The eastern extent of seasonal iron limitation in the high latitude North Atlantic Ocean
2
2019
... 铁在海洋环境中的分布具有空间异质性,是影响海洋初级生产力的限制因子之一[1 ] .由于铁在海洋中主要以溶解度较低的三价态(Fe3+ )形式存在,这使得铁容易发生沉降而被“清除”,因此溶解铁对海洋生物而言是一种稀缺资源.大洋环境铁的浓度只有0.1~1.0 nmol/L[1 ] ,赤道和一般海区铁含量也仅维持在0.3~0.5 nmol/L,这些低浓度的可利用铁成为了限制海洋浮游植物的因素(一般情况下,浮游植物依赖铁的下线阈值为0.3 nmol/L)[2 ] .高营养—低叶绿素(High Nutrient-Low Chlorophyll,HNLC)海区的发现以及冰芯样品中铁和CO2 浓度的负相关性,催生了铁假说理论,该假说认为铁缺乏限制了藻类对营养盐的吸收,导致浮游生物叶绿素含量和生产力水平下降[3 ] .在随后的研究中,人们逐渐认识到了铁限制浮游植物的相关机制,包括作为辅酶因子,参与光合作用、呼吸作用以及固氮过程等[4 ] .进入21世纪,人们普遍认同了铁在海洋生产力和生物地球化学方面的重要性,并将其纳入全球海洋模式中. ...
... [1 ],赤道和一般海区铁含量也仅维持在0.3~0.5 nmol/L,这些低浓度的可利用铁成为了限制海洋浮游植物的因素(一般情况下,浮游植物依赖铁的下线阈值为0.3 nmol/L)[2 ] .高营养—低叶绿素(High Nutrient-Low Chlorophyll,HNLC)海区的发现以及冰芯样品中铁和CO2 浓度的负相关性,催生了铁假说理论,该假说认为铁缺乏限制了藻类对营养盐的吸收,导致浮游生物叶绿素含量和生产力水平下降[3 ] .在随后的研究中,人们逐渐认识到了铁限制浮游植物的相关机制,包括作为辅酶因子,参与光合作用、呼吸作用以及固氮过程等[4 ] .进入21世纪,人们普遍认同了铁在海洋生产力和生物地球化学方面的重要性,并将其纳入全球海洋模式中. ...
The effect of iron to the primary production in the ocean
1
2007
... 铁在海洋环境中的分布具有空间异质性,是影响海洋初级生产力的限制因子之一[1 ] .由于铁在海洋中主要以溶解度较低的三价态(Fe3+ )形式存在,这使得铁容易发生沉降而被“清除”,因此溶解铁对海洋生物而言是一种稀缺资源.大洋环境铁的浓度只有0.1~1.0 nmol/L[1 ] ,赤道和一般海区铁含量也仅维持在0.3~0.5 nmol/L,这些低浓度的可利用铁成为了限制海洋浮游植物的因素(一般情况下,浮游植物依赖铁的下线阈值为0.3 nmol/L)[2 ] .高营养—低叶绿素(High Nutrient-Low Chlorophyll,HNLC)海区的发现以及冰芯样品中铁和CO2 浓度的负相关性,催生了铁假说理论,该假说认为铁缺乏限制了藻类对营养盐的吸收,导致浮游生物叶绿素含量和生产力水平下降[3 ] .在随后的研究中,人们逐渐认识到了铁限制浮游植物的相关机制,包括作为辅酶因子,参与光合作用、呼吸作用以及固氮过程等[4 ] .进入21世纪,人们普遍认同了铁在海洋生产力和生物地球化学方面的重要性,并将其纳入全球海洋模式中. ...
铁对海洋初级生产力的影响
1
2007
... 铁在海洋环境中的分布具有空间异质性,是影响海洋初级生产力的限制因子之一[1 ] .由于铁在海洋中主要以溶解度较低的三价态(Fe3+ )形式存在,这使得铁容易发生沉降而被“清除”,因此溶解铁对海洋生物而言是一种稀缺资源.大洋环境铁的浓度只有0.1~1.0 nmol/L[1 ] ,赤道和一般海区铁含量也仅维持在0.3~0.5 nmol/L,这些低浓度的可利用铁成为了限制海洋浮游植物的因素(一般情况下,浮游植物依赖铁的下线阈值为0.3 nmol/L)[2 ] .高营养—低叶绿素(High Nutrient-Low Chlorophyll,HNLC)海区的发现以及冰芯样品中铁和CO2 浓度的负相关性,催生了铁假说理论,该假说认为铁缺乏限制了藻类对营养盐的吸收,导致浮游生物叶绿素含量和生产力水平下降[3 ] .在随后的研究中,人们逐渐认识到了铁限制浮游植物的相关机制,包括作为辅酶因子,参与光合作用、呼吸作用以及固氮过程等[4 ] .进入21世纪,人们普遍认同了铁在海洋生产力和生物地球化学方面的重要性,并将其纳入全球海洋模式中. ...
The case for iron
1
1991
... 铁在海洋环境中的分布具有空间异质性,是影响海洋初级生产力的限制因子之一[1 ] .由于铁在海洋中主要以溶解度较低的三价态(Fe3+ )形式存在,这使得铁容易发生沉降而被“清除”,因此溶解铁对海洋生物而言是一种稀缺资源.大洋环境铁的浓度只有0.1~1.0 nmol/L[1 ] ,赤道和一般海区铁含量也仅维持在0.3~0.5 nmol/L,这些低浓度的可利用铁成为了限制海洋浮游植物的因素(一般情况下,浮游植物依赖铁的下线阈值为0.3 nmol/L)[2 ] .高营养—低叶绿素(High Nutrient-Low Chlorophyll,HNLC)海区的发现以及冰芯样品中铁和CO2 浓度的负相关性,催生了铁假说理论,该假说认为铁缺乏限制了藻类对营养盐的吸收,导致浮游生物叶绿素含量和生产力水平下降[3 ] .在随后的研究中,人们逐渐认识到了铁限制浮游植物的相关机制,包括作为辅酶因子,参与光合作用、呼吸作用以及固氮过程等[4 ] .进入21世纪,人们普遍认同了铁在海洋生产力和生物地球化学方面的重要性,并将其纳入全球海洋模式中. ...
Heme in the marine environment: From cells to the iron cycle
2
2014
... 铁在海洋环境中的分布具有空间异质性,是影响海洋初级生产力的限制因子之一[1 ] .由于铁在海洋中主要以溶解度较低的三价态(Fe3+ )形式存在,这使得铁容易发生沉降而被“清除”,因此溶解铁对海洋生物而言是一种稀缺资源.大洋环境铁的浓度只有0.1~1.0 nmol/L[1 ] ,赤道和一般海区铁含量也仅维持在0.3~0.5 nmol/L,这些低浓度的可利用铁成为了限制海洋浮游植物的因素(一般情况下,浮游植物依赖铁的下线阈值为0.3 nmol/L)[2 ] .高营养—低叶绿素(High Nutrient-Low Chlorophyll,HNLC)海区的发现以及冰芯样品中铁和CO2 浓度的负相关性,催生了铁假说理论,该假说认为铁缺乏限制了藻类对营养盐的吸收,导致浮游生物叶绿素含量和生产力水平下降[3 ] .在随后的研究中,人们逐渐认识到了铁限制浮游植物的相关机制,包括作为辅酶因子,参与光合作用、呼吸作用以及固氮过程等[4 ] .进入21世纪,人们普遍认同了铁在海洋生产力和生物地球化学方面的重要性,并将其纳入全球海洋模式中. ...
