3 - 4 3 - 4 3 - b 值的关系式: ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
... [24 ]数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
... (a)不同培养环境中ε p μ /CO2 的比值,(b)不同光照条件下ε p μ /CO2 的比值,数据来自参考文献[80 ];(a)连续培养数据来自参考文献[24 ],分批培养数据来参考文献[63 ] ...
... εp versus μ /CO2 from different cultures(a), εp versus μ /CO2 from different irradiance experiments(b) (data from reference[80 ]);Chemostat incubation data from reference[24 ], dilute batch cultures data from reference[63 ] ...
Long-chain alkenes and alkenones in the marine coccolithophorid Emiliania huxleyi
2
1980
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
... [25 ,26 ,34 ,35 ,36 ,37 ],此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Long chain (n-C37-C39) alkenones in the Prymnesiophyceae: Distribution of alkenones and other lipids and their taxonomic significance
2
1984
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
... ,26 ,34 ,35 ,36 ,37 ],此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Long-chain alkenones and alkyl alkenoates and the fossil coccolith record of marine sediments
2
1990
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
... [27 ],但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Alkenones in Gephyrocapsa oceanica:Implications for studies of paleoclimate
1
1995
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Global synchroneity of late Quaternary coccolith datum levels Validation by oxygen isotopes
1
1977
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Essai d‘établissement d'une biostratigraphie du nannoplancton calcaire dans le Pleistocéne de I'Atlantique Nord‐oriental
1
1977
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Taxonomy, Evolution, and Biostratigraphy of Late Triassic-Early Jurassic Calcareous Nannofossils
1
1987
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Alkenones in cretaceous black shales, Blake-Bahama Basin, western North Atlantic
1
1986
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Recognition of alkenones in a lower Aptian porcellanite from the west-central Pacific
1
2004
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
PA1202
1
2012
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Coccolith δ18 O and alkenone records in middle Pliocene orbitally controlled deposits: High-frequency temperature and salinity variations of sea surface water
1
2007
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Long chain alkenones in the Early Pliocene Sicilian sediments (Trubi Formation—Punta di Maiata section): Implications for the alkenone paleothermometry
1
2011
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
Glacial-interglacial productivity changes recorded by alkenones and microfossils in late Pliocene eastern equatorial Pacific and Atlantic upwelling zones
2
2010
... 现代大洋中烯酮的生产者主要为Emiliania huxleyi 和Gephrocapsa oceanica [25 ,26 ,27 ,28 ] .其中E. huxleyi 在地质记录中最早出现于0.27 Ma[29 ] 、G.oceanca 最早出现于1.85 Ma[30 ] ,但长链烯酮在地质记录中最早可追溯到白垩纪[31 ,32 ,33 ] ,更早期的生产者被认为是Noelaerhabdaceae科的其他属种,如Reticulofenestra spp. ,Cyclicargolithus spp.和Pseudoemiliania lacunosa 等[25 ,26 ,34 ,35 ,36 ,37 ] ,此外还有非钙化的现代生产者如Isochrysis galbana [27 ] ,但主要分布在近岸,不作为开阔大洋的烯酮来源来讨论. ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Biotic and geochemical evidence for a global latitudinal shift in ocean biogeochemistry and export productivity during the late Pliocene
1
2011
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
Further evaluation of long-chain alkenones as indicators of paleoceanographic conditions
2
1988
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
... [39 ,40 ,41 ,42 ]、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
The South Atlantic in the Late Quaternary: Reconstruction of Material Budgets and Current Systems
1
2004
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
Sea surface temperatures and environmental conditions during the “warm Pliocene” interval (~ 4.1-3.2 Ma) in the Eastern Mediterranean (Cyprus)
1
2017
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
Calibration of unsaturation patterns in long-chain ketone compositions for palaeotemperature assessment
1
1987
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
Late miocene atmospheric CO2 concentrations and the expansion of C4 grasses
2
1999
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
Miocene evolution of atmospheric carbon dioxide
5
1999
... 长链烯酮由于其可以长时间保存,含量丰富且不易受到溶解作用的影响,在古环境研究中应用广泛.作为常用替代性指标,长链烯酮堆积速率可以指示古生产力[38 ,39 ] 、烯酮不饱和度(U37 K ' [39 ,40 ,41 ,42 ] 、C37:2 烯酮碳同位素δ13 C可以用于计算海水及大气p CO2 [17 ,43 ,44 ] . ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... [44 ]的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... 对于p CO2 与全球气候变化之响应原因,Pagani等[65 ] 通对对比p CO2 重建与C4 植被扩张记录,认为造成始新世—渐新世p CO2 逐渐降低可能与南极冰盖扩张及C4 植被扩张有关.Pagani等[13 ] 通过对始新世到渐新世界线(E-O界线)附近的p CO2 的重建发现p CO2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是p CO2 的降低的结果,而晚中新世10 Ma左右p CO2 升高则是对东南极冰盖的扩张的响应[44 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时p CO2 的降低对应北半球冰川作用加强.Palmer等[14 ] 对29 ka以来的p CO2 进行重建,认为p CO2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的p CO2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的p CO2 存在高度一致性(图3 ). ...
