Early Archean (3.3-Billion to 3.5-billion-year-old) microfossils from Warrawoona Group, Australia
1
1987
... 蓝细菌被认为是出现在地球上最早的生命之一.根据化石纪录,其大约起源于35亿前[1 ] ,虽然不能确定此时的蓝细菌是否具有光合细胞,但毫无疑问其是最早进行光合活动的微生物.作为重要的初级生产者,蓝细菌进行光合作用向环境提供氧气,对地球大气环境演变和此后的真核生物诞生起到了至关重要的作用[2 ] . ...
The evolutionary diversification of cyanobacteria: Molecular-phylogenetic and paleontological perspectives
5
2006
... 蓝细菌被认为是出现在地球上最早的生命之一.根据化石纪录,其大约起源于35亿前[1 ] ,虽然不能确定此时的蓝细菌是否具有光合细胞,但毫无疑问其是最早进行光合活动的微生物.作为重要的初级生产者,蓝细菌进行光合作用向环境提供氧气,对地球大气环境演变和此后的真核生物诞生起到了至关重要的作用[2 ] . ...
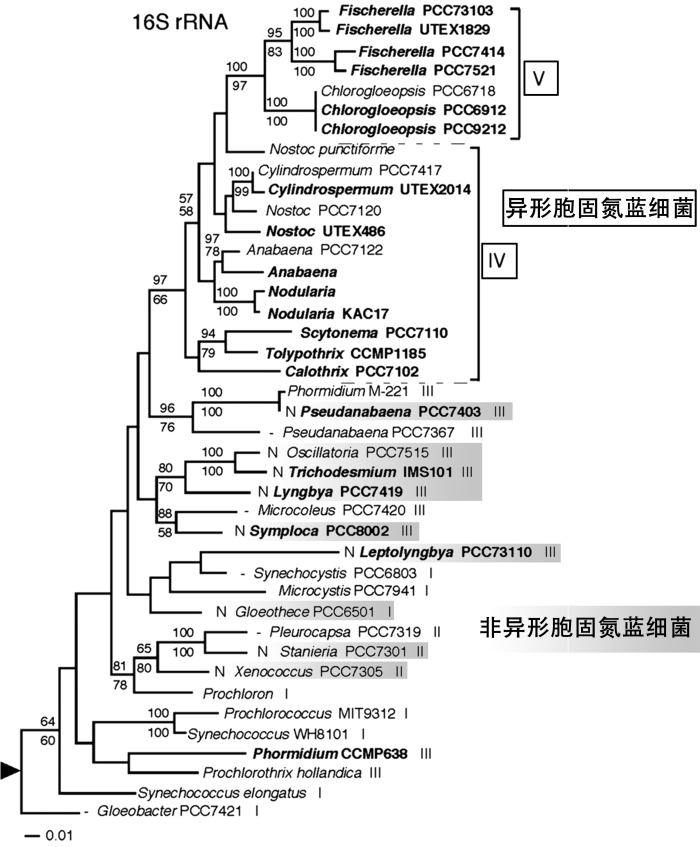
... 罗马数字I-V为蓝细菌形态学分类(Subsection I~V);黑色方框:异形胞固氮蓝细菌;N及灰色标识:非异形胞固氮蓝细菌;无标识:非固氮蓝细菌[2 ] ...
... Roman numerals denote cyanobacteria subsections I~V. Black box: Heterocystous diazotropic cyanobacteria; Grey shadow and N: Nonheterocystous diazotrophic cyanobacteria; Non-shaded: Non-diazotrophic cyanobacteria[2 ] ...
... 证据显示,蓝细菌异形细胞的分化出现在24.5亿~21.0亿年前[2 ] .在这一时期,大气氧气浓度增加,因此蓝细菌异形细胞的分化应该是为了应对大气中突增的氧气浓度[2 ,16 ] .与单细胞蓝细菌及非异形胞丝状蓝细菌不同,异形胞蓝细菌的16S rRNA序列在系统发育树上形成单源聚类(图1 )[5 ,17 ,18 ] . ...
... [2 ,16 ].与单细胞蓝细菌及非异形胞丝状蓝细菌不同,异形胞蓝细菌的16S rRNA序列在系统发育树上形成单源聚类(图1 )[5 ,17 ,18 ] . ...
Generic assignments, strain histories and properties of pure cultures of Cyanobacteria
1
1979
... 尽管蓝细菌又被称作蓝藻或蓝绿藻,但其为原核生物,属于格兰氏阴性菌.相较于其他细菌,蓝细菌有着多种形态结构.传统分类学根据蓝细菌生长形态,将蓝细菌分为5组(Section)[3 ] :组I,单细胞蓝细菌,以二分裂方式或出芽方式生殖,如其中的聚球藻属(Synechococcus );组II,单细胞蓝细菌,以多重分裂方式或结合二分裂方式生殖,如其中的小皮果蓝细菌属(Dermocarpella );组III,丝状蓝细菌,藻丝以营养细胞组成,单一面分裂,如其中的束毛藻属(Trichodesmium );组IV,丝状蓝细菌,藻丝中包含营养细胞以及在缺少氮营养的环境下分化出的异形细胞,单一面分裂,如其中的鱼腥藻属(Anabaena );组V,丝状蓝细菌,藻丝中包含营养细胞以及在缺少氮营养的环境下分化出的异形细胞,多重面分裂,如其中的费氏蓝细菌属(Fischerella) .但仅根据形态学分类过于简单,因为部分蓝细菌表现出相似的外形但亲缘较远,而有些虽然表现形态不同,但亲缘较近.因此之后结合系统发育学,更新了蓝细菌分类(图1 )[4 ,5 ] . ...
Recent changes (2008) in cyanobacteria taxonomy based on a combination of molecular background with phenotype and ecological consequences (genus and species concept)
1
2010
... 尽管蓝细菌又被称作蓝藻或蓝绿藻,但其为原核生物,属于格兰氏阴性菌.相较于其他细菌,蓝细菌有着多种形态结构.传统分类学根据蓝细菌生长形态,将蓝细菌分为5组(Section)[3 ] :组I,单细胞蓝细菌,以二分裂方式或出芽方式生殖,如其中的聚球藻属(Synechococcus );组II,单细胞蓝细菌,以多重分裂方式或结合二分裂方式生殖,如其中的小皮果蓝细菌属(Dermocarpella );组III,丝状蓝细菌,藻丝以营养细胞组成,单一面分裂,如其中的束毛藻属(Trichodesmium );组IV,丝状蓝细菌,藻丝中包含营养细胞以及在缺少氮营养的环境下分化出的异形细胞,单一面分裂,如其中的鱼腥藻属(Anabaena );组V,丝状蓝细菌,藻丝中包含营养细胞以及在缺少氮营养的环境下分化出的异形细胞,多重面分裂,如其中的费氏蓝细菌属(Fischerella) .但仅根据形态学分类过于简单,因为部分蓝细菌表现出相似的外形但亲缘较远,而有些虽然表现形态不同,但亲缘较近.因此之后结合系统发育学,更新了蓝细菌分类(图1 )[4 ,5 ] . ...
Bergey’s Manual of Systematics of Archaea and Bacteria
2
2015
... 尽管蓝细菌又被称作蓝藻或蓝绿藻,但其为原核生物,属于格兰氏阴性菌.相较于其他细菌,蓝细菌有着多种形态结构.传统分类学根据蓝细菌生长形态,将蓝细菌分为5组(Section)[3 ] :组I,单细胞蓝细菌,以二分裂方式或出芽方式生殖,如其中的聚球藻属(Synechococcus );组II,单细胞蓝细菌,以多重分裂方式或结合二分裂方式生殖,如其中的小皮果蓝细菌属(Dermocarpella );组III,丝状蓝细菌,藻丝以营养细胞组成,单一面分裂,如其中的束毛藻属(Trichodesmium );组IV,丝状蓝细菌,藻丝中包含营养细胞以及在缺少氮营养的环境下分化出的异形细胞,单一面分裂,如其中的鱼腥藻属(Anabaena );组V,丝状蓝细菌,藻丝中包含营养细胞以及在缺少氮营养的环境下分化出的异形细胞,多重面分裂,如其中的费氏蓝细菌属(Fischerella) .但仅根据形态学分类过于简单,因为部分蓝细菌表现出相似的外形但亲缘较远,而有些虽然表现形态不同,但亲缘较近.因此之后结合系统发育学,更新了蓝细菌分类(图1 )[4 ,5 ] . ...
... 证据显示,蓝细菌异形细胞的分化出现在24.5亿~21.0亿年前[2 ] .在这一时期,大气氧气浓度增加,因此蓝细菌异形细胞的分化应该是为了应对大气中突增的氧气浓度[2 ,16 ] .与单细胞蓝细菌及非异形胞丝状蓝细菌不同,异形胞蓝细菌的16S rRNA序列在系统发育树上形成单源聚类(图1 )[5 ,17 ,18 ] . ...
Long chain glycolipids with pentose head groups as biomarkers for marine endosymbiotic heterocystous cyanobacteria
4
2015
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
... 如前所述,C6 HGs由独立生存异形胞蓝细菌产生,通常生活于淡水或半咸水中,C5 HGs由与硅藻共生异形胞蓝细菌产生,分布于海洋中.Bale等[6 ] 在亚马孙陆架表层水体中检测到大量C5 HGs,但未检测到C6 HGs;Bauersachs等[59 ] 也提到未在现代海洋沉积物中检测到HGs(即C6 HGs).基于以上报道,我们最近研究发现,根据C6 HGs与C5 HGs含量比例,可以指示长江口—东海中陆源有机质输入情况,据此提出了一个新指标FHG [60 ] .我们的进一步工作还发现FHG 同样可以较好地应用于指示珠江口—南海区域的陆源有机质输入情况,并且在长江口外4 m长沉积柱中与BIT(Branched and Isoprenoid Tetraether)变化趋势较为一致,可以运用于长江口陆源有机质输入的历史重建中. ...