... 对铁赋存状态的理解,除了关注胶体铁与配体之间的络合程度或者颗粒铁的可溶性与可矿性之外,更需关注生物作用对铁的影响.由于铁的稀缺性和理化属性,铁的加工与转化和生物类群紧密相关(图2 )[8 ] .例如,浮游植物和细菌是小型微粒铁池的主要贡献者[20 ] ,而病毒则是构成胶体铁的重要组成部分[21 ] .另外,与金属酶结合的胞内铁在微生物溶解和病毒裂解之后会释放到外周环境中[22 ] .这些释放的胞内铁有助于环境胶体铁池内金属酶、铁结合组分(血红素和铁硫簇)以及储铁蛋白活力的发挥[4 ,22 ] .虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
Biogenic iron dust: A novel approach to ocean iron fertilization as a means of large scale removal of carbon dioxide from the atmosphere
1
2019
... “GEOTRACES”全球计划(www.geotraces.org)是研究铁的全球模式最为系统的计划之一,它采用物理和化学方法记录了整个海洋中铁元素的分布情况.另外,该计划也发起了一系列相互交错的“区段”研究,用于了解大洋“施铁肥”的响应和生态效应.铁肥试验拉开了海洋铁生物地球化学研究的序幕.迄今为止,已在不同海区进行了十几次施铁肥实验,几乎所有的实验都诱导了以硅藻为主的浮游植物大量繁殖,并消耗过剩的营养盐,使叶绿素的含量升高且降低了表层海水中的CO2 分压[5 ] .然而,铁假说的另一半内容,即通过添加铁降低大气中CO2 含量进而缓解温室效应却尚存疑惑,因为藻类的生长有可能产生其他温室气体如二甲基硫化物(Dimethyl Sulfide,DMS).Watson等[6 ] 借助模型计算认为:添加铁降低大气中CO2 的含量应该是一种长效行为,短期添加效果抑或难以实现.近期,Zhou等[7 ] 从化学角度提出了一种修正的假说,即“铁铝假说”,该假说认为铝和铁在冰川间期CO2 浓度和气候变化中扮演着重要的角色,铝与超氧化物结合形成铝—超氧复合物,该复合物可以催化Fe(Ⅲ)的还原,从而促进藻类和细菌对铁的利用,同时铝也可以促进浮游植物对溶解性有机磷的摄取以及对氮和碳的固定.“铁铝假说”对铁施肥结果进行了部分修正,然而“铁铝假说”在刺激浮游植物生长和影响海气CO2 方面的作用还有待进一步证明. ...
Effect of iron supply on Southern Ocean CO2 uptake and implications for glacial atmospheric CO2
1
2000
... “GEOTRACES”全球计划(www.geotraces.org)是研究铁的全球模式最为系统的计划之一,它采用物理和化学方法记录了整个海洋中铁元素的分布情况.另外,该计划也发起了一系列相互交错的“区段”研究,用于了解大洋“施铁肥”的响应和生态效应.铁肥试验拉开了海洋铁生物地球化学研究的序幕.迄今为止,已在不同海区进行了十几次施铁肥实验,几乎所有的实验都诱导了以硅藻为主的浮游植物大量繁殖,并消耗过剩的营养盐,使叶绿素的含量升高且降低了表层海水中的CO2 分压[5 ] .然而,铁假说的另一半内容,即通过添加铁降低大气中CO2 含量进而缓解温室效应却尚存疑惑,因为藻类的生长有可能产生其他温室气体如二甲基硫化物(Dimethyl Sulfide,DMS).Watson等[6 ] 借助模型计算认为:添加铁降低大气中CO2 的含量应该是一种长效行为,短期添加效果抑或难以实现.近期,Zhou等[7 ] 从化学角度提出了一种修正的假说,即“铁铝假说”,该假说认为铝和铁在冰川间期CO2 浓度和气候变化中扮演着重要的角色,铝与超氧化物结合形成铝—超氧复合物,该复合物可以催化Fe(Ⅲ)的还原,从而促进藻类和细菌对铁的利用,同时铝也可以促进浮游植物对溶解性有机磷的摄取以及对氮和碳的固定.“铁铝假说”对铁施肥结果进行了部分修正,然而“铁铝假说”在刺激浮游植物生长和影响海气CO2 方面的作用还有待进一步证明. ...
Aluminum effects on marine phytoplankton: Implications for a revised iron hypothesis (Iron-Aluminum Hypothesis)
1
2018
... “GEOTRACES”全球计划(www.geotraces.org)是研究铁的全球模式最为系统的计划之一,它采用物理和化学方法记录了整个海洋中铁元素的分布情况.另外,该计划也发起了一系列相互交错的“区段”研究,用于了解大洋“施铁肥”的响应和生态效应.铁肥试验拉开了海洋铁生物地球化学研究的序幕.迄今为止,已在不同海区进行了十几次施铁肥实验,几乎所有的实验都诱导了以硅藻为主的浮游植物大量繁殖,并消耗过剩的营养盐,使叶绿素的含量升高且降低了表层海水中的CO2 分压[5 ] .然而,铁假说的另一半内容,即通过添加铁降低大气中CO2 含量进而缓解温室效应却尚存疑惑,因为藻类的生长有可能产生其他温室气体如二甲基硫化物(Dimethyl Sulfide,DMS).Watson等[6 ] 借助模型计算认为:添加铁降低大气中CO2 的含量应该是一种长效行为,短期添加效果抑或难以实现.近期,Zhou等[7 ] 从化学角度提出了一种修正的假说,即“铁铝假说”,该假说认为铝和铁在冰川间期CO2 浓度和气候变化中扮演着重要的角色,铝与超氧化物结合形成铝—超氧复合物,该复合物可以催化Fe(Ⅲ)的还原,从而促进藻类和细菌对铁的利用,同时铝也可以促进浮游植物对溶解性有机磷的摄取以及对氮和碳的固定.“铁铝假说”对铁施肥结果进行了部分修正,然而“铁铝假说”在刺激浮游植物生长和影响海气CO2 方面的作用还有待进一步证明. ...
The integral role of iron in ocean biogeochemistry
12
2017
... 除了物理学的观测和化学的测定,生物作用介导的铁行为也是重要的一环,尤其是微生物驱动下的铁循环过程.现有的研究已表明,微生物在铁的赋存状态、加工、转化、生态链传输以及循环通量上发挥着重要的作用[8 ] .整合全球海洋铁循环中所涉及的微生物过程及其与碳、氮循环的复杂联系成为了当下关注的重点. ...