A carbon isotope record of CO2 levels during the late Quaternary
7
1990
... 碳同位素分馏εp 指环境中的溶解二氧化碳与生物进行光合作用、形成有机碳过程的碳同位素分馏[45 ,46 ,47 ] . ...
... 式中:A和B为常数,数值根据不同的海洋环境而定.随后Jasper等[45 ,62 ] 提出碳同位素分馏的公式(1). ...
... 其中δ 13 C C O 2 a q 2(aq) 的碳同位素组成,可利用同时代有孔虫壳体的碳同位素求得[45 ,62 ] .δ13 Corg 为浮游生物的碳同位素组成,可通过烯酮碳同位素δ13 C37:2 求得[17 ,45 ,62 ] : ...
... ,45 ,62 ]: ...
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 由公式(8)可知,扩散模型中εp 不仅随[CO2(aq) ]变化,还与藻类生长参数有关,为了方便计算,Jasper等[45 ] 将公式(2)和公式(3)简写为: ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
Fractionation of carbon isotopes by phytoplankton and estimates of ancient CO2 levels
1
1992
... 碳同位素分馏εp 指环境中的溶解二氧化碳与生物进行光合作用、形成有机碳过程的碳同位素分馏[45 ,46 ,47 ] . ...
The post-Paleozoic chronology and mechanism of 13 C depletion in primary marine organic matter
1
1989
... 碳同位素分馏εp 指环境中的溶解二氧化碳与生物进行光合作用、形成有机碳过程的碳同位素分馏[45 ,46 ,47 ] . ...
Metabolic fractionation of carbon isotopes in marine plankton—I. Temperature and respiration experiments
1
1968
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
The relationship between delta 13 C of organic matter and [CO2(aq) ] in ocean surface water: Data from a JGOFS site in the northeast Atlantic Ocean and a model
3
1992
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
... ,49 ,50 ]、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
Changes in the δ13 C of surface water particulate organic matter across the subtropical convergence in the SW Indian Ocean
7
1993
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
... 式中:γ 表示假设的固定的CO2 需求量.Francois等[50 ] 发现在被动扩散模型中除了[CO2(aq) ]的影响,细胞渗透性、表面积和生长速率(μ )也会影响εp ,并引入了羧化速率(G )的定义: ...
... 式中:C 为细胞内碳含量.根据Francois的假设,二氧化碳流入和流出的细胞膜的渗透率相等,也就是k 1 =k -1 时[50 ] ,公式(2)可以由以上公式代入并简化为: ...
... Rau等[59 ] 总结了前人的工作,总结了εp 的影响因素包括Ce 、细胞半径、细胞生长速率、细胞膜的渗透性(P)、温度、一定的pH和盐度等[50 ,53 ,55 ] ,并将各个影响因子写入模型中: ...
... 式中:b 为光合作用过程中所有影响总碳同位素分馏的生理作用值,包括生长速率和细胞体表面积比值[24 ,56 ,62 ] ;εf 为光合作用中的碳同位素分馏[50 ,51 ] . ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
On the relationship between carbon isotope discrimination and the intercellular carbon dioxide concentration in leaves
2
1982
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
... 式中:b 为光合作用过程中所有影响总碳同位素分馏的生理作用值,包括生长速率和细胞体表面积比值[24 ,56 ,62 ] ;εf 为光合作用中的碳同位素分馏[50 ,51 ] . ...
Carbon isotope discrimination and photosynthesis
1
1989
... 式中:δ 13 C C O 2 a q 13 Corg 是有机物的碳同位素,可通过地质记录直接测得;εp 的主要影响因素是溶解二氧化碳浓度[CO2(aq) ][48 ,49 ] ,其次还有细胞生长速率μ [24 ,49 ,50 ] 、细胞体积与表面积比值V/SA[22 ] .对于εp 值的变化使用C3 植物的扩散模型解释:CO2(aq) 随二氧化碳浓度梯度进入和流出细胞,细胞内二氧化碳到达叶绿体进行光合作用,这2个过程为碳运输过程;到达叶绿体后的CO2(aq) 通过光合作用固定形成有机碳,称为碳固定过程.εp 值即为碳运输和碳固定过程中的碳同位素分馏[51 ,52 ] : ...
Dependence of phytoplankton carbon isotopic composition on growth rate and [CO2(aq) ]: Theoretical considerations and experimental results
9
1995
... 式中:εf 和εt 均为常数,分别代表碳固定和碳被动扩散过程中的碳同位素分馏系数,Ce 和Ci 分别代表细胞外和细胞内二氧化碳浓度.εf 值为所有与碳固定有关的同位素变化值,包括Rubisco酶,β羧化酶等催化反应[53 ,54 ] .Goericke等[55 ] 测量了现代海水表层不同纬度浮游植物有机物的εf 值为25‰ ‰ . Hayes[56 ] 将浓度梯度Ce -Ci 考虑在内,重新整理了公式(2)得到: ...