... HGs沉积记录能够为我们提供丰富而新颖的信息.首先,目前仅在异形胞蓝细菌膜脂中检测到HGs的存在,相较于其他常用的古环境生标来说,HGs最明显的优势在于有确定的生物来源以及在不同种属间的某些分布差异.例如C6 HG32 triol与其相应酮—二醇异形胞糖脂目前只发现于Stigonematales目蓝细菌菌株中.而此类蓝细菌通常生活于热带及亚热带淡水或地表环境中,因此C6 HG32 triol及keto-ol在湖相沉积物中的出现或许可以反映陆地古湖泊和古气候的某些信息.其次,结合异形胞蓝细菌固氮功能,HGs确定的生源降低了追踪古环境固氮活动的难度.一直以来,对于固氮活动的追踪是通过沉积物有机质氮同位素,但是同位素信息会受多种因素的干扰,具有多解性,不是固氮作用的确切指标.因此,C6 HGs与C5 HGs含量记录,结合δ15 N信息,可以分别用来重建湖泊及海洋历史固氮情况.第三,对于海洋沉积物中的C5 HGs,除能用于指示固氮信息外,还能反映固氮蓝细菌对海洋碳汇的贡献.这是因为生产C5 HGs的固氮蓝细菌通常与硅藻共生,而硅藻较重的硅质细胞在水体中会携带C5 HGs沉入海底,把蓝细菌固定的碳也能快速输入到深海,起到有效的生物碳泵作用.上层海水分层情况下往往存在大量硅藻—异形胞蓝细菌共生体,异形胞蓝细菌固氮提供营养导致硅藻的勃发.例如在北太平洋副热带环流分层水体中就存在这种勃发,虽然持续时间只有几天,但其能够快速沉降,构成当地总POC年通量的20%,对全球变暧起到重要的负反馈作用[67 ] .在过去的报道中,C5 HGs在3 000 m水深处被检测到[6 ,68 ] ,说明其可以有效地被输出至深海和沉积物中.据此,可以实现对海洋异形胞蓝细菌活动及其固碳与储碳过程的追踪,重建有关的碳氮生物地球化学过程. ...
Lipid biomarker signatures as tracers for harmful cyanobacterial blooms in the Baltic Sea
2
2017
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
... Bauersachs等[7 ] 提出可以用包括HGs等作为有害蓝细菌的生标化合物,追踪波罗的海地质历史上的蓝细菌勃发情况,并给出了几株异形胞蓝细菌分布的比例情况.同年,Sollai等[58 ] 利用HGs在波罗的海沉积柱的分布情况反演了波罗的海地质史上异形胞蓝细菌群落变化,发现其与波罗的海水文环境变化相对应.该研究涉及到全世新以来的样品.结合HGs分布的主成分分析(Principal Component Analysis,PCA)等数据,分析了波罗的海不同水文环境时期演变,包括早期淡水期、海水侵入期及完全半咸水期(图5 ).在早期淡水时期,(C6 ) HG26 diol与(C6 ) HG28 diol占主导,反映出此时期以例如Anabaena 及Aphanizomenon 为优势异形胞蓝细菌;在距今7.2~3.5 ka,波罗的海由淡水完全转变为半咸水后,(C6 ) HG26 diol逐渐占主导,反映出以Nostocaceae 科蓝细菌为优势菌的分布情况. ...
Diazotrophic microbial community of coastal microbial mats of the southern North Sea
2
2011
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
... Bauersachs等[8 ] 探究了荷兰北海堰洲岛Schiermonnikoog潮汐带中固氮微生物、HGs以及δ15 N的分布情况.他们发现在潮上带检测出异形胞蓝细菌的存在,并且发现了大量HGs(如C6 HG26 diol和C6 HG28 triol),同时相较于潮间带,潮上带表现出更负的δ15 N(潮间带、潮上带δ15 N变化范围分别为2‰~6‰和-1.2‰~1‰).异形胞蓝细菌、HGs与δ15 N之间良好的对应性首次证明了HGs在运用于指示固氮活动的可靠性. ...
Cyanobacterial heterocyst glycolipids in cultures and environmental samples: Diversity and biomarker potential
4
2012
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
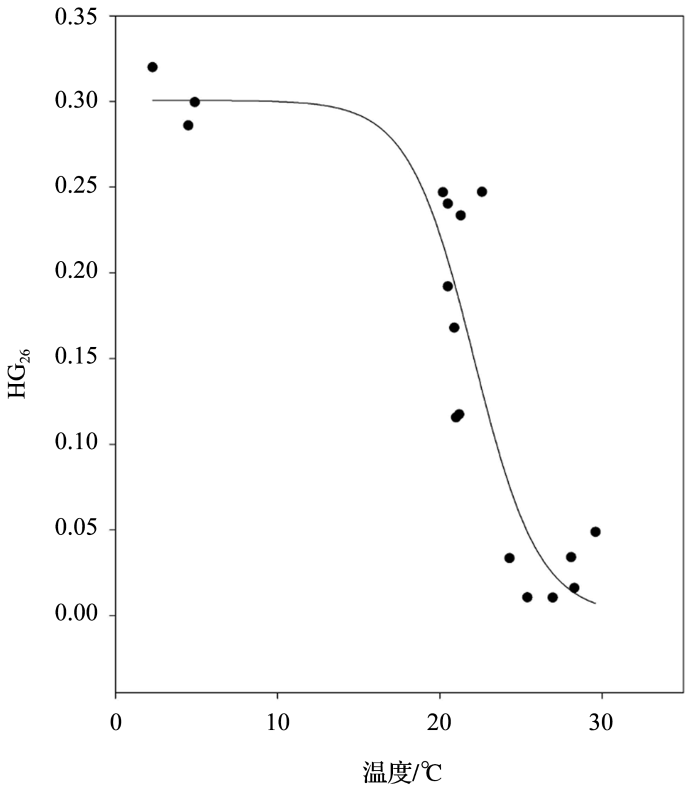
... 这种细胞膜内脂类化合物结构相对比例变化,或许是异形细胞为优化气体跨膜扩散而产生的生理适应.即温度升高,呼吸加强,氧气消耗速率增加,异形细胞对抑制气体向胞内扩散的需求降低.2012年,Wörmer等[9 ] 在2处极地生态环境及12处西班牙淡水样品中分析HGs分布时,发现HG26 指标确实会受环境温度的影响.但HG26 指标与环境温度关系呈现一个“S”型曲线,在温度高于25 ℃或低于15 ℃时,其指示温度变化的能力基本消失(图4 ). ...
... [
9 ]
HG<sub>26</sub> index vs. temperature in Spanish freshwaters and polar environment samples and fitted sigmoidal curve<sup>[<xref ref-type="bibr" rid="R9">9</xref>]</sup> Fig.4 ![]()
2014年Bauersachs等[56 ] 对HG26 (其文中写作HGI26 )指数与温度关系作了进一步探讨,并发现酮—醇结构HGs同分异构的比例会随着温度变化而变化.他们在文章中另外提出了基于HG28 diol与其相应的酮—醇HGs的指标(HGI28 ),以及基于同分异构HGs的指标等,并建立了温度与HGI26 和HGI28 等的换算关系式,但不同菌株呈现出不同的T-HGI26或28 关系.之后,Bauersachs等[57 ] 在德国淡水湖Lake Schreventeich采集水颗粒物及表层沉积物对HG26 指示温度作进一步分析验证,重新定义HDI(Heterocyst Diol Index)为diol-HGs占其相关keto-ol-HGs的比例,使HDI与温度正相关,又提出基于C6 HG28 triol及其相应酮—醇异形胞糖脂指标(HTI28 ).他们通过双尾t检验计算验证得到3个HGs指标与表层水温度的显著相关性(r 2 >0.70,p <0.001),同时也发现HGs指标与水体溶解氧含量或pH没有或者仅有微弱相关性.这是首个在自然环境中验证HGs温度指标的报告. ...
... [
9 ]
Fig.4 ![]()
2014年Bauersachs等[56 ] 对HG26 (其文中写作HGI26 )指数与温度关系作了进一步探讨,并发现酮—醇结构HGs同分异构的比例会随着温度变化而变化.他们在文章中另外提出了基于HG28 diol与其相应的酮—醇HGs的指标(HGI28 ),以及基于同分异构HGs的指标等,并建立了温度与HGI26 和HGI28 等的换算关系式,但不同菌株呈现出不同的T-HGI26或28 关系.之后,Bauersachs等[57 ] 在德国淡水湖Lake Schreventeich采集水颗粒物及表层沉积物对HG26 指示温度作进一步分析验证,重新定义HDI(Heterocyst Diol Index)为diol-HGs占其相关keto-ol-HGs的比例,使HDI与温度正相关,又提出基于C6 HG28 triol及其相应酮—醇异形胞糖脂指标(HTI28 ).他们通过双尾t检验计算验证得到3个HGs指标与表层水温度的显著相关性(r 2 >0.70,p <0.001),同时也发现HGs指标与水体溶解氧含量或pH没有或者仅有微弱相关性.这是首个在自然环境中验证HGs温度指标的报告. ...
Reconciling the incompatible: N2 fixation and O2
1
2006
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
N2 fixation in phototrophs: Adaptation to a specialized way of life
1
2001
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
Nitrogen fixation and photosynthetic oxygen evolution in cyanobacteria
1
2003
... 许多蓝细菌是重要的固氮微生物,广泛分布于淡水[6 ] 、半咸水[7 ] 、海洋[8 ] 、极地[9 ] 及土壤环境中.生物固氮是微生物将分子态氮转化为有机胺的过程.在这一过程中打破N2 分子内稳定的三键需要极高的能量,因此生物固氮活动需要固氮酶的催化.但是光合产氧活动与固氮活动理论上是不兼容的,因为固氮酶活性会受到氧气的抑制,暴露在氧气环境下,固氮酶会迅速且永久失活[10 ] .因此蓝细菌进化出不同的策略,以保证在进行产氧光合作用的同时,能够给固氮酶提供无氧环境[11 ,12 ] .其中一个策略就是将光合活动与固氮活动在空间上分开,蓝细菌会分化出异形细胞以阻止氮气进入此细胞内,异形细胞便作为固氮场所. ...
Nitrogen fixation by marine cyanobacteria
1
2010
... 蓝细菌是海洋中重要的固氮微生物,为海洋生态系统提供营养盐,平衡海洋生态系统的氮损失.目前已知海洋固氮蓝细菌大致分成3类[13 ] :丝状非异形胞蓝细菌Trichodesmium 、异形胞蓝细菌(如Richelia 和Calothrix 等)以及单细胞蓝细菌(如Crocosphaera ).其中,异形胞蓝细菌通常与硅藻共生,其固氮贡献在某些区域十分可观.如在加利福尼亚海湾,共生异形胞蓝细菌R. intraxellularis 固氮量可达35%~48%[14 ] ;在热带北大西洋海域,共生异形胞蓝细菌为表层水提供近70%的氮需求[15 ] . ...
Summer surface waters in the Gulf of California: Prime habitat for biological N2 fixation
1
2007
... 蓝细菌是海洋中重要的固氮微生物,为海洋生态系统提供营养盐,平衡海洋生态系统的氮损失.目前已知海洋固氮蓝细菌大致分成3类[13 ] :丝状非异形胞蓝细菌Trichodesmium 、异形胞蓝细菌(如Richelia 和Calothrix 等)以及单细胞蓝细菌(如Crocosphaera ).其中,异形胞蓝细菌通常与硅藻共生,其固氮贡献在某些区域十分可观.如在加利福尼亚海湾,共生异形胞蓝细菌R. intraxellularis 固氮量可达35%~48%[14 ] ;在热带北大西洋海域,共生异形胞蓝细菌为表层水提供近70%的氮需求[15 ] . ...