... 对铁赋存状态的理解,除了关注胶体铁与配体之间的络合程度或者颗粒铁的可溶性与可矿性之外,更需关注生物作用对铁的影响.由于铁的稀缺性和理化属性,铁的加工与转化和生物类群紧密相关(图2 )[8 ] .例如,浮游植物和细菌是小型微粒铁池的主要贡献者[20 ] ,而病毒则是构成胶体铁的重要组成部分[21 ] .另外,与金属酶结合的胞内铁在微生物溶解和病毒裂解之后会释放到外周环境中[22 ] .这些释放的胞内铁有助于环境胶体铁池内金属酶、铁结合组分(血红素和铁硫簇)以及储铁蛋白活力的发挥[4 ,22 ] .虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
... 颗粒和溶解(可溶性和胶体)铁池的成分(据参考文献[
8 ]修改)
<strong>The components of particulate and dissolved iron pools </strong>(<strong>soluble and colloidal components</strong>) (<strong>modified after reference</strong>[<xref ref-type="bibr" rid="R8">8</xref>]) Fig.2 ![]()
对于铁的来源,高纬度海区的铁主要来自河流输入、沉积物的再悬浮以及人类活动的陆源输送;相比而言,低纬度地区的远海或大洋,大气沉降是铁的主要供应方式.除了大气尘埃,上升流和洋中脊热液活动也是远海铁库的重要来源.此外,有机铁络合配体的循环同样也构成了大洋铁的重要组成部分.建模试验表明[8 ] ,配体循环所携带的铁含量大于空气尘埃,并且可实现远距离输送.然而,对于不同环境下配体生成的速率和作用还存在不确定性,对铁络合配体的产生和消耗的理解仍在继续深化中. ...
... [
8 ])
Fig.2 ![]()
对于铁的来源,高纬度海区的铁主要来自河流输入、沉积物的再悬浮以及人类活动的陆源输送;相比而言,低纬度地区的远海或大洋,大气沉降是铁的主要供应方式.除了大气尘埃,上升流和洋中脊热液活动也是远海铁库的重要来源.此外,有机铁络合配体的循环同样也构成了大洋铁的重要组成部分.建模试验表明[8 ] ,配体循环所携带的铁含量大于空气尘埃,并且可实现远距离输送.然而,对于不同环境下配体生成的速率和作用还存在不确定性,对铁络合配体的产生和消耗的理解仍在继续深化中. ...
... 对于铁的来源,高纬度海区的铁主要来自河流输入、沉积物的再悬浮以及人类活动的陆源输送;相比而言,低纬度地区的远海或大洋,大气沉降是铁的主要供应方式.除了大气尘埃,上升流和洋中脊热液活动也是远海铁库的重要来源.此外,有机铁络合配体的循环同样也构成了大洋铁的重要组成部分.建模试验表明[8 ] ,配体循环所携带的铁含量大于空气尘埃,并且可实现远距离输送.然而,对于不同环境下配体生成的速率和作用还存在不确定性,对铁络合配体的产生和消耗的理解仍在继续深化中. ...
... 除了“化学态”铁源,“生物态铁”亦是铁的另一大源泉,这种形式的铁源有3个特征.首先,相对碳、氮、磷而言,铁的化学计量值变化幅度更大.例如,南大洋低铁水域浮游植物中的Fe/P化学计量比通常不足高铁水域的1/5[9 ] ,这种化学计量的可塑性超出了主要营养素的常见水平,但对铁的生物循环却至关重要.其次,浮游动物、细菌和病毒作用下的铁再生对铁的生物供应起到了关键作用,生物态的再生铁和颗粒铁的再矿化共同维持了垂直剖面中的铁浓度.再次,生物态铁的来源或补充得益于铁池的生物利用,亚铁通过光化学方法产生,三价铁的获取则依靠高亲和力的铁还原酶系统来捕获有机铁,并且该机制普遍存在于贫铁环境的生物体中[8 ,9 ] .对于与浮游植物共生菌,特别是玫瑰杆菌来说,直接摄取血红素可能是一种有效的铁获取途径[23 ] .在高丰度浮游植物条件下,异养菌对血红素的直接转运可能是有机铁颗粒再循环进入微食物环的重要途径[23 ] . ...
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
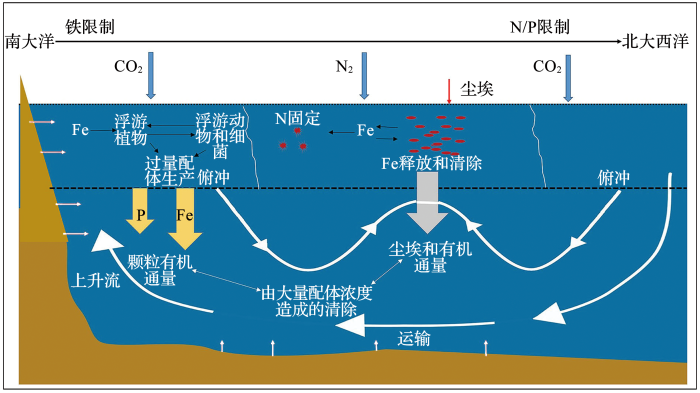
... 总结海洋铁循环的运作方式及其与N/C/P循环的关系,我们可以看到低纬度地区中尘埃铁的主导地位与高纬度地区中沉积物和热液铁源的作用不同.高纬度地区的铁供应推动了海洋碳循环,而低纬度地区的尘埃供应有助于维持海洋的固氮存量.微量元素铁的分布相比主导元素更为复杂,其中的驱动因素有深海铁源、颗粒铁沉降、铁的再生与清除、不同水团的影响以及远程运输的配体特性等.综合现有知识,Tagliabue等[8 ] 勾勒了一幅海洋铁循环示意图(图7 ).在高纬度地区,热液和沉积物是铁的主要来源[55 ] .混有铁元素的水流和混合作用是促进生物活性的主要基础,其通过微生物介导的铁再生来维持.高纬度地区的铁通常有限,因此配体以及溶解态络合物可能会潜沉和朝赤道方向输送(图7 ).相反,低纬度地区,大气沉降构成了铁的主要来源,如果尘埃供应量低同时上升流消失,随之而来的铁胁迫会导致固氮率降低[57 ] .低纬度地区的铁受到沉降有机物中再生铁与成岩颗粒铁之间的平衡关系的影响[58 ] .铁的清除率与未络合铁的数量密切相关,低纬度地区的铁可能会受到含有饱和配体的高纬度水的俯冲和赤道流的远程控制[59 ] .这一观点符合太平洋和大西洋的情形,因为两者的经向环流均以高纬度深水团为主.目前,针对铁的精细循环有过多种模型,但不同算法和维度的结果差异甚大,这些差异导致计算出的铁停留时间范围不一致,从几年到几百年不等,这种不确定性干扰了对铁的深度预测.为了减少这种不确定性,需要从新的海洋调查数据中提取更多的铁循环过程和控制因素等信息,以代入更精确的运算模型,求取外部铁供应和内部铁循环的周转时间. ...
... 海洋中铁循环的主要进程(据参考文献[
8 ]修改)
<strong>Representation of the major processes in the ocean iron cycle </strong>(<strong>modified after reference </strong>[<xref ref-type="bibr" rid="R8">8</xref>]) Fig.7 ![]()
5 展 望 得益于技术的发展和研究的深入,我们对海洋铁分布的认识已取得重大进展.然而,需要指出的是,现今对铁分布的变化以及与生物地球化学循环的联系方面的认识依旧比较零散,需要将铁研究整合到一个综合的“化学—生物—物理”理论框架中,该框架将是未来我们深入认识铁及其生态功能的依据.未来的工作中需要加强以下方面的研究: ...