... 式中:k 1 和k -1 为扩散进入和离开细胞的速率常数,k 与细胞膜渗透性有关,假设进入和流出细胞的速率常数相等,此时k 1 =k -1 .Laws等[53 ] 结合培养数据和现场观察发现,不考虑呼吸作用和光呼吸作用时,μ /[CO2(aq) ]与εp 存在线性相关,因此生长速率又可以表示为: ...
... 式中:k 是细胞膜对二氧化碳的渗透率与表面积乘积[53 ,57 ] ,细胞内碳含量C 与细胞体积成比例[58 ] ,因此公式(6)可以整理为: ...
... Rau等[59 ] 总结了前人的工作,总结了εp 的影响因素包括Ce 、细胞半径、细胞生长速率、细胞膜的渗透性(P)、温度、一定的pH和盐度等[50 ,53 ,55 ] ,并将各个影响因子写入模型中: ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... [53 ,55 ],细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Organic carbon 13 C variations in sedimentary rocks as chemostratigraphic and paleoenvironmental tools
2
1997
... 式中:εf 和εt 均为常数,分别代表碳固定和碳被动扩散过程中的碳同位素分馏系数,Ce 和Ci 分别代表细胞外和细胞内二氧化碳浓度.εf 值为所有与碳固定有关的同位素变化值,包括Rubisco酶,β羧化酶等催化反应[53 ,54 ] .Goericke等[55 ] 测量了现代海水表层不同纬度浮游植物有机物的εf 值为25‰ ‰ . Hayes[56 ] 将浓度梯度Ce -Ci 考虑在内,重新整理了公式(2)得到: ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Variations of marine plankton δ13 C with latitude, temperature, and dissolved CO2 in the world ocean
5
1994
... 式中:εf 和εt 均为常数,分别代表碳固定和碳被动扩散过程中的碳同位素分馏系数,Ce 和Ci 分别代表细胞外和细胞内二氧化碳浓度.εf 值为所有与碳固定有关的同位素变化值,包括Rubisco酶,β羧化酶等催化反应[53 ,54 ] .Goericke等[55 ] 测量了现代海水表层不同纬度浮游植物有机物的εf 值为25‰ ‰ . Hayes[56 ] 将浓度梯度Ce -Ci 考虑在内,重新整理了公式(2)得到: ...
... Rau等[59 ] 总结了前人的工作,总结了εp 的影响因素包括Ce 、细胞半径、细胞生长速率、细胞膜的渗透性(P)、温度、一定的pH和盐度等[50 ,53 ,55 ] ,并将各个影响因子写入模型中: ...
... 并提出b 值是关于εf 的函数:b =(εf -εp )Ce ,其中εf 值为25‰ ‰ [55 ] ,Pagani[60 ] 计算了不同εf 值时的b 值下的[CO2(aq) ],随后总结了不同εf 下的b 值(图1 ),可通过[PO4 3 - εf 值时的b 值: ...
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Factors controlling 13 C contents of sedimentary organic compounds: Principles and evidence
2
1993
... 式中:εf 和εt 均为常数,分别代表碳固定和碳被动扩散过程中的碳同位素分馏系数,Ce 和Ci 分别代表细胞外和细胞内二氧化碳浓度.εf 值为所有与碳固定有关的同位素变化值,包括Rubisco酶,β羧化酶等催化反应[53 ,54 ] .Goericke等[55 ] 测量了现代海水表层不同纬度浮游植物有机物的εf 值为25‰ ‰ . Hayes[56 ] 将浓度梯度Ce -Ci 考虑在内,重新整理了公式(2)得到: ...
... 式中:b 为光合作用过程中所有影响总碳同位素分馏的生理作用值,包括生长速率和细胞体表面积比值[24 ,56 ,62 ] ;εf 为光合作用中的碳同位素分馏[50 ,51 ] . ...
Effect of growth rate and CO2 concentration on carbon isotopic fractionation by the marine diatom Phaeodactylum tricornutum
3
1997
... 式中:k 是细胞膜对二氧化碳的渗透率与表面积乘积[53 ,57 ] ,细胞内碳含量C 与细胞体积成比例[58 ] ,因此公式(6)可以整理为: ...
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Effect of phytoplankton cell geometry on carbon isotopic fractionation
5
1998
... 式中:k 是细胞膜对二氧化碳的渗透率与表面积乘积[53 ,57 ] ,细胞内碳含量C 与细胞体积成比例[58 ] ,因此公式(6)可以整理为: ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... [58 ],从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... 式中:V∶SAfossil 为每个样品的颗石形态,V∶SAEhux 为常数,反应当代颗石藻E.huxleyi 的细胞大小.来自连续培养数据显示r =(2.6±0.3)μm;V∶SAEhux =(0.9±0.1)μm[58 ] . ...