Extensive bloom of a N2 -fixing diatom/cyanobacterial association in the tropical Atlantic Ocean
1
1999
... 蓝细菌是海洋中重要的固氮微生物,为海洋生态系统提供营养盐,平衡海洋生态系统的氮损失.目前已知海洋固氮蓝细菌大致分成3类[13 ] :丝状非异形胞蓝细菌Trichodesmium 、异形胞蓝细菌(如Richelia 和Calothrix 等)以及单细胞蓝细菌(如Crocosphaera ).其中,异形胞蓝细菌通常与硅藻共生,其固氮贡献在某些区域十分可观.如在加利福尼亚海湾,共生异形胞蓝细菌R. intraxellularis 固氮量可达35%~48%[14 ] ;在热带北大西洋海域,共生异形胞蓝细菌为表层水提供近70%的氮需求[15 ] . ...
Heterocyst formation in Anaebena
1
2000
... 证据显示,蓝细菌异形细胞的分化出现在24.5亿~21.0亿年前[2 ] .在这一时期,大气氧气浓度增加,因此蓝细菌异形细胞的分化应该是为了应对大气中突增的氧气浓度[2 ,16 ] .与单细胞蓝细菌及非异形胞丝状蓝细菌不同,异形胞蓝细菌的16S rRNA序列在系统发育树上形成单源聚类(图1 )[5 ,17 ,18 ] . ...
Evolutionary relationships among cyanobacteria and green chloroplasts
1
1988
... 证据显示,蓝细菌异形细胞的分化出现在24.5亿~21.0亿年前[2 ] .在这一时期,大气氧气浓度增加,因此蓝细菌异形细胞的分化应该是为了应对大气中突增的氧气浓度[2 ,16 ] .与单细胞蓝细菌及非异形胞丝状蓝细菌不同,异形胞蓝细菌的16S rRNA序列在系统发育树上形成单源聚类(图1 )[5 ,17 ,18 ] . ...
rDNA analyses of planktonic heterocystous cyanobacteria, including members of the genera Anabaenopsis and Cyanospira
1
2002
... 证据显示,蓝细菌异形细胞的分化出现在24.5亿~21.0亿年前[2 ] .在这一时期,大气氧气浓度增加,因此蓝细菌异形细胞的分化应该是为了应对大气中突增的氧气浓度[2 ,16 ] .与单细胞蓝细菌及非异形胞丝状蓝细菌不同,异形胞蓝细菌的16S rRNA序列在系统发育树上形成单源聚类(图1 )[5 ,17 ,18 ] . ...
Endosymbiotic heterocystous cyanobacteria synthesize different heterocyst glycolipids than free-living heterocystous cyanobacteria
4
2013
... 异形胞蓝细菌以2种形式生存.一种是独立生活种类,例如Nostocales及Stigonematales目蓝细菌菌株[19 ] ,它们广泛生长于各气候带的淡水或半咸水中[20 ] .而在海洋中,独立生活的异形胞蓝细菌比较罕见,它们通常以与硅藻体内或体外共生的形式生存.如异形胞蓝细菌Richelia 常被发现与根管藻(Rhizosolenia )、半管藻(Hemiaulus )及几内亚藻(Guinardia )等硅藻体内共生,而Calothrix 常附着在角毛藻(Chatoceros )或幅杆藻(Bacteriastrum )上,以体外共生形式生存[21 ,22 ] .在共生体内,异形胞蓝细菌为宿主提供生长所需的氮营养[23 ] ,使得硅藻可以生长在寡营养海域.共生异形胞蓝细菌是热带及亚热带寡营养海区的重要固氮微生物. ...
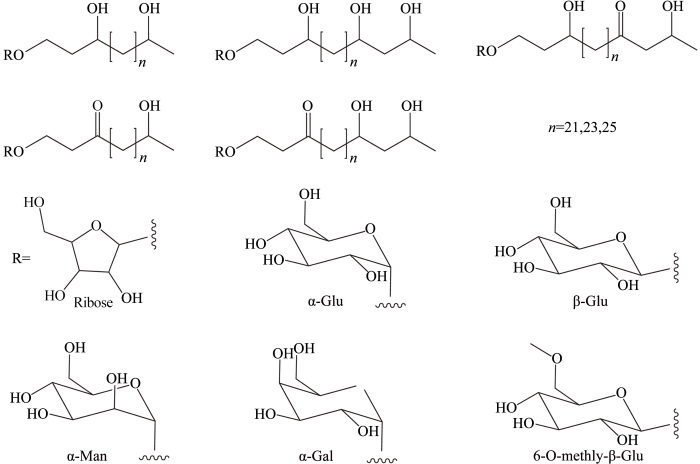
... HGs由糖苷基连接长链二醇、三醇、酮醇或酮二醇烷基组成.糖基头通常为六碳糖苷(C
6 ),最近又发现了五碳糖苷(C
5 ).C
6 糖大多为葡萄糖,同时也存在少数半乳糖
[48 ] 或甘露糖
[49 ] .最新的研究发现
Calothrix 异形胞糖脂C
6 糖基头C-6位置连有一甲基
[45 ] .烷基烃链一般含有26~32个碳原子,羟基或羰基位于烃链上C-3,ω-1及ω-3位置(
图3 ).
图3 <strong>HGs</strong>结构图<sup>[<xref ref-type="bibr" rid="R19">19</xref>,<xref ref-type="bibr" rid="R45">45</xref>,<xref ref-type="bibr" rid="R49">49</xref>]</sup> HGs structures<sup>[<xref ref-type="bibr" rid="R19">19</xref>,<xref ref-type="bibr" rid="R45">45</xref>,<xref ref-type="bibr" rid="R49">49</xref>]</sup> Fig.3 ![]()
对HGs的缩写命名没有统一规定,目前其缩写先后标明糖苷基头碳原子数、烷烃链碳原子数及烷烃链官能团信息.在C5 HGs被发现之前,C6 HGs的缩写一般仅包括烷烃链碳原子数及烷烃链官能团信息,即Cn n 代表烷烃链碳原子数.如C26 HG diol指代由六碳糖苷与含二十六碳的二醇连接形成的糖脂.随着C5 HGs被解析出来后,HGs的缩写要指明是六碳或五碳糖基头,烷烃链碳原子数则以下标标在HG字母后,即Cm n m 代表糖基头碳原子数,n 代表烷烃链碳原子数.如C5 HG30 triol指代由五碳糖苷与含三十碳的三醇连接形成的糖脂,而上述C26 HG diol则改为C6 HG26 diol.但是上述命名不包括甲基化HGs,并且HGs化合物的缩写命名并不能体现出烷烃链上官能团所在位置,也不能体现出糖苷基头的结构类型. ...
... [
19 ,
45 ,
49 ]
Fig.3 ![]()
对HGs的缩写命名没有统一规定,目前其缩写先后标明糖苷基头碳原子数、烷烃链碳原子数及烷烃链官能团信息.在C5 HGs被发现之前,C6 HGs的缩写一般仅包括烷烃链碳原子数及烷烃链官能团信息,即Cn n 代表烷烃链碳原子数.如C26 HG diol指代由六碳糖苷与含二十六碳的二醇连接形成的糖脂.随着C5 HGs被解析出来后,HGs的缩写要指明是六碳或五碳糖基头,烷烃链碳原子数则以下标标在HG字母后,即Cm n m 代表糖基头碳原子数,n 代表烷烃链碳原子数.如C5 HG30 triol指代由五碳糖苷与含三十碳的三醇连接形成的糖脂,而上述C26 HG diol则改为C6 HG26 diol.但是上述命名不包括甲基化HGs,并且HGs化合物的缩写命名并不能体现出烷烃链上官能团所在位置,也不能体现出糖苷基头的结构类型. ...
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
Is the distribution of nitrogen-fixing cyanobacteria in the oceans related to temperature?
1
2009
... 异形胞蓝细菌以2种形式生存.一种是独立生活种类,例如Nostocales及Stigonematales目蓝细菌菌株[19 ] ,它们广泛生长于各气候带的淡水或半咸水中[20 ] .而在海洋中,独立生活的异形胞蓝细菌比较罕见,它们通常以与硅藻体内或体外共生的形式生存.如异形胞蓝细菌Richelia 常被发现与根管藻(Rhizosolenia )、半管藻(Hemiaulus )及几内亚藻(Guinardia )等硅藻体内共生,而Calothrix 常附着在角毛藻(Chatoceros )或幅杆藻(Bacteriastrum )上,以体外共生形式生存[21 ,22 ] .在共生体内,异形胞蓝细菌为宿主提供生长所需的氮营养[23 ] ,使得硅藻可以生长在寡营养海域.共生异形胞蓝细菌是热带及亚热带寡营养海区的重要固氮微生物. ...
Nitrogen-fixation by the cyanobacterial symbiont of the diatom genus Hemiaulus
1
1991
... 异形胞蓝细菌以2种形式生存.一种是独立生活种类,例如Nostocales及Stigonematales目蓝细菌菌株[19 ] ,它们广泛生长于各气候带的淡水或半咸水中[20 ] .而在海洋中,独立生活的异形胞蓝细菌比较罕见,它们通常以与硅藻体内或体外共生的形式生存.如异形胞蓝细菌Richelia 常被发现与根管藻(Rhizosolenia )、半管藻(Hemiaulus )及几内亚藻(Guinardia )等硅藻体内共生,而Calothrix 常附着在角毛藻(Chatoceros )或幅杆藻(Bacteriastrum )上,以体外共生形式生存[21 ,22 ] .在共生体内,异形胞蓝细菌为宿主提供生长所需的氮营养[23 ] ,使得硅藻可以生长在寡营养海域.共生异形胞蓝细菌是热带及亚热带寡营养海区的重要固氮微生物. ...
Marine Nitrogen-Fixing Diatom-Cyanobacteria Symbioses
1
1992
... 异形胞蓝细菌以2种形式生存.一种是独立生活种类,例如Nostocales及Stigonematales目蓝细菌菌株[19 ] ,它们广泛生长于各气候带的淡水或半咸水中[20 ] .而在海洋中,独立生活的异形胞蓝细菌比较罕见,它们通常以与硅藻体内或体外共生的形式生存.如异形胞蓝细菌Richelia 常被发现与根管藻(Rhizosolenia )、半管藻(Hemiaulus )及几内亚藻(Guinardia )等硅藻体内共生,而Calothrix 常附着在角毛藻(Chatoceros )或幅杆藻(Bacteriastrum )上,以体外共生形式生存[21 ,22 ] .在共生体内,异形胞蓝细菌为宿主提供生长所需的氮营养[23 ] ,使得硅藻可以生长在寡营养海域.共生异形胞蓝细菌是热带及亚热带寡营养海区的重要固氮微生物. ...