... [
8 ])
Fig.7 ![]()
5 展 望 得益于技术的发展和研究的深入,我们对海洋铁分布的认识已取得重大进展.然而,需要指出的是,现今对铁分布的变化以及与生物地球化学循环的联系方面的认识依旧比较零散,需要将铁研究整合到一个综合的“化学—生物—物理”理论框架中,该框架将是未来我们深入认识铁及其生态功能的依据.未来的工作中需要加强以下方面的研究: ...
... (1)生物监测与物理观测的耦合,加强以稳定同位素为代表的示踪量化.以往采用了3 He来识别大洋中脊的铁羽流[60 ] ,并推导出铁和氦的热液通量关系.此外,在一些“GEOTRACES”区段上测定了镭的衰变[61 ] ,并且可将其与溶解铁数据集相关联,估算出了铁边界数值.未来的研究中,稳定同位素法结合海洋涡旋的测定是一个重要方向,可以用于确定铁的浓度在传输间的动态变化,还可以有效规避远程信号与交错过程的信号混杂[8 ] .在这种情形下铁的分布特征具有“指纹化”,可用来追踪铁的循环路径[8 ] .此外,结合物理场景(水团、密度或涡旋)的考虑,还有助于定量关键过程的速率,并建立铁与主要营养素之间的偶联或去偶联关系,有利于全球模型参数化的统一. ...
... [8 ].此外,结合物理场景(水团、密度或涡旋)的考虑,还有助于定量关键过程的速率,并建立铁与主要营养素之间的偶联或去偶联关系,有利于全球模型参数化的统一. ...
Iron biogeochemistry in aquatic systems: From source to bioavailability
13
2014
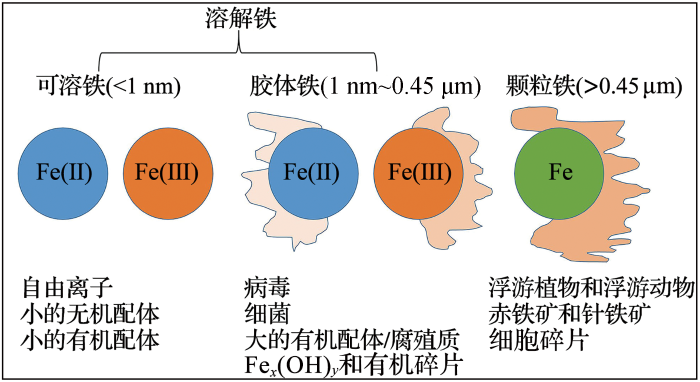
... 海水中铁的存在形式主要有溶解态、胶体态和颗粒态3种,铁的赋存形态受自身氧化还原状态和多种有机配体络合物的共同影响(图1 )[9 ,10 ] .溶解性无机铁主要有Fe(Ⅱ)和Fe(Ⅲ) 2种形式,Fe(Ⅱ)在含氧环境中很不稳定,易被氧化成Fe(Ⅲ).Fe(Ⅲ)主要以Fe(OH)2 + 2+ 2种水合形式存在,而Fe(Ⅲ)自由离子的浓度很低,一般仅为0.1~0.7 nmol/L[11 ] ,大洋环境则更低(10-3 nmol/L)[12 ] .胶体状态中,纯净的胶体铁并不常见,大部分都含有有机基质.海洋中的胶体铁有3种可能的形成机制:一是铁的无机沉淀物凝聚在有机基质中;二是无机Fe(Ⅲ)的水解物吸附在胶体表面或者与胶体发生络合反应;三是由有机Fe(Ⅲ)络合物与胶体缔合而成[11 ] .浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
... 海洋中铁的粒径、种类及相关生物示意图(据参考文献[
9 ]修改)
<strong>The various size fractions</strong>,<strong> species</strong>,<strong> and associated biology of Fe that exist in marine waters </strong>(<strong>modified after reference </strong>[<xref ref-type="bibr" rid="R9">9</xref>]) Fig.1 ![]()
相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
... [
9 ])
Fig.1 ![]()
相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
... 对铁赋存状态的理解,除了关注胶体铁与配体之间的络合程度或者颗粒铁的可溶性与可矿性之外,更需关注生物作用对铁的影响.由于铁的稀缺性和理化属性,铁的加工与转化和生物类群紧密相关(图2 )[8 ] .例如,浮游植物和细菌是小型微粒铁池的主要贡献者[20 ] ,而病毒则是构成胶体铁的重要组成部分[21 ] .另外,与金属酶结合的胞内铁在微生物溶解和病毒裂解之后会释放到外周环境中[22 ] .这些释放的胞内铁有助于环境胶体铁池内金属酶、铁结合组分(血红素和铁硫簇)以及储铁蛋白活力的发挥[4 ,22 ] .虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
... 除了“化学态”铁源,“生物态铁”亦是铁的另一大源泉,这种形式的铁源有3个特征.首先,相对碳、氮、磷而言,铁的化学计量值变化幅度更大.例如,南大洋低铁水域浮游植物中的Fe/P化学计量比通常不足高铁水域的1/5[9 ] ,这种化学计量的可塑性超出了主要营养素的常见水平,但对铁的生物循环却至关重要.其次,浮游动物、细菌和病毒作用下的铁再生对铁的生物供应起到了关键作用,生物态的再生铁和颗粒铁的再矿化共同维持了垂直剖面中的铁浓度.再次,生物态铁的来源或补充得益于铁池的生物利用,亚铁通过光化学方法产生,三价铁的获取则依靠高亲和力的铁还原酶系统来捕获有机铁,并且该机制普遍存在于贫铁环境的生物体中[8 ,9 ] .对于与浮游植物共生菌,特别是玫瑰杆菌来说,直接摄取血红素可能是一种有效的铁获取途径[23 ] .在高丰度浮游植物条件下,异养菌对血红素的直接转运可能是有机铁颗粒再循环进入微食物环的重要途径[23 ] . ...
... ,9 ].对于与浮游植物共生菌,特别是玫瑰杆菌来说,直接摄取血红素可能是一种有效的铁获取途径[23 ] .在高丰度浮游植物条件下,异养菌对血红素的直接转运可能是有机铁颗粒再循环进入微食物环的重要途径[23 ] . ...
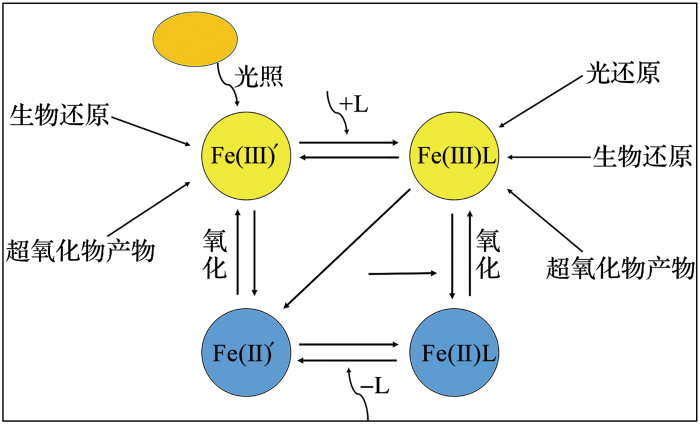
... 微生物介导的铁氧化过程主要包括4类:即O2 氧化、生物矿化、光化学过程以及硝酸盐氧化(图3 )[9 ,26 ] . ...