A model of photosynthetic 13 C fractionation by marine phytoplankton based on diffusive molecular CO2 uptake
2
1996
... Rau等[59 ] 总结了前人的工作,总结了εp 的影响因素包括Ce 、细胞半径、细胞生长速率、细胞膜的渗透性(P)、温度、一定的pH和盐度等[50 ,53 ,55 ] ,并将各个影响因子写入模型中: ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
The alkenone-CO2 proxy and ancient atmospheric carbon dioxide
3
2002
... 利用εp 重建[CO2(aq) ]的方法基于2点假设:① 2(aq) ;② 2(aq) 从环境中进入碳固定位点的方式为简单扩散[60 ] .Hayes等[61 ] 首次将εp 作为p CO2 指标应用到沉积物组分中生物标志物中的εp 分析中,并将εp 与[CO2(aq) ]转化为简单对数公式[61 ] : ...
... 并提出b 值是关于εf 的函数:b =(εf -εp )Ce ,其中εf 值为25‰ ‰ [55 ] ,Pagani[60 ] 计算了不同εf 值时的b 值下的[CO2(aq) ],随后总结了不同εf 下的b 值(图1 ),可通过[PO4 3 - εf 值时的b 值: ...
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Isotopic compositions and probable origins of organic molecules in the Eocene Messel shale
2
1987
... 利用εp 重建[CO2(aq) ]的方法基于2点假设:① 2(aq) ;② 2(aq) 从环境中进入碳固定位点的方式为简单扩散[60 ] .Hayes等[61 ] 首次将εp 作为p CO2 指标应用到沉积物组分中生物标志物中的εp 分析中,并将εp 与[CO2(aq) ]转化为简单对数公式[61 ] : ...
... [61 ]: ...
NATO ASI Series (Series I: Global Environmental Change), vol 17
5
1994
... 式中:A和B为常数,数值根据不同的海洋环境而定.随后Jasper等[45 ,62 ] 提出碳同位素分馏的公式(1). ...
... 其中δ 13 C C O 2 a q 2(aq) 的碳同位素组成,可利用同时代有孔虫壳体的碳同位素求得[45 ,62 ] .δ13 Corg 为浮游生物的碳同位素组成,可通过烯酮碳同位素δ13 C37:2 求得[17 ,45 ,62 ] : ...
... ,62 ]: ...
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 式中:b 为光合作用过程中所有影响总碳同位素分馏的生理作用值,包括生长速率和细胞体表面积比值[24 ,56 ,62 ] ;εf 为光合作用中的碳同位素分馏[50 ,51 ] . ...
The effects of varying CO2 concentration on lipid composition and carbon isotope fractionation in Emiliania huxleyi
5
2000
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
... (a)不同培养环境中ε p μ /CO2 的比值,(b)不同光照条件下ε p μ /CO2 的比值,数据来自参考文献[80 ];(a)连续培养数据来自参考文献[24 ],分批培养数据来参考文献[63 ] ...
... εp versus μ /CO2 from different cultures(a), εp versus μ /CO2 from different irradiance experiments(b) (data from reference[80 ]);Chemostat incubation data from reference[24 ], dilute batch cultures data from reference[63 ] ...
Carbon isotope variability in monosaccharides and lipids of aquatic algae and terrestrial plants
1
2002
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
Marked decline in atmospheric carbon dioxide concentrations during the Paleogene
4
2005
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 对于p CO2 与全球气候变化之响应原因,Pagani等[65 ] 通对对比p CO2 重建与C4 植被扩张记录,认为造成始新世—渐新世p CO2 逐渐降低可能与南极冰盖扩张及C4 植被扩张有关.Pagani等[13 ] 通过对始新世到渐新世界线(E-O界线)附近的p CO2 的重建发现p CO2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是p CO2 的降低的结果,而晚中新世10 Ma左右p CO2 升高则是对东南极冰盖的扩张的响应[44 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时p CO2 的降低对应北半球冰川作用加强.Palmer等[14 ] 对29 ka以来的p CO2 进行重建,认为p CO2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的p CO2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的p CO2 存在高度一致性(图3 ). ...
... ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
<strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
65 ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Alkenone and boron-based Pliocene p CO2 records
6
2010
... 式中:Δδ 为烯酮碳同位素的不饱和度,其不受生长速率影响[24 ,45 ,62 ] ,培养数据显示Δδ 值变化范围为3.1‰ ‰ [63 ,64 ] .在p CO2 重建中,Δδ 值通常为4.2‰ [13 ,15 ,16 ,65 ,66 ] . ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... 对于p CO2 与全球气候变化之响应原因,Pagani等[65 ] 通对对比p CO2 重建与C4 植被扩张记录,认为造成始新世—渐新世p CO2 逐渐降低可能与南极冰盖扩张及C4 植被扩张有关.Pagani等[13 ] 通过对始新世到渐新世界线(E-O界线)附近的p CO2 的重建发现p CO2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是p CO2 的降低的结果,而晚中新世10 Ma左右p CO2 升高则是对东南极冰盖的扩张的响应[44 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时p CO2 的降低对应北半球冰川作用加强.Palmer等[14 ] 对29 ka以来的p CO2 进行重建,认为p CO2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的p CO2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的p CO2 存在高度一致性(图3 ). ...
... ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
<strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
66 ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Controls on the molecular distribution and carbon isotopic composition of alkenones in certain haptophyte algae
2
2001
... Laws等[67 ] 通过全球数据对比,对公式(12)进行了校正: ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
Influence of nutrients on carbon isotope fractionation by natural populations of Prymnesiophyte algae in NE Pacific
0
1999
Controls on the carbon isotopic composition of southern ocean phytoplankton
0
1999
Biomarker-Based Inferences of Past Climate: The Alkenone p CO2 Proxy
2
2014
... 长链烯酮δ13 C重建p CO2 的方法假设:① ② b 值与[PO4 3 - ③ εf 值(25‰ ‰ b 值[70 ] . ...
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
Latitudinal variations in plankton |[delta]|13 C: Implications for CO2 and productivity in past oceans
1
1989
... 但该计算方法中的误差包括:① b 值时参数[PO4 3 - 4 3 - [66 ,70 ] ,与过去海水[PO4 3 - ② 2(aq) 转换p CO2 时利用亨利定律(Herry’s law)计算,需要用到同时代的古温度值,而古温度估算主要来自于生物标志物指标如TEX86和U37 k ' p CO2 值造成一定误差;③ p CO2 的方法依赖于扩散作用模型[公式(3)],但[CO2(aq) ]不是εp 值的唯一影响因素[公式(6)],还包括细胞几何特征[58 ] 、生长速率[24 ,49 ,50 ,53 ] .而培养环境如光照、温度和营养可利用性对细胞几何特征和生长速率又有影响[63 ,67 ,71 ] . ...
Influence of CO2 and nitrogen limitation on the coccolith volume of Emiliania huxleyi (Haptophyta)
2
2012
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Calcite production by coccolithophores in the south east Pacific Ocean
1
2008
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
Coccolithophore cell size and the Paleogene decline in atmospheric CO2
2
2008
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
... [74 ,75 ,76 ]对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
Refining ancient carbon dioxide estimates: Significance of coccolithophore cell size for alkenone‐based p CO2 records
1
2007
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
Coccolithophore size rules—Reconstructing ancient cell geometry and cellular calcite quota from fossil coccoliths
1
2008
... 培养数据显示颗石藻的细胞大小会影响εP 值[53 ,57 ] ,CO2(aq) 进入和流出细胞的通量与颗石藻表面积成正比[53 ,55 ] ,细胞内碳含量数值可通过细胞体积进行估算[58 ] .颗石大小与细胞大小成正比,可通过测量颗石的几何参数计算颗石大小从而估算细胞大小[72 ,73 ,74 ] .培养实验、现场观测和沉积记录均显示培养数据显示更大的细胞对应更高的体表面积比值,εp 值更低[58 ] ,从而对p CO2 的估算产生影响.Henderiks等[74 ,75 ,76 ] 对颗石藻大小进行校正使得颗石藻大小对模型的不确定性减小: ...
Co-variation of metabolic rates and cell-size in coccolithophores
1
2015
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Influence of cell cycle phase on calcification in the coccolithophore Emiliania huxleyi
1
2008
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Biogenic sediments in the Eauripic Rise of the equatorial western Pacific during the last 265 kyr
1
1996
... 大量室内培养、现场观察数据证明εp 与μ /[CO2(aq) ]存在线性相关,浮游植物的生长速率μ 会影响εp 值(图2 )[22 ,50 ,53 ,54 ,55 ] ,Bidigare等[24 ] 通过在氮营养盐限制,连续光照培养中发现E.huxleyi εp 与μ /[CO2(aq) ]负相关,Ribesell等[63 ] 在营养盐充足、16小时光照8小时黑暗(16∶8 h)循环光照条件下分批次培养E.huxleyi 并对比Bidigare等[24 ] 数据发现,后者εp 与μ /[CO2(aq) ]比值更高,趋势更陡,说明不同培养环境差异影响μ 值改变、从而影响碳同位素分馏.培养数据显示生长速率μ 会受到环境变化包括p CO2 、光照、温度、营养盐可利用性(氮、磷、铁等)的影响[72 ,77 ,78 ] ,生长速率的变化会改变细胞碳需求,改变细胞内部、细胞表面和细胞外周围的[CO2(aq) ][59 ] .Pagani等[44 ] 认为应在开阔大洋低生产力地区,表层水[PO4 3 - 13 C以减小对μ 值的影响.同时颗石Sr/Ca比和烯酮堆积速率可以反应海区生产力[37 ,79 ] ,结合生产力与b 值对照,选择生产力波动较小区域数据、降低μ 值对b 值的影响、从而减小模型的不确定性. ...
Light-dependent carbon isotope fractionation in the coccolithophorid Emiliania huxleyi
3
2002
... (a)不同培养环境中ε p μ /CO2 的比值,(b)不同光照条件下ε p μ /CO2 的比值,数据来自参考文献[80 ];(a)连续培养数据来自参考文献[24 ],分批培养数据来参考文献[63 ] ...
... εp versus μ /CO2 from different cultures(a), εp versus μ /CO2 from different irradiance experiments(b) (data from reference[80 ]);Chemostat incubation data from reference[24 ], dilute batch cultures data from reference[63 ] ...