Nitrogen fixation and transfer in open ocean diatom-cyanobacterial symbioses
1
2011
... 异形胞蓝细菌以2种形式生存.一种是独立生活种类,例如Nostocales及Stigonematales目蓝细菌菌株[19 ] ,它们广泛生长于各气候带的淡水或半咸水中[20 ] .而在海洋中,独立生活的异形胞蓝细菌比较罕见,它们通常以与硅藻体内或体外共生的形式生存.如异形胞蓝细菌Richelia 常被发现与根管藻(Rhizosolenia )、半管藻(Hemiaulus )及几内亚藻(Guinardia )等硅藻体内共生,而Calothrix 常附着在角毛藻(Chatoceros )或幅杆藻(Bacteriastrum )上,以体外共生形式生存[21 ,22 ] .在共生体内,异形胞蓝细菌为宿主提供生长所需的氮营养[23 ] ,使得硅藻可以生长在寡营养海域.共生异形胞蓝细菌是热带及亚热带寡营养海区的重要固氮微生物. ...
Cyanobacterial heterocysts
4
2010
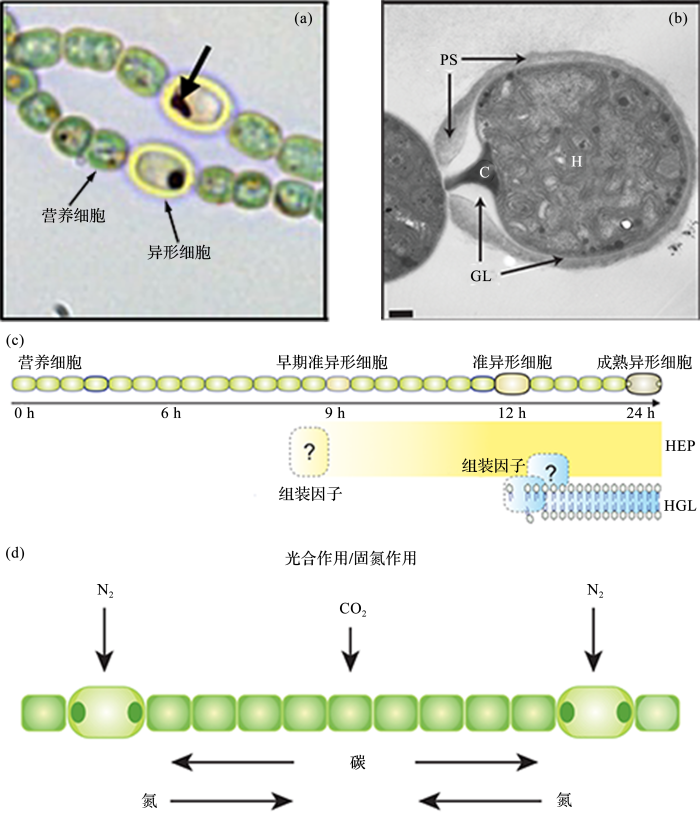
... 异形细胞在外形上与正常营养细胞有着明显差异,其细胞体形更大,形状更圆,色素含量低且细胞壁更厚(图2 a)[24 ,25 ,26 ] .异形细胞细胞壁有两层膜,其中内膜由异形胞糖脂(Heterocyst Glycolipids,HGs)组成(图2 b)[27 ] .异形细胞的分化保证了蓝细菌在光合活动的同时也可以进行固氮活动,一方面,其厚厚的细胞壁可以阻止氧气进入,创造出适合固氮酶作用的微氧环境;另一方面,异形细胞保持高呼吸速率以消耗胞内的微量氧气[28 ] . ...
... (a) Anavaena 藻丝[26 ] ;(b) 异形细胞透射电摬图[24 ] ;(c) 异形细胞分化示意图[27 ] ;(d) 异形细胞与营养细胞物质交换示意图[34 ] ;GL, HGL:糖脂层;PS, HEP:多聚糖层;C:极性藻青藻颗粒;H:异形细胞 ...
... (a) Filament of Anabaena [26 ] ;(b) Transmission electron micrograph of hetercyst[24 ] ; (c) Heterocyst formation diagram[27 ] ; (d) Transport of metabolites between heterocyst and vegetative cell[34 ] . GL, HGL: Glycolipids layer; PS, HEP: Polysaccharide layer; C: Polar cyanophycin granule; H: Heterocyst ...
... 并非在所有环境下异形胞蓝细菌均会分化出异形细胞.当在氮营养充足的情况下,为避免固氮活动消耗大量的能量[29 ] ,它们并不会分化出异形细胞.只有当异形胞蓝细菌感受到环境中氮营养短缺,并且成熟异形细胞的固氮活动已经达到极限时,新的异形细胞才开始分化[24 ,30 ] .异形细胞发育要在约24 h内完成(图2 c),这一期间涉及到营养细胞完成分化至异形细胞,并且在此过程中要尽量减少其余营养细胞体内基因表达和新陈代谢的变化.如果在异形细胞开始发育后的9~12 h内加入氮营养盐或PatS五肽时,异形细胞的分化呈现出可逆状态;过了这一时间点后,营养细胞将最终完成异形细胞的分化[31 ,32 ] .异形细胞分化的中后阶段表现出结构及生理改变(图2 c),在这一阶段,异形细胞的细胞壁开始覆盖上两层,内层为糖脂层——用以阻止氧气进入,外层为多聚糖层——用以保护脆性糖脂层[27 ] . ...
Pigments and lipids of heterocysts
1
1969
... 异形细胞在外形上与正常营养细胞有着明显差异,其细胞体形更大,形状更圆,色素含量低且细胞壁更厚(图2 a)[24 ,25 ,26 ] .异形细胞细胞壁有两层膜,其中内膜由异形胞糖脂(Heterocyst Glycolipids,HGs)组成(图2 b)[27 ] .异形细胞的分化保证了蓝细菌在光合活动的同时也可以进行固氮活动,一方面,其厚厚的细胞壁可以阻止氧气进入,创造出适合固氮酶作用的微氧环境;另一方面,异形细胞保持高呼吸速率以消耗胞内的微量氧气[28 ] . ...
Glycolipid composition of the heterocyst envelope of Anabaena sp. PCC 7120 is crucial for diazotrophic growth and relies on the UDP-galactose 4-epimerase HgdA
3
811
... 异形细胞在外形上与正常营养细胞有着明显差异,其细胞体形更大,形状更圆,色素含量低且细胞壁更厚(图2 a)[24 ,25 ,26 ] .异形细胞细胞壁有两层膜,其中内膜由异形胞糖脂(Heterocyst Glycolipids,HGs)组成(图2 b)[27 ] .异形细胞的分化保证了蓝细菌在光合活动的同时也可以进行固氮活动,一方面,其厚厚的细胞壁可以阻止氧气进入,创造出适合固氮酶作用的微氧环境;另一方面,异形细胞保持高呼吸速率以消耗胞内的微量氧气[28 ] . ...
... (a) Anavaena 藻丝[26 ] ;(b) 异形细胞透射电摬图[24 ] ;(c) 异形细胞分化示意图[27 ] ;(d) 异形细胞与营养细胞物质交换示意图[34 ] ;GL, HGL:糖脂层;PS, HEP:多聚糖层;C:极性藻青藻颗粒;H:异形细胞 ...
... (a) Filament of Anabaena [26 ] ;(b) Transmission electron micrograph of hetercyst[24 ] ; (c) Heterocyst formation diagram[27 ] ; (d) Transport of metabolites between heterocyst and vegetative cell[34 ] . GL, HGL: Glycolipids layer; PS, HEP: Polysaccharide layer; C: Polar cyanophycin granule; H: Heterocyst ...
The cell wall in heterocyst formation by Anabaena sp. PCC 7120 From vegetative cells to heterocysts
4
2009
... 异形细胞在外形上与正常营养细胞有着明显差异,其细胞体形更大,形状更圆,色素含量低且细胞壁更厚(图2 a)[24 ,25 ,26 ] .异形细胞细胞壁有两层膜,其中内膜由异形胞糖脂(Heterocyst Glycolipids,HGs)组成(图2 b)[27 ] .异形细胞的分化保证了蓝细菌在光合活动的同时也可以进行固氮活动,一方面,其厚厚的细胞壁可以阻止氧气进入,创造出适合固氮酶作用的微氧环境;另一方面,异形细胞保持高呼吸速率以消耗胞内的微量氧气[28 ] . ...
... (a) Anavaena 藻丝[26 ] ;(b) 异形细胞透射电摬图[24 ] ;(c) 异形细胞分化示意图[27 ] ;(d) 异形细胞与营养细胞物质交换示意图[34 ] ;GL, HGL:糖脂层;PS, HEP:多聚糖层;C:极性藻青藻颗粒;H:异形细胞 ...
... (a) Filament of Anabaena [26 ] ;(b) Transmission electron micrograph of hetercyst[24 ] ; (c) Heterocyst formation diagram[27 ] ; (d) Transport of metabolites between heterocyst and vegetative cell[34 ] . GL, HGL: Glycolipids layer; PS, HEP: Polysaccharide layer; C: Polar cyanophycin granule; H: Heterocyst ...
... 并非在所有环境下异形胞蓝细菌均会分化出异形细胞.当在氮营养充足的情况下,为避免固氮活动消耗大量的能量[29 ] ,它们并不会分化出异形细胞.只有当异形胞蓝细菌感受到环境中氮营养短缺,并且成熟异形细胞的固氮活动已经达到极限时,新的异形细胞才开始分化[24 ,30 ] .异形细胞发育要在约24 h内完成(图2 c),这一期间涉及到营养细胞完成分化至异形细胞,并且在此过程中要尽量减少其余营养细胞体内基因表达和新陈代谢的变化.如果在异形细胞开始发育后的9~12 h内加入氮营养盐或PatS五肽时,异形细胞的分化呈现出可逆状态;过了这一时间点后,营养细胞将最终完成异形细胞的分化[31 ,32 ] .异形细胞分化的中后阶段表现出结构及生理改变(图2 c),在这一阶段,异形细胞的细胞壁开始覆盖上两层,内层为糖脂层——用以阻止氧气进入,外层为多聚糖层——用以保护脆性糖脂层[27 ] . ...
Heterocyst metabolism and development
1
1994
... 异形细胞在外形上与正常营养细胞有着明显差异,其细胞体形更大,形状更圆,色素含量低且细胞壁更厚(图2 a)[24 ,25 ,26 ] .异形细胞细胞壁有两层膜,其中内膜由异形胞糖脂(Heterocyst Glycolipids,HGs)组成(图2 b)[27 ] .异形细胞的分化保证了蓝细菌在光合活动的同时也可以进行固氮活动,一方面,其厚厚的细胞壁可以阻止氧气进入,创造出适合固氮酶作用的微氧环境;另一方面,异形细胞保持高呼吸速率以消耗胞内的微量氧气[28 ] . ...