... 铁氧化还原循环示意图(据参考文献[
9 ]修改)
<strong>Representation of the Fe redox cycle </strong>(<strong>modified after reference </strong>[<xref ref-type="bibr" rid="R9">9</xref>]) Fe(Ⅲ)':无机三价铁;Fe(Ⅱ)':无机二价铁 ...
... [
9 ])
Fe(Ⅲ)':无机三价铁;Fe(Ⅱ)':无机二价铁 ...
... 铁的还原过程复杂而多样(图3 ),包括胶体铁的还原、铁—有机配体的光还原或生物还原[9 ] .铁在海洋中主要以无机态Fe(III)΄或者绑定有机配体的Fe(III)L形式存在,其中Fe(III)L是主要的存在形式(>99% [11 ] . ...
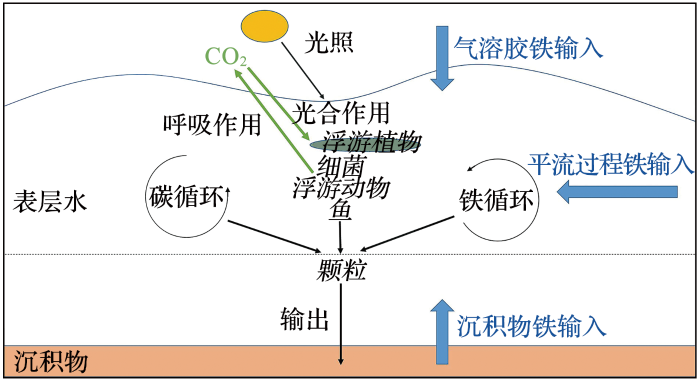
... 除了氮,铁与碳元素的关系也被关注,浮游植物的光合作用是固定大气CO2 的主要方式,将无机碳转化为有机碳是食物网碳循环的主要推手,铁的利用与再生为该网络的运行提供了驱动力(图6 )[9 ] .浮游植物、细菌和病毒的铁再生作用在铁的生物供应中扮演着“中转站”的作用[52 ] .有研究发现,再生铁有利于提升浮游植物的固碳效率,铁的吸收对于固碳速率有着根本的影响[24 ] .加利福尼亚海域44个站点设计的铁添加实验,证明了铁限制浮游植物的生长,主要因为钝化了体内有机碳的合成[53 ] . ...
... 铁循环和碳循环关系示意图(据参考文献[
9 ]修改)
<strong>Schematic of the links between iron </strong>(<strong>Fe</strong>)<strong> and carbon </strong>(<strong>C</strong>)<strong> cycling </strong>(<strong>modified after reference </strong>[<xref ref-type="bibr" rid="R9">9</xref>]) 蓝色表示铁输入,绿色表示碳过程,斜体表示生物相互作用 ...
... [
9 ])
蓝色表示铁输入,绿色表示碳过程,斜体表示生物相互作用 ...
Iron plays a role in nitrate drawdown by phytoplankton in Lake Erie surface waters as observed in lake-wide assessments
1
2012
... 海水中铁的存在形式主要有溶解态、胶体态和颗粒态3种,铁的赋存形态受自身氧化还原状态和多种有机配体络合物的共同影响(图1 )[9 ,10 ] .溶解性无机铁主要有Fe(Ⅱ)和Fe(Ⅲ) 2种形式,Fe(Ⅱ)在含氧环境中很不稳定,易被氧化成Fe(Ⅲ).Fe(Ⅲ)主要以Fe(OH)2 + 2+ 2种水合形式存在,而Fe(Ⅲ)自由离子的浓度很低,一般仅为0.1~0.7 nmol/L[11 ] ,大洋环境则更低(10-3 nmol/L)[12 ] .胶体状态中,纯净的胶体铁并不常见,大部分都含有有机基质.海洋中的胶体铁有3种可能的形成机制:一是铁的无机沉淀物凝聚在有机基质中;二是无机Fe(Ⅲ)的水解物吸附在胶体表面或者与胶体发生络合反应;三是由有机Fe(Ⅲ)络合物与胶体缔合而成[11 ] .浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
Iron sources, existing forms and their limiting action on the primary productivity of phytoplankton in seawater
3
1998
... 海水中铁的存在形式主要有溶解态、胶体态和颗粒态3种,铁的赋存形态受自身氧化还原状态和多种有机配体络合物的共同影响(图1 )[9 ,10 ] .溶解性无机铁主要有Fe(Ⅱ)和Fe(Ⅲ) 2种形式,Fe(Ⅱ)在含氧环境中很不稳定,易被氧化成Fe(Ⅲ).Fe(Ⅲ)主要以Fe(OH)2 + 2+ 2种水合形式存在,而Fe(Ⅲ)自由离子的浓度很低,一般仅为0.1~0.7 nmol/L[11 ] ,大洋环境则更低(10-3 nmol/L)[12 ] .胶体状态中,纯净的胶体铁并不常见,大部分都含有有机基质.海洋中的胶体铁有3种可能的形成机制:一是铁的无机沉淀物凝聚在有机基质中;二是无机Fe(Ⅲ)的水解物吸附在胶体表面或者与胶体发生络合反应;三是由有机Fe(Ⅲ)络合物与胶体缔合而成[11 ] .浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
... [11 ].浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
... 铁的还原过程复杂而多样(图3 ),包括胶体铁的还原、铁—有机配体的光还原或生物还原[9 ] .铁在海洋中主要以无机态Fe(III)΄或者绑定有机配体的Fe(III)L形式存在,其中Fe(III)L是主要的存在形式(>99% [11 ] . ...
海洋中铁的来源、形态和对初级生产力的限制作用
3
1998
... 海水中铁的存在形式主要有溶解态、胶体态和颗粒态3种,铁的赋存形态受自身氧化还原状态和多种有机配体络合物的共同影响(图1 )[9 ,10 ] .溶解性无机铁主要有Fe(Ⅱ)和Fe(Ⅲ) 2种形式,Fe(Ⅱ)在含氧环境中很不稳定,易被氧化成Fe(Ⅲ).Fe(Ⅲ)主要以Fe(OH)2 + 2+ 2种水合形式存在,而Fe(Ⅲ)自由离子的浓度很低,一般仅为0.1~0.7 nmol/L[11 ] ,大洋环境则更低(10-3 nmol/L)[12 ] .胶体状态中,纯净的胶体铁并不常见,大部分都含有有机基质.海洋中的胶体铁有3种可能的形成机制:一是铁的无机沉淀物凝聚在有机基质中;二是无机Fe(Ⅲ)的水解物吸附在胶体表面或者与胶体发生络合反应;三是由有机Fe(Ⅲ)络合物与胶体缔合而成[11 ] .浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
... [11 ].浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
... 铁的还原过程复杂而多样(图3 ),包括胶体铁的还原、铁—有机配体的光还原或生物还原[9 ] .铁在海洋中主要以无机态Fe(III)΄或者绑定有机配体的Fe(III)L形式存在,其中Fe(III)L是主要的存在形式(>99% [11 ] . ...