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Utilization of inorganic carbon by marine microalgae
1
1987
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
The diversity of inorganic carbon acquisition mechanisms in eukaryotic microalgae
1
2002
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
CO2 concentrating mechanisms in algae: Mechanisms, environmental modulation, and evolution
1
2005
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Inorganic carbon acquisition by eukaryotic algae: Four current questions
1
2010
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Extra- and intra-cellular carbonic anhydrase in relation to culture age in a high-calcifying strain of Emiliania huxleyi lohmann
1
1994
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
The development of a CO2 -concentrating mechanism in Emiliania huxleyi
2
1996
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
Inorganic carbon transport in relation to culture age and inorganic carbon concentration in a high-calcifying strain of Emiliania huxleyi (Prymnesiophyceae) 1
1
1996
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Bicarbonate utilization by marine phytoplankton species
1
2010
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Acquisition and use of bicarbonate by Emiliania huxleyi
1
2002
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Sources of inorganic carbon for marine microalgal photosynthesis: A reassessment of delta 13 C data from batch culture studies of thalassiosira pseudonana and Emiliania huxleyi
1
1998
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
Effect of CO2 concentration on the carbon acquisition of bloom-forming marine phytoplankton
2
2003
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
Increased CO2 and iron availability effects on carbon assimilation and calcification on the formation of Emiliania huxleyi blooms in a coastal phytoplankton community
2
2018
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
Carbon acquisition of marine phytoplankton: Effect of photoperiod length
3
2006
... 长链烯酮重建p CO2 模型的可行性建立在被动扩散模型上[60 ] ,即被动扩散CO2(aq) 进行光合作用,但是对比过去p CO2 记录,现代p CO2 浓度值较低,许多浮游藻类(如硅藻和鞭毛藻类)演化出对碳主动吸收机制即碳浓缩机制(CO2 Concentrating Mechanism,CCM),利用碳酸酐酶(CA)转化HCO3 - 2(aq) 供藻类利用[53 ,57 ,81 ,82 ,83 ,84 ] .对于颗石藻是否存在CCM仍有争议,CCM的存在会对ɛp 值产生影响,从而影响对p CO2 的估算.Nimer等[85 ] 通过培养重钙化程度的E. huxleyi Bigelow(No.8E)和轻钙化程度的E. huxleyi Lohmann(No.79)发现颗石藻在稳定期的培养环境中溶解无机碳(Dissolved Inorganic Carbon,DIC)耗尽,钙化速率降低到60%时,重钙化程度的E. huxleyi 同时检测到了体外碳酸酐酶(eCA)和体内碳酸酐酶(iCA)的活性.Nimer等[86 ] 通过培养实验发现E. huxleyi 体内[CO2(aq) ]在不同pH情况下是体外[CO2(aq) ]的几倍,认为E.huxleyi 存在逆浓度梯度主动运输CO2(aq) 的能力.同年培养实验发现培养环境中[CO2(aq) ]降低时E.huxleyi 中钙化程度中等类型E.huxleyi (Lohmann)Hay&Mohler会利用HCO3 - [87 ] .Nimer等[88 ] 通过培养实验发现在培养稳定期低DIC浓度下可检测到E.huxleyi 细胞体外碳酸酐酶(eCA)的活性,但在G.oceanica 未发现eCA活性,认为G.oceanica 可能存在阴离子交换机制利用HCO3 - [89 ] 对E.huxleyi (PCC.B11)进行培养时发现在任何DIC浓度下都存在离子交换通道吸收HCO3 - 2 和HCO3 - E.huxleyi 存在2种HCO3 - [90 ] 通过分批培养的E.huxleyi δ13 C和生长速率数据发现,E.huxleyi 在低生长速率时DIC主要利用HCO3 - 2 的吸收.Rost等[80 ] 在不同光照强度下对E.huxleyi (PML B92/11)进行分批次培养时,发现光照强度增加导致εp 降低,认为εp 降低主要由HCO3 - [91 ] 在不同[CO2(aq) ]条件下的培养实验中发现,颗石藻E.huxleyi 可以主动吸收HCO3 - p CO2 浓度发生改变,室外培养实验也得到相同的结果[92 ] .不同光周期环境下分批次培养E.huxleyi (B92/11),发现p CO2 浓度降低时CO2 渗出增加,认为E.huxleyi CCM机制较低,同时在光照循环条件下E.huxleyi 会利用HCO3 - 2 演化中半饱和浓度K1/2 降低,说明CCM机制效率低,同时未检测到eCA活性[93 ] . ...