Isotopic Tracers of the marine nitrogen cycle: Present and past
1
2006
... 并非在所有环境下异形胞蓝细菌均会分化出异形细胞.当在氮营养充足的情况下,为避免固氮活动消耗大量的能量[29 ] ,它们并不会分化出异形细胞.只有当异形胞蓝细菌感受到环境中氮营养短缺,并且成熟异形细胞的固氮活动已经达到极限时,新的异形细胞才开始分化[24 ,30 ] .异形细胞发育要在约24 h内完成(图2 c),这一期间涉及到营养细胞完成分化至异形细胞,并且在此过程中要尽量减少其余营养细胞体内基因表达和新陈代谢的变化.如果在异形细胞开始发育后的9~12 h内加入氮营养盐或PatS五肽时,异形细胞的分化呈现出可逆状态;过了这一时间点后,营养细胞将最终完成异形细胞的分化[31 ,32 ] .异形细胞分化的中后阶段表现出结构及生理改变(图2 c),在这一阶段,异形细胞的细胞壁开始覆盖上两层,内层为糖脂层——用以阻止氧气进入,外层为多聚糖层——用以保护脆性糖脂层[27 ] . ...
Heterocyst differentiation and pattern formation in cyanobacteria: A chorus of signals
1
2006
... 并非在所有环境下异形胞蓝细菌均会分化出异形细胞.当在氮营养充足的情况下,为避免固氮活动消耗大量的能量[29 ] ,它们并不会分化出异形细胞.只有当异形胞蓝细菌感受到环境中氮营养短缺,并且成熟异形细胞的固氮活动已经达到极限时,新的异形细胞才开始分化[24 ,30 ] .异形细胞发育要在约24 h内完成(图2 c),这一期间涉及到营养细胞完成分化至异形细胞,并且在此过程中要尽量减少其余营养细胞体内基因表达和新陈代谢的变化.如果在异形细胞开始发育后的9~12 h内加入氮营养盐或PatS五肽时,异形细胞的分化呈现出可逆状态;过了这一时间点后,营养细胞将最终完成异形细胞的分化[31 ,32 ] .异形细胞分化的中后阶段表现出结构及生理改变(图2 c),在这一阶段,异形细胞的细胞壁开始覆盖上两层,内层为糖脂层——用以阻止氧气进入,外层为多聚糖层——用以保护脆性糖脂层[27 ] . ...
PatS and products of nitrogen fixation control heterocyst pattern
1
2001
... 并非在所有环境下异形胞蓝细菌均会分化出异形细胞.当在氮营养充足的情况下,为避免固氮活动消耗大量的能量[29 ] ,它们并不会分化出异形细胞.只有当异形胞蓝细菌感受到环境中氮营养短缺,并且成熟异形细胞的固氮活动已经达到极限时,新的异形细胞才开始分化[24 ,30 ] .异形细胞发育要在约24 h内完成(图2 c),这一期间涉及到营养细胞完成分化至异形细胞,并且在此过程中要尽量减少其余营养细胞体内基因表达和新陈代谢的变化.如果在异形细胞开始发育后的9~12 h内加入氮营养盐或PatS五肽时,异形细胞的分化呈现出可逆状态;过了这一时间点后,营养细胞将最终完成异形细胞的分化[31 ,32 ] .异形细胞分化的中后阶段表现出结构及生理改变(图2 c),在这一阶段,异形细胞的细胞壁开始覆盖上两层,内层为糖脂层——用以阻止氧气进入,外层为多聚糖层——用以保护脆性糖脂层[27 ] . ...
Effect on heterocyst differentiation of nitrogen fixation in vegetative cells of the Cyanobacterium Anabaena variabilis ATCC 29413
1
2001
... 并非在所有环境下异形胞蓝细菌均会分化出异形细胞.当在氮营养充足的情况下,为避免固氮活动消耗大量的能量[29 ] ,它们并不会分化出异形细胞.只有当异形胞蓝细菌感受到环境中氮营养短缺,并且成熟异形细胞的固氮活动已经达到极限时,新的异形细胞才开始分化[24 ,30 ] .异形细胞发育要在约24 h内完成(图2 c),这一期间涉及到营养细胞完成分化至异形细胞,并且在此过程中要尽量减少其余营养细胞体内基因表达和新陈代谢的变化.如果在异形细胞开始发育后的9~12 h内加入氮营养盐或PatS五肽时,异形细胞的分化呈现出可逆状态;过了这一时间点后,营养细胞将最终完成异形细胞的分化[31 ,32 ] .异形细胞分化的中后阶段表现出结构及生理改变(图2 c),在这一阶段,异形细胞的细胞壁开始覆盖上两层,内层为糖脂层——用以阻止氧气进入,外层为多聚糖层——用以保护脆性糖脂层[27 ] . ...
Is the periplasm continuous in filamentous multicellular cyanobacteria?
1
2006
... 异形细胞与营养细胞相互依赖,他们之间的代谢物质与信号交换通过连续细胞周质或微小的胞间连丝进行传递(图2 d)[33 ,34 ] .异形细胞内缺乏光系统II(Photosystem II),不能进行光合作用[35 ] ,因此需要依靠营养细胞光合活动提供碳源.在异形胞蓝细菌Anabaena PCC 7120藻丝中,营养细胞会提供给异形细胞谷氨酸盐,以供后者合成谷氨酰胺及其他氨基酸[36~38] ;相反的,异形细胞会以氨基酸的形式提供给营养细胞生长所需要的氮营养[39 ] . ...
Heterocyst differentiation: From single mutants to global approaches
3
2012
... (a) Anavaena 藻丝[26 ] ;(b) 异形细胞透射电摬图[24 ] ;(c) 异形细胞分化示意图[27 ] ;(d) 异形细胞与营养细胞物质交换示意图[34 ] ;GL, HGL:糖脂层;PS, HEP:多聚糖层;C:极性藻青藻颗粒;H:异形细胞 ...
... (a) Filament of Anabaena [26 ] ;(b) Transmission electron micrograph of hetercyst[24 ] ; (c) Heterocyst formation diagram[27 ] ; (d) Transport of metabolites between heterocyst and vegetative cell[34 ] . GL, HGL: Glycolipids layer; PS, HEP: Polysaccharide layer; C: Polar cyanophycin granule; H: Heterocyst ...
... 异形细胞与营养细胞相互依赖,他们之间的代谢物质与信号交换通过连续细胞周质或微小的胞间连丝进行传递(图2 d)[33 ,34 ] .异形细胞内缺乏光系统II(Photosystem II),不能进行光合作用[35 ] ,因此需要依靠营养细胞光合活动提供碳源.在异形胞蓝细菌Anabaena PCC 7120藻丝中,营养细胞会提供给异形细胞谷氨酸盐,以供后者合成谷氨酰胺及其他氨基酸[36~38] ;相反的,异形细胞会以氨基酸的形式提供给营养细胞生长所需要的氮营养[39 ] . ...
Absence of photosystem 2 in heterocysts of the blue-green alga Anabaena
1
1972
... 异形细胞与营养细胞相互依赖,他们之间的代谢物质与信号交换通过连续细胞周质或微小的胞间连丝进行传递(图2 d)[33 ,34 ] .异形细胞内缺乏光系统II(Photosystem II),不能进行光合作用[35 ] ,因此需要依靠营养细胞光合活动提供碳源.在异形胞蓝细菌Anabaena PCC 7120藻丝中,营养细胞会提供给异形细胞谷氨酸盐,以供后者合成谷氨酰胺及其他氨基酸[36~38] ;相反的,异形细胞会以氨基酸的形式提供给营养细胞生长所需要的氮营养[39 ] . ...
Carbon cycling in Anabaena sp. PCC 7120. Sucrose synthesis in the heterocysts and possible role in Nitrogen Fixation
0
2007
Role of NtcA, a cyanobacterial global nitrogen regulator, in the regulation of sucrose metabolism gene expression in Anabaena sp. PCC 7120
0
2009
The GS-GOGAT pathway is not operative in the heterocysts. Cloning and expression of glsF gene from the cyanobacterium Anabaena sp. PCC 7120
0
2000
Regulation of cellular differentiation in filamentous cyanobacteria in free-living and plant-associated symbiotic growth states
1
2002
... 异形细胞与营养细胞相互依赖,他们之间的代谢物质与信号交换通过连续细胞周质或微小的胞间连丝进行传递(图2 d)[33 ,34 ] .异形细胞内缺乏光系统II(Photosystem II),不能进行光合作用[35 ] ,因此需要依靠营养细胞光合活动提供碳源.在异形胞蓝细菌Anabaena PCC 7120藻丝中,营养细胞会提供给异形细胞谷氨酸盐,以供后者合成谷氨酰胺及其他氨基酸[36~38] ;相反的,异形细胞会以氨基酸的形式提供给营养细胞生长所需要的氮营养[39 ] . ...
New glycolipid specific to Nitrogen-fixing blue-green algae
1
1968
... 1968年Nichols等[40 ] 在利用硅胶层析分析几株蓝细菌(如Anabaena cylindrical )极性脂时发现,固氮蓝细菌含有一种极性小于磷酯酰乙醇胺的脂类化合物,而这一化合物在非固氮蓝细菌及其他藻类中均未检测到.进一步分析发现,这一脂类化合物连有糖苷基头.随后的研究发现,这类糖脂在正常营养细胞中不存在,只产出于异形细胞[41 ] .目前,已对大约20株异形胞蓝细菌的糖脂进行了分析,其结构逐渐被解析及完善,并被命名为HGs[26,42~47] . ...
Lipid composition of heterocysts
1
1969
... 1968年Nichols等[40 ] 在利用硅胶层析分析几株蓝细菌(如Anabaena cylindrical )极性脂时发现,固氮蓝细菌含有一种极性小于磷酯酰乙醇胺的脂类化合物,而这一化合物在非固氮蓝细菌及其他藻类中均未检测到.进一步分析发现,这一脂类化合物连有糖苷基头.随后的研究发现,这类糖脂在正常营养细胞中不存在,只产出于异形细胞[41 ] .目前,已对大约20株异形胞蓝细菌的糖脂进行了分析,其结构逐渐被解析及完善,并被命名为HGs[26,42~47] . ...
Monohexoside derivatives of long-chain polyhydroxy alcohols: A novel class of glycolipid specific to heterocystous algae
0
1972
Structural studies on the glycolipids from the envelope of the heterocyst of Anabaena cylindrica
0
1973
Biosynthesis of the heterocyst glycolipids in the cyanobacterium Anabaena cylindrica
0
1995
A novel heterocyst glycolipid detected in a pelagic N2 -fixing cyanobacterium of the genus Calothrix
4
2018
... HGs由糖苷基连接长链二醇、三醇、酮醇或酮二醇烷基组成.糖基头通常为六碳糖苷(C6 ),最近又发现了五碳糖苷(C5 ).C6 糖大多为葡萄糖,同时也存在少数半乳糖[48 ] 或甘露糖[49 ] .最新的研究发现Calothrix 异形胞糖脂C6 糖基头C-6位置连有一甲基[45 ] .烷基烃链一般含有26~32个碳原子,羟基或羰基位于烃链上C-3,ω-1及ω-3位置(图3 ). ...