The eastern extent of seasonal iron limitation in the high latitude North Atlantic Ocean
1
2019
... 海水中铁的存在形式主要有溶解态、胶体态和颗粒态3种,铁的赋存形态受自身氧化还原状态和多种有机配体络合物的共同影响(图1 )[9 ,10 ] .溶解性无机铁主要有Fe(Ⅱ)和Fe(Ⅲ) 2种形式,Fe(Ⅱ)在含氧环境中很不稳定,易被氧化成Fe(Ⅲ).Fe(Ⅲ)主要以Fe(OH)2 + 2+ 2种水合形式存在,而Fe(Ⅲ)自由离子的浓度很低,一般仅为0.1~0.7 nmol/L[11 ] ,大洋环境则更低(10-3 nmol/L)[12 ] .胶体状态中,纯净的胶体铁并不常见,大部分都含有有机基质.海洋中的胶体铁有3种可能的形成机制:一是铁的无机沉淀物凝聚在有机基质中;二是无机Fe(Ⅲ)的水解物吸附在胶体表面或者与胶体发生络合反应;三是由有机Fe(Ⅲ)络合物与胶体缔合而成[11 ] .浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
Ability of marine eukaryotic red tide microalgae to utilize insoluble iron
1
2005
... 海水中铁的存在形式主要有溶解态、胶体态和颗粒态3种,铁的赋存形态受自身氧化还原状态和多种有机配体络合物的共同影响(图1 )[9 ,10 ] .溶解性无机铁主要有Fe(Ⅱ)和Fe(Ⅲ) 2种形式,Fe(Ⅱ)在含氧环境中很不稳定,易被氧化成Fe(Ⅲ).Fe(Ⅲ)主要以Fe(OH)2 + 2+ 2种水合形式存在,而Fe(Ⅲ)自由离子的浓度很低,一般仅为0.1~0.7 nmol/L[11 ] ,大洋环境则更低(10-3 nmol/L)[12 ] .胶体状态中,纯净的胶体铁并不常见,大部分都含有有机基质.海洋中的胶体铁有3种可能的形成机制:一是铁的无机沉淀物凝聚在有机基质中;二是无机Fe(Ⅲ)的水解物吸附在胶体表面或者与胶体发生络合反应;三是由有机Fe(Ⅲ)络合物与胶体缔合而成[11 ] .浮游植物除了利用铁的自由离子之外,还可以利用由胶体Fe(Ⅲ)转化而来的活性铁离子.铁的第三种形态——颗粒铁,对于生物而言不可直接利用,但颗粒铁却扮演了“铁库”的角色,藻类和细菌在特定的条件下,可以释放增溶物质(生物酶、电子受体)将颗粒铁转化为溶解铁.在溶解铁不足的情况下,一些海区也会看到藻类的爆发性增殖.Naito等[13 ] 研究认为,海洋真核微藻具有利用不溶铁的能力,部分微藻可以利用FePO4 和FeS颗粒进行生长,藻类生物量的高低可能取决于自身种类和不溶性铁类型的组合. ...
Multiple siderophores: Bug or feature?
1
2018
... 相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
Endocytosis-mediated siderophore uptake as a strategy for Fe acquisition in diatoms
1
2018
... 相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
Iron Cycle in Oceans
2
2016
... 相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
... [16 ].EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
Domoic acid: The synergy of iron, copper, and the toxicity of diatoms
1
2005
... 相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
Microbial Extracellular Polymeric Substances (EPSs) in ocean systems
1
2017
... 相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
Saccharides enhance iron bioavailability to Southern Ocean phytoplankton
1
2011
... 相比于无机铁,有机铁是一类更易以络合形式存在的类型.有机络合铁几乎是海水中溶解铁形态的普遍特征.常见的有机铁包括铁载体(siderophores)和卟啉(ferroporphyrin)等.铁载体是细菌从周围环境中富集铁的重要方式,能产生铁载体的细菌有多种,但它们对铁载体的吸收不具有严格的特异性[14 ] .在大西洋中最主要的一类铁载体为铁草胺(ferrioxamine),它占整个溶解铁池的0.5% % [15 ] .络合态有机铁中,卟啉是重要的一类,它包括叶绿素和叶绿素分解产物,如褐藻素、亚铁血红素和维生素B12 (一种潜在的铁配体)[16 ] .软骨藻酸(domoic acid)是拟菱形藻(Pseudonitzschia sp.)产生的一种可以络合铁的藻毒素.研究发现,在铁限制环境中软骨藻酸对促进甲藻吸收高亲和力铁具有重要作用[17 ] .对于配体型有机铁,有报道显示腐殖质(Humus, HS)和胞外多糖(Exopolysaccharides, EPS)络合铁是铁池的重要组成部分,HS在沿海和深海中均有存在,可以占配体总量的一半以上[16 ] .EPS中含有大量酸性多糖,如尿酸,它可以络合铁且pH值越高形成的络合体越稳定,这一点与Fe-HS络合体相似[18 ] .有研究表明,与铁—铁载体配合物相比,EPS提高了铁与其他配体的稳定性,同时也提高了铁在浮游植物中的可利用性[19 ] . ...
The role of heterotrophic bacteria in iron-limited ocean ecosystems
5
1996
... 对铁赋存状态的理解,除了关注胶体铁与配体之间的络合程度或者颗粒铁的可溶性与可矿性之外,更需关注生物作用对铁的影响.由于铁的稀缺性和理化属性,铁的加工与转化和生物类群紧密相关(图2 )[8 ] .例如,浮游植物和细菌是小型微粒铁池的主要贡献者[20 ] ,而病毒则是构成胶体铁的重要组成部分[21 ] .另外,与金属酶结合的胞内铁在微生物溶解和病毒裂解之后会释放到外周环境中[22 ] .这些释放的胞内铁有助于环境胶体铁池内金属酶、铁结合组分(血红素和铁硫簇)以及储铁蛋白活力的发挥[4 ,22 ] .虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
... 微生物在铁循环中扮演着重要的角色,这源于微生物具有较大的铁配额(molFe/cell或μmolFe/molC),使得大部分生物铁包含在细菌中.评估生物铁池的大小需要估计生态系统中生产者和消费者的生物量以及铁配额比例.海洋原核生物比真核藻类有更高的铁配额,在铁限制的条件下,硅藻的铁配额为(3.0±1.5) μmolFe/molC,而异养细菌的铁配额达到(7.5±1.7) μmolFe/molC[20 ] .野外调查的数据也支持这一结论,在亚北极太平洋异养细菌和浮游植物的铁配额分别为(6.1±2.5) μmolFe/molC和(3.7±2.3) μmolFe/molC[20 ,24 ] .海洋细菌的高铁配额与其自身生理需求有关.例如,自养的光合细菌在光合作用中需要铁的参与,异养细菌在电子传递链中也需要大量的铁作为氧化还原催化剂.Tortell等[20 ] 计算了亚北极太平洋浮游植物、蓝藻和异养细菌的铁含量(生物量与铁配额的乘积).结果表明:整个生物铁含量中细菌占80% [20 ] . ...
... [20 ,24 ].海洋细菌的高铁配额与其自身生理需求有关.例如,自养的光合细菌在光合作用中需要铁的参与,异养细菌在电子传递链中也需要大量的铁作为氧化还原催化剂.Tortell等[20 ] 计算了亚北极太平洋浮游植物、蓝藻和异养细菌的铁含量(生物量与铁配额的乘积).结果表明:整个生物铁含量中细菌占80% [20 ] . ...
... [20 ]计算了亚北极太平洋浮游植物、蓝藻和异养细菌的铁含量(生物量与铁配额的乘积).结果表明:整个生物铁含量中细菌占80% [20 ] . ...