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
... [93 ,97 ,98 ].如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
CO2 -concentrating mechanisms in three southern hemisphere strains of Emiliania huxleyi
2
2013
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
... ,94 ,96 ],有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
The role of extracellular carbonic anhydrase activity in inorganic carbon utilization of Phaeocystis globosa (Prymnesiophyceae): A comparison with other marine algae using the isotopic disequilibrium technique
1
2000
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
Utilization of inorganic carbon by the coccolithophorid Emiliania huxleyi (Lohmann) Kamptner
1
1992
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
Isotope disequilibrium and mass spectrometric studies of inorganic carbon acquisition by phytoplankton
1
2007
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
Carbon concentrating mechanisms in eukaryotic marine phytoplankton
1
2011
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
A diatom record of CO2 decline since the late Miocene
1
2017
... 总的来说,基于培养实验数据,部分重钙化的E.huxleyi 属种如PML B92/11利用HCO3 - [93 ,94 ] ,E.huxleyi 的部分属种使用eCA催化水解反应提高细胞外CO2(aq) 浓度从而利用CO2 作为碳源[95 ] .培养数据显示E.huxleyi 的eCA不随CO2 的改变发生变化[91 ,92 ] .有的培养实验和现场实验认为E.huxleyi 存在CCMs机制[86 ,94 ,96 ] ,有的培养实验认为E.huxleyi 不存在CCMs或者CCMs的效率比较低[93 ,97 ,98 ] .如果考虑到藻类CCMs机制的存在,基于有机质δ13 C的p CO2 数据会比实际数据偏高[99 ] ,因此,了解CCMs机制对提高长时间尺度中长链烯酮p CO2 重建精度至关重要. ...
CO2 drawdown following the middle Miocene expansion of the Antarctic Ice Sheet
3
2013
... Jasper等[45 ] 最早将长链烯酮重建p CO2 方法应用在墨西哥湾北部100 ka的沉积物记录中,随后长链烯酮重建p CO2 的方法得到了广泛应用,大量的研究结果恢复了自中始新世以来的p CO2 记录,并探讨了p CO2 与全球气候之间的响应.记录显示p CO2 自新生代以来逐渐降低,其中中始新世p CO2 较高,约为2 000×10-6 ,随后晚始新世降低到1 000×10-6 [16 ] .晚渐新世时p CO2 降低至约350×10-6 ,随后在25 Ma左右又快速降低,早中新世到中中新世p CO2 维持在较低的水平(260×10-6 ~190×10-6 )[44 ] ,但中中新世的p CO2 存在争议,Badger等[100 ] 的数据显示中中新世(14 Ma)p CO2 约为300×10-6 ,比Pagani等[44 ] 的p CO2 数据高.晚中新世时期10 Ma左右p CO2 升高至320×10-6 ~250×10-6[43 ] .Seki等[66 ] 的p CO2 数据显示5 Ma是上新世最温暖的时期,p CO2 达到300×10-6 ~400×10-6 ,p CO2 降低至工业革命前水平274×10-6 ~285×10-6 .Pagani等[15 ] 的数据显示p CO2 在早上新世4~5 Ma,p CO2 浓度的均值为390×10-6 ~280×10-6 , 4.5 Ma时p CO2 和温度达到最高,36×10-6 ~415×10-6 ,随后5~0.5 Ma时p CO2 持续下降.Zhang等[17 ] 对单个站位40 Ma以来长时间尺度p CO2 记录重建显示,p CO2 在中中新世浓度最高,平均400×10-6 ~500×10-6 ,随着中中新世气候转型约14 Ma时p CO2 浓度下降,总体趋势大致与前人的研究工作一致.29 ka以来的p CO2 记录显示,p CO2 在11~17 ka时快速上升[14 ] .对比其他p CO2 重建指标包括浮游有孔虫硼同位素,土壤次生碳酸盐等数据,结果基本一致,显示新生代以来p CO2 在始新世极热事件达到峰值,渐新世逐渐降低,晚中新世小幅度升高至450×10-6 左右后上新世逐渐降低至工业革命前的280×10-6 . ...
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
100 ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Plio-Pleistocene climate sensitivity evaluated using high-resolution CO2 records
2
2015
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
101 ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Middle Miocene climate instability associated with high-amplitude CO2 variability
2
2014
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
102 ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
Paleoatmospheric signatures in neogene fossil leaves
2
1993
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
103 ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
The impact of Miocene atmospheric carbon dioxide fluctuations on climate and the evolution of terrestrial ecosystems
2
2008
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
104 ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
An Early Pleistocene atmospheric CO2 record based on pedogenic carbonate from the Chinese loess deposits
2
2015
... 对于
p CO
2 与全球气候变化之响应原因,Pagani等
[65 ] 通对对比
p CO
2 重建与C
4 植被扩张记录,认为造成始新世—渐新世
p CO
2 逐渐降低可能与南极冰盖扩张及C
4 植被扩张有关.Pagani等
[13 ] 通过对始新世到渐新世界线(E-O界线)附近的
p CO
2 的重建发现
p CO
2 的降低早于南极冰盖扩张时间,认为南极冰盖的扩张是
p CO
2 的降低的结果,而晚中新世10 Ma左右
p CO
2 升高则是对东南极冰盖的扩张的响应
[44 ] .Seki等
[66 ] 的
p CO
2 数据显示5 Ma是上新世最温暖的时期,3.2~2.8 Ma时
p CO
2 的降低对应北半球冰川作用加强.Palmer等
[14 ] 对29 ka以来的
p CO
2 进行重建,认为
p CO
2 在11~17 ka快速上升,海气交换加强,可能受到东亚季风的影响.对比烯酮碳同位素重建的
p CO
2 与其他指标包括硼同位素、叶片气孔和碳酸盐岩重建的
p CO
2 存在高度一致性(图
3 ).