... ,
45 ,
49 ]
HGs structures<sup>[<xref ref-type="bibr" rid="R19">19</xref>,<xref ref-type="bibr" rid="R45">45</xref>,<xref ref-type="bibr" rid="R49">49</xref>]</sup> Fig.3 ![]()
对HGs的缩写命名没有统一规定,目前其缩写先后标明糖苷基头碳原子数、烷烃链碳原子数及烷烃链官能团信息.在C5 HGs被发现之前,C6 HGs的缩写一般仅包括烷烃链碳原子数及烷烃链官能团信息,即Cn n 代表烷烃链碳原子数.如C26 HG diol指代由六碳糖苷与含二十六碳的二醇连接形成的糖脂.随着C5 HGs被解析出来后,HGs的缩写要指明是六碳或五碳糖基头,烷烃链碳原子数则以下标标在HG字母后,即Cm n m 代表糖基头碳原子数,n 代表烷烃链碳原子数.如C5 HG30 triol指代由五碳糖苷与含三十碳的三醇连接形成的糖脂,而上述C26 HG diol则改为C6 HG26 diol.但是上述命名不包括甲基化HGs,并且HGs化合物的缩写命名并不能体现出烷烃链上官能团所在位置,也不能体现出糖苷基头的结构类型. ...
... ,
45 ,
49 ]
Fig.3 ![]()
对HGs的缩写命名没有统一规定,目前其缩写先后标明糖苷基头碳原子数、烷烃链碳原子数及烷烃链官能团信息.在C5 HGs被发现之前,C6 HGs的缩写一般仅包括烷烃链碳原子数及烷烃链官能团信息,即Cn n 代表烷烃链碳原子数.如C26 HG diol指代由六碳糖苷与含二十六碳的二醇连接形成的糖脂.随着C5 HGs被解析出来后,HGs的缩写要指明是六碳或五碳糖基头,烷烃链碳原子数则以下标标在HG字母后,即Cm n m 代表糖基头碳原子数,n 代表烷烃链碳原子数.如C5 HG30 triol指代由五碳糖苷与含三十碳的三醇连接形成的糖脂,而上述C26 HG diol则改为C6 HG26 diol.但是上述命名不包括甲基化HGs,并且HGs化合物的缩写命名并不能体现出烷烃链上官能团所在位置,也不能体现出糖苷基头的结构类型. ...
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
Rapid analysis of long-chain glycolipids in heterocystous cyanobacteria using high-performance liquid chromatography coupled to electrospray ionization tandem mass spectrometry
1
2009
... 尽管HGs发现于20世纪70年代左右,但相关研究在很长时间里仅限于对培养菌株的膜脂结构与成分分析.直到2009年Bauersachs等[46 ] 开发出HGs HPLC分析方法,并成功在荷兰北海堰洲岛Schiermonnikoog中检测出HGs后,才极大地促进了对于HGs在自然环境样品中的研究.2017年Bale等[54 ] 提出HGs的绝对定量方法,又进一步推动了其在自然环境,特别是古环境中的研究与应用. ...
Chemistry of glycolipids from the heterocysts of nitrogen-fixing cyanobacteria
1
1999
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
Heterocyst glycolipids from five nitrogen-fixing cyanobacteria
1
1996
... HGs由糖苷基连接长链二醇、三醇、酮醇或酮二醇烷基组成.糖基头通常为六碳糖苷(C6 ),最近又发现了五碳糖苷(C5 ).C6 糖大多为葡萄糖,同时也存在少数半乳糖[48 ] 或甘露糖[49 ] .最新的研究发现Calothrix 异形胞糖脂C6 糖基头C-6位置连有一甲基[45 ] .烷基烃链一般含有26~32个碳原子,羟基或羰基位于烃链上C-3,ω-1及ω-3位置(图3 ). ...
Heterocyst glycolipids from nitrogen-fixing cyanobacteria other than nostocaceae
3
1998
... HGs由糖苷基连接长链二醇、三醇、酮醇或酮二醇烷基组成.糖基头通常为六碳糖苷(C6 ),最近又发现了五碳糖苷(C5 ).C6 糖大多为葡萄糖,同时也存在少数半乳糖[48 ] 或甘露糖[49 ] .最新的研究发现Calothrix 异形胞糖脂C6 糖基头C-6位置连有一甲基[45 ] .烷基烃链一般含有26~32个碳原子,羟基或羰基位于烃链上C-3,ω-1及ω-3位置(图3 ). ...
... ,
49 ]
HGs structures<sup>[<xref ref-type="bibr" rid="R19">19</xref>,<xref ref-type="bibr" rid="R45">45</xref>,<xref ref-type="bibr" rid="R49">49</xref>]</sup> Fig.3 ![]()
对HGs的缩写命名没有统一规定,目前其缩写先后标明糖苷基头碳原子数、烷烃链碳原子数及烷烃链官能团信息.在C5 HGs被发现之前,C6 HGs的缩写一般仅包括烷烃链碳原子数及烷烃链官能团信息,即Cn n 代表烷烃链碳原子数.如C26 HG diol指代由六碳糖苷与含二十六碳的二醇连接形成的糖脂.随着C5 HGs被解析出来后,HGs的缩写要指明是六碳或五碳糖基头,烷烃链碳原子数则以下标标在HG字母后,即Cm n m 代表糖基头碳原子数,n 代表烷烃链碳原子数.如C5 HG30 triol指代由五碳糖苷与含三十碳的三醇连接形成的糖脂,而上述C26 HG diol则改为C6 HG26 diol.但是上述命名不包括甲基化HGs,并且HGs化合物的缩写命名并不能体现出烷烃链上官能团所在位置,也不能体现出糖苷基头的结构类型. ...
... ,
49 ]
Fig.3 ![]()
对HGs的缩写命名没有统一规定,目前其缩写先后标明糖苷基头碳原子数、烷烃链碳原子数及烷烃链官能团信息.在C5 HGs被发现之前,C6 HGs的缩写一般仅包括烷烃链碳原子数及烷烃链官能团信息,即Cn n 代表烷烃链碳原子数.如C26 HG diol指代由六碳糖苷与含二十六碳的二醇连接形成的糖脂.随着C5 HGs被解析出来后,HGs的缩写要指明是六碳或五碳糖基头,烷烃链碳原子数则以下标标在HG字母后,即Cm n m 代表糖基头碳原子数,n 代表烷烃链碳原子数.如C5 HG30 triol指代由五碳糖苷与含三十碳的三醇连接形成的糖脂,而上述C26 HG diol则改为C6 HG26 diol.但是上述命名不包括甲基化HGs,并且HGs化合物的缩写命名并不能体现出烷烃链上官能团所在位置,也不能体现出糖苷基头的结构类型. ...
Intact polar glycosidic GDGTs in sediments settle from water column as evidenced from downcore sediment records
1
2018
... 另外,活体微生物膜脂化合物是完整极性脂(Intact Polar Lipid,IPL),其中含具有环境示踪意义的核心脂(Core Lipid,CL).一般来说IPL存在于活细胞膜内,在细胞死亡降解后极性头(如磷脂)极易降解生成CL进入环境载体保存下来.但糖脂相对于磷脂较难降解,能够在沉积物中长久保存[50 ] .目前看来,从异形胞蓝细菌纯培养所提取出的HGs结构与自然环境包括沉积物中发现的HGs一致,所以HGs也能够在活体死亡后较长久地保存于沉积物中. ...
Distribution of long chain heterocyst glycolipids in N2 -fixing cyanobacteria of the order Stigonematales
1
2014
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
Distribution of heterocyst glycolipids in cyanobacteria
2
70
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
... 微生物细胞膜内脂类分布会随着生存环境的变化而变化,尤其对温度比较敏感[55 ] .这一特征被广泛应用于古环境的温度重建.2009年Bauersachs等[52 ] 在培养3株异形胞蓝细菌(Anabaena CCY9613, Nostoc CCY0012及Nostoc CCY9926)时发现,其酮—醇结构相较于二醇结构HGs的比例会随着温度的升高而降低,并提出HG26 指标,其公式定义为C6 HG26 keto-ol占总C6 HG26 (包括C6 HG26 diol和C6 HG26 keto-ol)的比例. ...
Identification of the glycosyl transferase required for synthesis of the principal glycolipid characteristic of heterocysts of Anabaena sp. strain PCC 7120
1
2007
... 目前,所有分化出异形细胞的蓝细菌都含有HGs,并且HGs仅发现于异形胞蓝细菌膜脂中.也就是说,HGs是异形胞蓝细菌的独特生物标志化合物.培养研究表明,通过HGs结构能在科甚至属的层面上区分不同类型的蓝细菌[6 ,19 ,47 ,51 ,52 ] .如C6 HG26 diol及其相应酮—醇异形胞糖脂主要产出于Nostocaceae 科异形胞蓝细菌;C6 HG30 triol及相应酮—二醇异形胞糖脂主要产出于Scytonemataceae 科蓝细菌中;而C6 HG32 triol及相应酮—二醇异形胞糖脂则主要产出于Stigonematales目蓝细菌菌株.上述C6 糖脂目前皆发现于陆地咸淡水环境中独立生活的异形胞蓝细菌中,而在海洋共生种中,其糖脂则为C5 糖脂,主要为C5 HG30 diol与C5 HG30 triol.共生种与独立种的上述差异或许是由于共生异形胞蓝细菌适应宿主体内高浓度氧气的结果,由hglT 基因调控[53 ] .基于菌株培养进行的有关研究得出的资料还不是很完整,有关内容还会不断完善和补充.例如,最近发现,比较罕见的海洋独立种Calothrix CCY1611产出的是特殊结构的糖脂,即甲基化C6 HG28 triol[45 ] . ...
A method for quantifying heterocyst glycolipids in biomass and sediments
1
2017
... 尽管HGs发现于20世纪70年代左右,但相关研究在很长时间里仅限于对培养菌株的膜脂结构与成分分析.直到2009年Bauersachs等[46 ] 开发出HGs HPLC分析方法,并成功在荷兰北海堰洲岛Schiermonnikoog中检测出HGs后,才极大地促进了对于HGs在自然环境样品中的研究.2017年Bale等[54 ] 提出HGs的绝对定量方法,又进一步推动了其在自然环境,特别是古环境中的研究与应用. ...
Microbial fatty acids and thermal adaptation
1
1994
... 微生物细胞膜内脂类分布会随着生存环境的变化而变化,尤其对温度比较敏感[55 ] .这一特征被广泛应用于古环境的温度重建.2009年Bauersachs等[52 ] 在培养3株异形胞蓝细菌(Anabaena CCY9613, Nostoc CCY0012及Nostoc CCY9926)时发现,其酮—醇结构相较于二醇结构HGs的比例会随着温度的升高而降低,并提出HG26 指标,其公式定义为C6 HG26 keto-ol占总C6 HG26 (包括C6 HG26 diol和C6 HG26 keto-ol)的比例. ...