... [20 ]. ...
The ferrojan horse hypothesis: Iron-virus interactions in the ocean
1
2016
... 对铁赋存状态的理解,除了关注胶体铁与配体之间的络合程度或者颗粒铁的可溶性与可矿性之外,更需关注生物作用对铁的影响.由于铁的稀缺性和理化属性,铁的加工与转化和生物类群紧密相关(图2 )[8 ] .例如,浮游植物和细菌是小型微粒铁池的主要贡献者[20 ] ,而病毒则是构成胶体铁的重要组成部分[21 ] .另外,与金属酶结合的胞内铁在微生物溶解和病毒裂解之后会释放到外周环境中[22 ] .这些释放的胞内铁有助于环境胶体铁池内金属酶、铁结合组分(血红素和铁硫簇)以及储铁蛋白活力的发挥[4 ,22 ] .虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
Divergent responses of Atlantic coastal and oceanic Synechococcus to iron limitation
2
2015
... 对铁赋存状态的理解,除了关注胶体铁与配体之间的络合程度或者颗粒铁的可溶性与可矿性之外,更需关注生物作用对铁的影响.由于铁的稀缺性和理化属性,铁的加工与转化和生物类群紧密相关(图2 )[8 ] .例如,浮游植物和细菌是小型微粒铁池的主要贡献者[20 ] ,而病毒则是构成胶体铁的重要组成部分[21 ] .另外,与金属酶结合的胞内铁在微生物溶解和病毒裂解之后会释放到外周环境中[22 ] .这些释放的胞内铁有助于环境胶体铁池内金属酶、铁结合组分(血红素和铁硫簇)以及储铁蛋白活力的发挥[4 ,22 ] .虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
... ,22 ].虽然具体的贡献大小还有待确认,但对整个胞内和胞外的铁生物学过程具有不可忽视的作用.此外,海洋铁池不同成分的瞬态性质和环境影响,也是未来关注的重点[9 ] . ...
Direct heme uptake by phytoplankton-associated roseobacter bacteria
2
2017
... 除了“化学态”铁源,“生物态铁”亦是铁的另一大源泉,这种形式的铁源有3个特征.首先,相对碳、氮、磷而言,铁的化学计量值变化幅度更大.例如,南大洋低铁水域浮游植物中的Fe/P化学计量比通常不足高铁水域的1/5[9 ] ,这种化学计量的可塑性超出了主要营养素的常见水平,但对铁的生物循环却至关重要.其次,浮游动物、细菌和病毒作用下的铁再生对铁的生物供应起到了关键作用,生物态的再生铁和颗粒铁的再矿化共同维持了垂直剖面中的铁浓度.再次,生物态铁的来源或补充得益于铁池的生物利用,亚铁通过光化学方法产生,三价铁的获取则依靠高亲和力的铁还原酶系统来捕获有机铁,并且该机制普遍存在于贫铁环境的生物体中[8 ,9 ] .对于与浮游植物共生菌,特别是玫瑰杆菌来说,直接摄取血红素可能是一种有效的铁获取途径[23 ] .在高丰度浮游植物条件下,异养菌对血红素的直接转运可能是有机铁颗粒再循环进入微食物环的重要途径[23 ] . ...
... [23 ]. ...
Utilization of iron bound to strong organic ligands by plankton communities in the subarctic Pacific Ocean
3
1999
... 微生物在铁循环中扮演着重要的角色,这源于微生物具有较大的铁配额(molFe/cell或μmolFe/molC),使得大部分生物铁包含在细菌中.评估生物铁池的大小需要估计生态系统中生产者和消费者的生物量以及铁配额比例.海洋原核生物比真核藻类有更高的铁配额,在铁限制的条件下,硅藻的铁配额为(3.0±1.5) μmolFe/molC,而异养细菌的铁配额达到(7.5±1.7) μmolFe/molC[20 ] .野外调查的数据也支持这一结论,在亚北极太平洋异养细菌和浮游植物的铁配额分别为(6.1±2.5) μmolFe/molC和(3.7±2.3) μmolFe/molC[20 ,24 ] .海洋细菌的高铁配额与其自身生理需求有关.例如,自养的光合细菌在光合作用中需要铁的参与,异养细菌在电子传递链中也需要大量的铁作为氧化还原催化剂.Tortell等[20 ] 计算了亚北极太平洋浮游植物、蓝藻和异养细菌的铁含量(生物量与铁配额的乘积).结果表明:整个生物铁含量中细菌占80% [20 ] . ...
... 在酸性或近中性环境中微生物都可以利用O2 氧化Fe(Ⅱ),前者主要是亚硫铁杆菌介导的生物氧化[24 ] ,而后者主要是化学氧化.相比于酸性环境,中性环境中Fe(Ⅱ)更易被O2 快速氧化,与生物氧化过程形成竞争[27 ] ;但在低氧(<50 μmol/L)环境中,微氧型亚铁氧化菌可以抗衡O2 ,其所介导的生物氧化过程占主导.除此之外,微生物产生的氧自由基(O2 ·)也可以调节铁的氧化还原过程,O2 ·可以将 Fe(Ⅱ)氧化为Fe(Ⅲ)且产生H2 O2 [27 ] .Fe(Ⅲ)配合物可以直接或间接地与O2 ·发生还原反应,且Fe(Ⅲ)的还原速率高于Fe(Ⅱ)的氧化速率,在此期间O2 ·主要起到了介导Fe(Ⅲ)还原的作用[27 ] . ...
... 除了氮,铁与碳元素的关系也被关注,浮游植物的光合作用是固定大气CO2 的主要方式,将无机碳转化为有机碳是食物网碳循环的主要推手,铁的利用与再生为该网络的运行提供了驱动力(图6 )[9 ] .浮游植物、细菌和病毒的铁再生作用在铁的生物供应中扮演着“中转站”的作用[52 ] .有研究发现,再生铁有利于提升浮游植物的固碳效率,铁的吸收对于固碳速率有着根本的影响[24 ] .加利福尼亚海域44个站点设计的铁添加实验,证明了铁限制浮游植物的生长,主要因为钝化了体内有机碳的合成[53 ] . ...
What controls dissolved iron concentrations in the world ocean?
1
1997
... 微生物铁配额的存在一方面维持了其占据较高比例的铁拥有量,另一方面也有利于Fe浓度的相对稳定.一些活泼金属如Al,Mn和Pb等由于颗粒吸附作用会随着水深的增加而出现溶解度下降;但是在深海(>500 m)环境中,溶解铁浓度却保持相对稳定,这主要是由于细菌配额中Fe(Ⅲ)有机配体的存在,可以提高胶体铁的溶解度并降低被沉降的几率[25 ] . ...
The role of microorganisms in the geochemical iron cycle
1
2016
... 微生物介导的铁氧化过程主要包括4类:即O2 氧化、生物矿化、光化学过程以及硝酸盐氧化(图3 )[9 ,26 ] . ...
微生物在地球化学铁循环过程中的作用
1
2016
... 微生物介导的铁氧化过程主要包括4类:即O2 氧化、生物矿化、光化学过程以及硝酸盐氧化(图3 )[9 ,26 ] . ...