图3 不同指标下的<strong>50 Ma</strong>以来<strong><i>p</i></strong><strong>CO<sub>2</sub></strong>记录<sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> <strong>Cenozoic <i>p</i>CO<sub>2</sub> record from different proxies</strong><sup>[<xref ref-type="bibr" rid="R5">5</xref>,<xref ref-type="bibr" rid="R6">6</xref>,<xref ref-type="bibr" rid="R11">11</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R65">65</xref>,<xref ref-type="bibr" rid="R66">66</xref>,<xref ref-type="bibr" rid="R100">100</xref>,<xref ref-type="bibr" rid="R101">101</xref>,<xref ref-type="bibr" rid="R102">102</xref>,<xref ref-type="bibr" rid="R103">103</xref>,<xref ref-type="bibr" rid="R104">104</xref>,<xref ref-type="bibr" rid="R105">105</xref>]</sup> Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...
... ,
105 ]
Fig. 3 ![]()
6 结论与展望 本文介绍了长链烯酮重建大气二氧化碳的方法模型和限制.长链烯酮重建大气二氧化碳的方法是通过长链烯酮的碳同位素计算海水中溶解二氧化碳浓度,进而利用亨利定律使用温度和溶解二氧化碳浓度求得大气二氧化碳浓度.该方法建立在C3 植物的二氧化碳被动扩散模型上,即假设环境中溶解二氧化碳通过扩散机制进入细胞内,通过光合作用进行有机碳固定,同时考虑生长速率、细胞体表面积比值和主动运输机制对ɛp 值的影响.碳浓缩机制主要存在于高等陆生植物和海洋大型浮游藻类中,该机制的出现被认为是对现代较低的二氧化碳浓度的响应.颗石藻中是否存在碳浓缩机制还存在争议,碳浓缩机制的存在会对长时间尺度上大气二氧化碳的重建产生影响.对比长链烯酮重建的大气二氧化碳记录和其他大气二氧化碳指标记录显示,新生代以来大气p CO2 在始新世最高、随后逐渐降低. ...




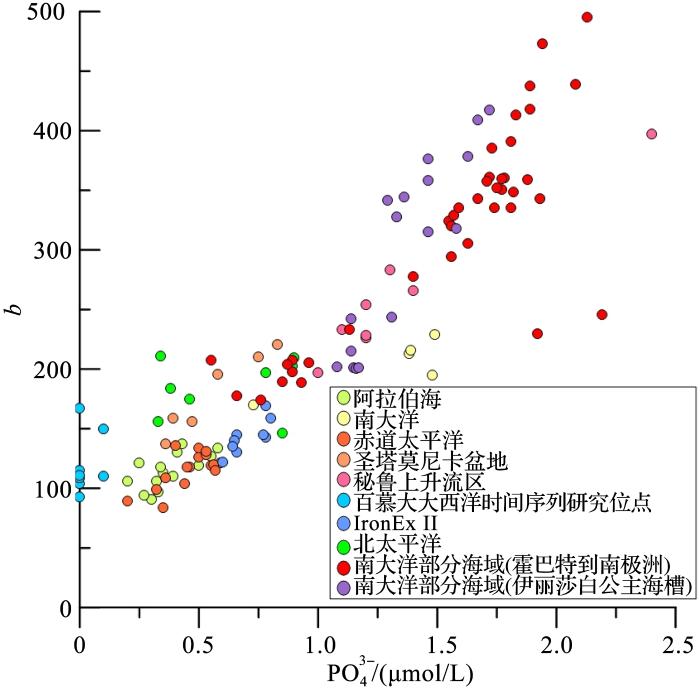
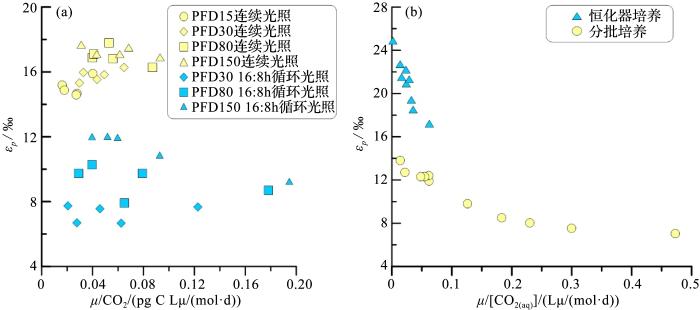
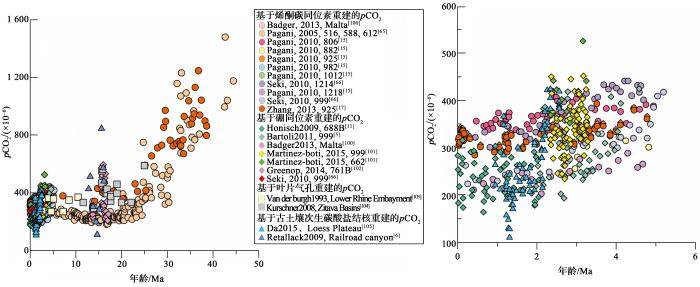

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}