Temperature induced changes in the heterocyst glycolipid composition of N2 fixing heterocystous cyanobacteria
1
2014
... 2014年Bauersachs等[56 ] 对HG26 (其文中写作HGI26 )指数与温度关系作了进一步探讨,并发现酮—醇结构HGs同分异构的比例会随着温度变化而变化.他们在文章中另外提出了基于HG28 diol与其相应的酮—醇HGs的指标(HGI28 ),以及基于同分异构HGs的指标等,并建立了温度与HGI26 和HGI28 等的换算关系式,但不同菌株呈现出不同的T-HGI26或28 关系.之后,Bauersachs等[57 ] 在德国淡水湖Lake Schreventeich采集水颗粒物及表层沉积物对HG26 指示温度作进一步分析验证,重新定义HDI(Heterocyst Diol Index)为diol-HGs占其相关keto-ol-HGs的比例,使HDI与温度正相关,又提出基于C6 HG28 triol及其相应酮—醇异形胞糖脂指标(HTI28 ).他们通过双尾t检验计算验证得到3个HGs指标与表层水温度的显著相关性(r 2 >0.70,p <0.001),同时也发现HGs指标与水体溶解氧含量或pH没有或者仅有微弱相关性.这是首个在自然环境中验证HGs温度指标的报告. ...
Seasonal lake surface water temperature trends reflected by heterocyst glycolipid-based molecular thermometers
1
2015
... 2014年Bauersachs等[56 ] 对HG26 (其文中写作HGI26 )指数与温度关系作了进一步探讨,并发现酮—醇结构HGs同分异构的比例会随着温度变化而变化.他们在文章中另外提出了基于HG28 diol与其相应的酮—醇HGs的指标(HGI28 ),以及基于同分异构HGs的指标等,并建立了温度与HGI26 和HGI28 等的换算关系式,但不同菌株呈现出不同的T-HGI26或28 关系.之后,Bauersachs等[57 ] 在德国淡水湖Lake Schreventeich采集水颗粒物及表层沉积物对HG26 指示温度作进一步分析验证,重新定义HDI(Heterocyst Diol Index)为diol-HGs占其相关keto-ol-HGs的比例,使HDI与温度正相关,又提出基于C6 HG28 triol及其相应酮—醇异形胞糖脂指标(HTI28 ).他们通过双尾t检验计算验证得到3个HGs指标与表层水温度的显著相关性(r 2 >0.70,p <0.001),同时也发现HGs指标与水体溶解氧含量或pH没有或者仅有微弱相关性.这是首个在自然环境中验证HGs温度指标的报告. ...
The Holocene sedimentary record of cyanobacterial glycolipids in the Baltic Sea: An evaluation of their application as tracers of past nitrogen fixation
5
2017
... Sollai等[58 ] 首次尝试以上述HDI26或28 公式计算波罗的海全新世以来温度变化,发现从波罗的海演化至半咸水时期之后,相较于TEX86 重建记录(<17.5 ℃)显示出不正常的高值(约23 ℃),可能与异形胞固氮蓝细菌种群结构的变化有关.目前HGI26或28 仅在部分现代淡水环境表现出与温度的相关性,因此,这一指标若想应用于全球古温度重建,还需要更多的研究,包括验证HGs分布是否会受盐度和种群结构等环境参数影响等. ...
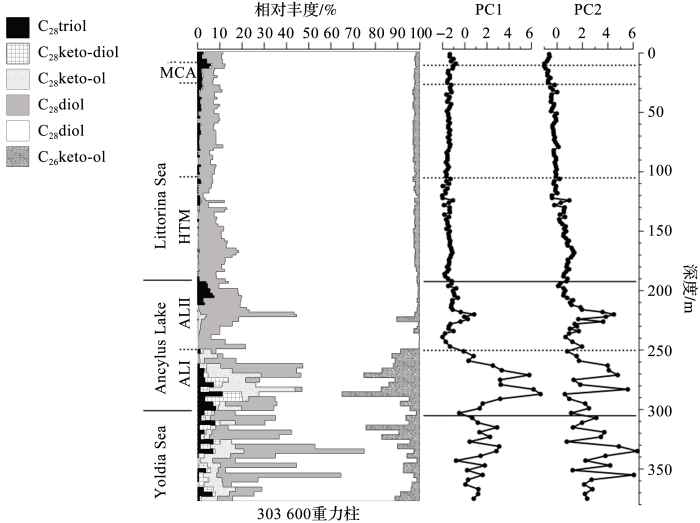
... Bauersachs等[7 ] 提出可以用包括HGs等作为有害蓝细菌的生标化合物,追踪波罗的海地质历史上的蓝细菌勃发情况,并给出了几株异形胞蓝细菌分布的比例情况.同年,Sollai等[58 ] 利用HGs在波罗的海沉积柱的分布情况反演了波罗的海地质史上异形胞蓝细菌群落变化,发现其与波罗的海水文环境变化相对应.该研究涉及到全世新以来的样品.结合HGs分布的主成分分析(Principal Component Analysis,PCA)等数据,分析了波罗的海不同水文环境时期演变,包括早期淡水期、海水侵入期及完全半咸水期(图5 ).在早期淡水时期,(C6 ) HG26 diol与(C6 ) HG28 diol占主导,反映出此时期以例如Anabaena 及Aphanizomenon 为优势异形胞蓝细菌;在距今7.2~3.5 ka,波罗的海由淡水完全转变为半咸水后,(C6 ) HG26 diol逐渐占主导,反映出以Nostocaceae 科蓝细菌为优势菌的分布情况. ...
... [
58 ]
The distribution of HGs in a sediment core in Baltic Sea and related hydrographical phases over the Holocene<sup>[<xref ref-type="bibr" rid="R58">58</xref>]</sup> Fig.5 ![]()
3.3.3 HGs分布与陆源有机质输入 每年有大量的陆地有机质会通过河流或风尘输入海洋中,构成海洋环境中有机碳的一大来源.海陆过渡带则是决定陆源有机碳最终归宿的关键区域:或者被封存到沉积物中成为碳汇,或者被氧化降解返回海洋—大气圈.因此,定量研究陆源有机质在海陆过渡带中总有机质的份额是区域和全球碳循环研究中的重要内容. ...
... [
58 ]
Fig.5 ![]()
3.3.3 HGs分布与陆源有机质输入 每年有大量的陆地有机质会通过河流或风尘输入海洋中,构成海洋环境中有机碳的一大来源.海陆过渡带则是决定陆源有机碳最终归宿的关键区域:或者被封存到沉积物中成为碳汇,或者被氧化降解返回海洋—大气圈.因此,定量研究陆源有机质在海陆过渡带中总有机质的份额是区域和全球碳循环研究中的重要内容. ...
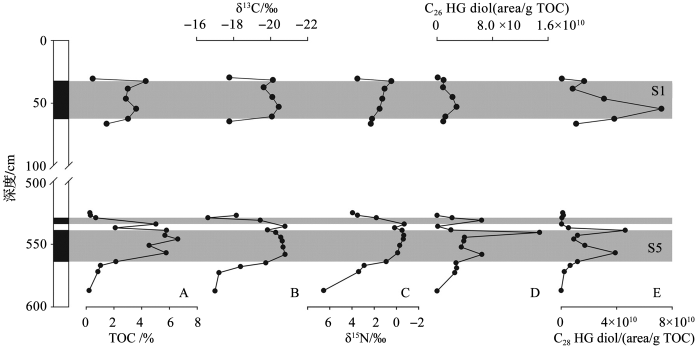
... 2010年,Bauersachs等[59 ] 首次利用HGs耦合δ15 N重建北冰洋始新世固氮活动.在北冰洋始新世沉积物中发现大量HGs的存在,并且当C6 HG26或28 diol含量升高时,δ15 N更偏负(图6 ).作者认为北冰洋始新世存在大量的异形胞蓝细菌,并且此时异形胞蓝细菌与Azolla 共生,当Azolla 孢子数量升高时,C6 HG26或28 diol同时升高.此时δ15 N平均为-1‰,与Azolla 培养条件下的同位素值一致,极好地反映出异形胞蓝细菌在此时固氮活动中发挥了重要作用,进一步证明了HGs指示固氮活动的潜力.之后Sollai等[58 ] 同样也在全新世沉积物中发现了δ15 N与HGs含量良好的对应关系.例如,当δ15 N变化范围为4‰~6‰时(通常认为这一δ15 N范围表明浮游植物的营养来源为NH4 + 3 - 15 N变得更负为1‰~3‰时,HGs含量明显升高. ...
Fossilized glycolipids reveal past oceanic N2 fixation by heterocystous cyanobacteria
6
2010
... 如前所述,C6 HGs由独立生存异形胞蓝细菌产生,通常生活于淡水或半咸水中,C5 HGs由与硅藻共生异形胞蓝细菌产生,分布于海洋中.Bale等[6 ] 在亚马孙陆架表层水体中检测到大量C5 HGs,但未检测到C6 HGs;Bauersachs等[59 ] 也提到未在现代海洋沉积物中检测到HGs(即C6 HGs).基于以上报道,我们最近研究发现,根据C6 HGs与C5 HGs含量比例,可以指示长江口—东海中陆源有机质输入情况,据此提出了一个新指标FHG [60 ] .我们的进一步工作还发现FHG 同样可以较好地应用于指示珠江口—南海区域的陆源有机质输入情况,并且在长江口外4 m长沉积柱中与BIT(Branched and Isoprenoid Tetraether)变化趋势较为一致,可以运用于长江口陆源有机质输入的历史重建中. ...
... 2010年,Bauersachs等[59 ] 首次利用HGs耦合δ15 N重建北冰洋始新世固氮活动.在北冰洋始新世沉积物中发现大量HGs的存在,并且当C6 HG26或28 diol含量升高时,δ15 N更偏负(图6 ).作者认为北冰洋始新世存在大量的异形胞蓝细菌,并且此时异形胞蓝细菌与Azolla 共生,当Azolla 孢子数量升高时,C6 HG26或28 diol同时升高.此时δ15 N平均为-1‰,与Azolla 培养条件下的同位素值一致,极好地反映出异形胞蓝细菌在此时固氮活动中发挥了重要作用,进一步证明了HGs指示固氮活动的潜力.之后Sollai等[58 ] 同样也在全新世沉积物中发现了δ15 N与HGs含量良好的对应关系.例如,当δ15 N变化范围为4‰~6‰时(通常认为这一δ15 N范围表明浮游植物的营养来源为NH4 + 3 - 15 N变得更负为1‰~3‰时,HGs含量明显升高. ...
... [
59 ]
HGs含量增加与δ15 N的负偏同时发生,代表固氮活动的增强 ...
... HGs含量增加与δ
15 N的负偏同时发生,代表固氮活动的增强
The bulk properties and HGs contents of in a sediment core in the Eastern Mediterranean<sup>[<xref ref-type="bibr" rid="R59">59</xref>]</sup> The increased contents of HGs coincide with a decrease of δ15 N suggesting enhanced dinitrogen fixation ...
... HGs具有作为古环境指标所需要稳定性.目前看来,尽管HGs一端连有糖苷极性头,但仍可通过超声法,甚至是ASE方法对其进行提取[59 ] .另外,在微生物纯培养中所提取出的HGs结构与自然环境包括沉积柱中分析的结构一致.C6 HGs已发现于最老49 Ma的沉积物中[59 ] .虽然目前没有C5 HGs在古环境研究中的相关报道,但作者所在实验室在琼东南海域采集的末次冰期以来的柱状沉积物中检测到了含量丰富的C5 HGs,说明其结构是比较稳定的,有可能用于追踪第四纪甚至新生代的环境变化. ...
... [59 ].虽然目前没有C5 HGs在古环境研究中的相关报道,但作者所在实验室在琼东南海域采集的末次冰期以来的柱状沉积物中检测到了含量丰富的C5 HGs,说明其结构是比较稳定的,有可能用于追踪第四纪甚至新生代的环境变化. ...
Assessment of sedimentary heterocyst glycolipids as tracers of freshwater input to the Changjiang Estuary and East China Sea
1
2019
... 如前所述,C6 HGs由独立生存异形胞蓝细菌产生,通常生活于淡水或半咸水中,C5 HGs由与硅藻共生异形胞蓝细菌产生,分布于海洋中.Bale等[6 ] 在亚马孙陆架表层水体中检测到大量C5 HGs,但未检测到C6 HGs;Bauersachs等[59 ] 也提到未在现代海洋沉积物中检测到HGs(即C6 HGs).基于以上报道,我们最近研究发现,根据C6 HGs与C5 HGs含量比例,可以指示长江口—东海中陆源有机质输入情况,据此提出了一个新指标FHG [60 ] .我们的进一步工作还发现FHG 同样可以较好地应用于指示珠江口—南海区域的陆源有机质输入情况,并且在长江口外4 m长沉积柱中与BIT(Branched and Isoprenoid Tetraether)变化趋势较为一致,可以运用于长江口陆源有机质输入的历史重建中. ...
The marine nitrogen cycle: Overview and challenges
1
2008
... 海洋中94%氮元素是以氮气形式存在[61 ] ,而氮气不能被大多数微生物直接利用,因此微生物固氮对于以氮限制为主要特征的海洋生态系统十分重要.生物固氮活动是海洋氮循环的起点,是海洋中最重要的固定氮来源,对海洋真核生物的发展演化起着重要的作用,同时也间接影响着碳循环.而蓝细菌作为海洋中最重要的固氮微生物,追踪其地质历史上的定量变化对还原古环境固氮过程十分重要. ...
2-Methylhopanoids as biomarkers for cyanobacterial oxygenic photosynthesis
1
1999
... 用生物标志物示踪微生物固氮历史已有多年历史,比如,2-甲基—细菌藿多醇或其成岩产物2-甲基—藿烷被认为是蓝细菌的生标物质,被广泛用于重建海洋固氮活动[62 ,63 ] .但有研究发现,编码2-甲基—藿多醇合成酶的基因也被发现存在于细菌域其他的部分微生物,如α-变形菌和醋酸杆菌,并非蓝细菌特有[64 ] .一些非蓝细菌菌株,如Rhodopseudomonas palustris TIE-1[65 ] 等被发现可以合成2-甲基—藿多醇.因此,需要谨慎运用2-甲基—细菌藿多醇来重建蓝细菌的活动历史. ...
N2 -fixing cyanobacteria supplied nutrient N for Cretaceous oceanic anoxic events
1
2004
... 用生物标志物示踪微生物固氮历史已有多年历史,比如,2-甲基—细菌藿多醇或其成岩产物2-甲基—藿烷被认为是蓝细菌的生标物质,被广泛用于重建海洋固氮活动[62 ,63 ] .但有研究发现,编码2-甲基—藿多醇合成酶的基因也被发现存在于细菌域其他的部分微生物,如α-变形菌和醋酸杆菌,并非蓝细菌特有[64 ] .一些非蓝细菌菌株,如Rhodopseudomonas palustris TIE-1[65 ] 等被发现可以合成2-甲基—藿多醇.因此,需要谨慎运用2-甲基—细菌藿多醇来重建蓝细菌的活动历史. ...
Identification of a methylase required for 2-methylhopanoid production and implications for the interpretation of sedimentary hopanes
1
2010
... 用生物标志物示踪微生物固氮历史已有多年历史,比如,2-甲基—细菌藿多醇或其成岩产物2-甲基—藿烷被认为是蓝细菌的生标物质,被广泛用于重建海洋固氮活动[62 ,63 ] .但有研究发现,编码2-甲基—藿多醇合成酶的基因也被发现存在于细菌域其他的部分微生物,如α-变形菌和醋酸杆菌,并非蓝细菌特有[64 ] .一些非蓝细菌菌株,如Rhodopseudomonas palustris TIE-1[65 ] 等被发现可以合成2-甲基—藿多醇.因此,需要谨慎运用2-甲基—细菌藿多醇来重建蓝细菌的活动历史. ...
Biosynthesis of 2-methylbacteriohopanepolyols by an anoxygenic phototroph
1
2007
... 用生物标志物示踪微生物固氮历史已有多年历史,比如,2-甲基—细菌藿多醇或其成岩产物2-甲基—藿烷被认为是蓝细菌的生标物质,被广泛用于重建海洋固氮活动[62 ,63 ] .但有研究发现,编码2-甲基—藿多醇合成酶的基因也被发现存在于细菌域其他的部分微生物,如α-变形菌和醋酸杆菌,并非蓝细菌特有[64 ] .一些非蓝细菌菌株,如Rhodopseudomonas palustris TIE-1[65 ] 等被发现可以合成2-甲基—藿多醇.因此,需要谨慎运用2-甲基—细菌藿多醇来重建蓝细菌的活动历史. ...
Isotopic fractionation of nitrogen and carbon in the synthesis of amino acids by microorganisms
1
1987
... 而HGs目前仅发现于分化异形细胞的蓝细菌,可以较肯定地认为HGs的检出指示了固氮活动的发生(具体见本文第2章).另外,在培养实验中发现,蓝细菌固氮过程中的氮同位素分馏(相较大气δ15 N~0)约为-2.6‰,固氮产物的δ15 N变化范围为-3‰~0[66 ] .近年来多个研究将HGs与δ15 N结合分析,验证了HGs与固氮活动之间的直接联系. ...
High diatom production and export in stratified waters—A potential negative feedback to global warming
1
2013
... HGs沉积记录能够为我们提供丰富而新颖的信息.首先,目前仅在异形胞蓝细菌膜脂中检测到HGs的存在,相较于其他常用的古环境生标来说,HGs最明显的优势在于有确定的生物来源以及在不同种属间的某些分布差异.例如C6 HG32 triol与其相应酮—二醇异形胞糖脂目前只发现于Stigonematales目蓝细菌菌株中.而此类蓝细菌通常生活于热带及亚热带淡水或地表环境中,因此C6 HG32 triol及keto-ol在湖相沉积物中的出现或许可以反映陆地古湖泊和古气候的某些信息.其次,结合异形胞蓝细菌固氮功能,HGs确定的生源降低了追踪古环境固氮活动的难度.一直以来,对于固氮活动的追踪是通过沉积物有机质氮同位素,但是同位素信息会受多种因素的干扰,具有多解性,不是固氮作用的确切指标.因此,C6 HGs与C5 HGs含量记录,结合δ15 N信息,可以分别用来重建湖泊及海洋历史固氮情况.第三,对于海洋沉积物中的C5 HGs,除能用于指示固氮信息外,还能反映固氮蓝细菌对海洋碳汇的贡献.这是因为生产C5 HGs的固氮蓝细菌通常与硅藻共生,而硅藻较重的硅质细胞在水体中会携带C5 HGs沉入海底,把蓝细菌固定的碳也能快速输入到深海,起到有效的生物碳泵作用.上层海水分层情况下往往存在大量硅藻—异形胞蓝细菌共生体,异形胞蓝细菌固氮提供营养导致硅藻的勃发.例如在北太平洋副热带环流分层水体中就存在这种勃发,虽然持续时间只有几天,但其能够快速沉降,构成当地总POC年通量的20%,对全球变暧起到重要的负反馈作用[67 ] .在过去的报道中,C5 HGs在3 000 m水深处被检测到[6 ,68 ] ,说明其可以有效地被输出至深海和沉积物中.据此,可以实现对海洋异形胞蓝细菌活动及其固碳与储碳过程的追踪,重建有关的碳氮生物地球化学过程. ...
C5 glycolipids of heterocystous cyanobacteria track symbiont abundance in the diatom Hemiaulus hauckii across the tropical North Atlantic
1
2018
... HGs沉积记录能够为我们提供丰富而新颖的信息.首先,目前仅在异形胞蓝细菌膜脂中检测到HGs的存在,相较于其他常用的古环境生标来说,HGs最明显的优势在于有确定的生物来源以及在不同种属间的某些分布差异.例如C6 HG32 triol与其相应酮—二醇异形胞糖脂目前只发现于Stigonematales目蓝细菌菌株中.而此类蓝细菌通常生活于热带及亚热带淡水或地表环境中,因此C6 HG32 triol及keto-ol在湖相沉积物中的出现或许可以反映陆地古湖泊和古气候的某些信息.其次,结合异形胞蓝细菌固氮功能,HGs确定的生源降低了追踪古环境固氮活动的难度.一直以来,对于固氮活动的追踪是通过沉积物有机质氮同位素,但是同位素信息会受多种因素的干扰,具有多解性,不是固氮作用的确切指标.因此,C6 HGs与C5 HGs含量记录,结合δ15 N信息,可以分别用来重建湖泊及海洋历史固氮情况.第三,对于海洋沉积物中的C5 HGs,除能用于指示固氮信息外,还能反映固氮蓝细菌对海洋碳汇的贡献.这是因为生产C5 HGs的固氮蓝细菌通常与硅藻共生,而硅藻较重的硅质细胞在水体中会携带C5 HGs沉入海底,把蓝细菌固定的碳也能快速输入到深海,起到有效的生物碳泵作用.上层海水分层情况下往往存在大量硅藻—异形胞蓝细菌共生体,异形胞蓝细菌固氮提供营养导致硅藻的勃发.例如在北太平洋副热带环流分层水体中就存在这种勃发,虽然持续时间只有几天,但其能够快速沉降,构成当地总POC年通量的20%,对全球变暧起到重要的负反馈作用[67 ] .在过去的报道中,C5 HGs在3 000 m水深处被检测到[6 ,68 ] ,说明其可以有效地被输出至深海和沉积物中.据此,可以实现对海洋异形胞蓝细菌活动及其固碳与储碳过程的追踪,重建有关的碳氮生物地球化学过程. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}