Iron metabolism in anoxic environments at near neutral pH
3
2000
... 在酸性或近中性环境中微生物都可以利用O2 氧化Fe(Ⅱ),前者主要是亚硫铁杆菌介导的生物氧化[24 ] ,而后者主要是化学氧化.相比于酸性环境,中性环境中Fe(Ⅱ)更易被O2 快速氧化,与生物氧化过程形成竞争[27 ] ;但在低氧(<50 μmol/L)环境中,微氧型亚铁氧化菌可以抗衡O2 ,其所介导的生物氧化过程占主导.除此之外,微生物产生的氧自由基(O2 ·)也可以调节铁的氧化还原过程,O2 ·可以将 Fe(Ⅱ)氧化为Fe(Ⅲ)且产生H2 O2 [27 ] .Fe(Ⅲ)配合物可以直接或间接地与O2 ·发生还原反应,且Fe(Ⅲ)的还原速率高于Fe(Ⅱ)的氧化速率,在此期间O2 ·主要起到了介导Fe(Ⅲ)还原的作用[27 ] . ...
... [27 ].Fe(Ⅲ)配合物可以直接或间接地与O2 ·发生还原反应,且Fe(Ⅲ)的还原速率高于Fe(Ⅱ)的氧化速率,在此期间O2 ·主要起到了介导Fe(Ⅲ)还原的作用[27 ] . ...
... [27 ]. ...
Iron-oxidizing bacteria in marine environments: Recent progresses and future directions
1
2018
... 参与铁氧化的细菌大部分属于微氧型亚铁氧化菌,少量来自δ-变形菌纲,大部分来自β-变形菌纲的披毛菌属(Gallionella )与纤毛菌属(Leptothrix )[28 ] .披毛菌属的代表为G. ferruginea ,营无机化能营养,属于亚硝化单胞菌目(Nitrosomonadales),需要足够的营养供给,其氧化Fe(Ⅱ)的产物为不溶性的氢氧化铁[29 ] .纤毛菌属的代表为L.discophora ,属于伯克氏菌目(Burkholderiales),与其他微氧型亚铁氧化菌不同的是,除了铁之外它还可以氧化锰[30 ] . ...
The role of iron-oxidizing bacteria in biocorrosion: A review
1
2018
... 参与铁氧化的细菌大部分属于微氧型亚铁氧化菌,少量来自δ-变形菌纲,大部分来自β-变形菌纲的披毛菌属(Gallionella )与纤毛菌属(Leptothrix )[28 ] .披毛菌属的代表为G. ferruginea ,营无机化能营养,属于亚硝化单胞菌目(Nitrosomonadales),需要足够的营养供给,其氧化Fe(Ⅱ)的产物为不溶性的氢氧化铁[29 ] .纤毛菌属的代表为L.discophora ,属于伯克氏菌目(Burkholderiales),与其他微氧型亚铁氧化菌不同的是,除了铁之外它还可以氧化锰[30 ] . ...
Production of biogenic Mn oxides by leptothrix discophora SS-1 in a chemically defined growth medium and evaluation of their Pb adsorption characteristics
1
1999
... 参与铁氧化的细菌大部分属于微氧型亚铁氧化菌,少量来自δ-变形菌纲,大部分来自β-变形菌纲的披毛菌属(Gallionella )与纤毛菌属(Leptothrix )[28 ] .披毛菌属的代表为G. ferruginea ,营无机化能营养,属于亚硝化单胞菌目(Nitrosomonadales),需要足够的营养供给,其氧化Fe(Ⅱ)的产物为不溶性的氢氧化铁[29 ] .纤毛菌属的代表为L.discophora ,属于伯克氏菌目(Burkholderiales),与其他微氧型亚铁氧化菌不同的是,除了铁之外它还可以氧化锰[30 ] . ...
Iron-oxidizing bacteria in marine environments: Recent progresses and future directions
3
2018
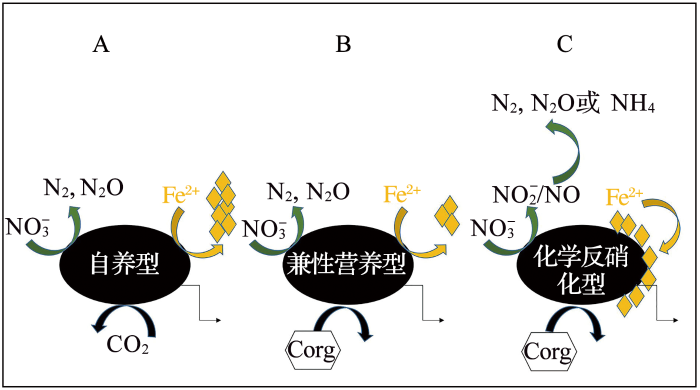
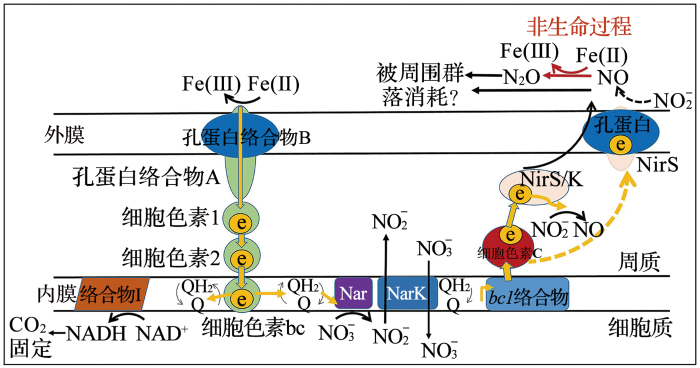
... 在厌氧条件下,厌氧铁氧化菌是氧化Fe(Ⅱ)的关键成员,能将硝酸根还原为亚硝酸根,利用电子供体将硝酸根还原为NO,N2 O和N2 ,在此过程中同时进行Fe(Ⅱ)的氧化[31 ] .该反应过程依赖于硝酸盐的还原,需亚铁氧化与硝酸根耦合,从而使亚铁氧化菌参与到氮循环中.厌氧硝化菌是另一种主要的铁氧化菌,其作用方式与细菌的自养特性、氢营养类型以及是否属于化学反硝化菌有关.厌氧硝化菌主要有3种类型(图4 ):① 2 获得碳,通过Fe(II)氧化还原酶氧化Fe(II);② ③ [32 ] .图5 展示了自养型Fe(II)氧化剂氧化Fe(II)的潜在机理,细菌通过Fe(II)氧化获得一个电子,并沿电子传递链传递,硝酸盐逐步还原为NO;在胞外,NO有可能被周围群落消耗或与水合Fe(II)反应[32 ] . ...
... 常见的硝酸盐依赖型厌氧铁氧化菌有嗜酸菌、嗜热古菌、脱氮硫杆菌等,分属于α-,β-,γ-及δ-变形菌纲[31 ] .Acidovorax纲的BoFeN1作为模式菌株以Fe(Ⅱ)与醋酸盐为电子供体、NO3 - 3 - . 醋酸盐的存在是因为Fe(Ⅱ)作为唯一电子供体难以维持菌株BoFeN1的连续传代培养,1 mmol/L的醋酸盐可以辅助4 mmol/L Fe(Ⅱ)氧化为Fe(Ⅲ)[33 ] .从沉积物中得到的脱氯单胞菌Dechloromonas sp. UWNR4具有亚铁氧化耦合硝酸盐还原的能力,而脱氯单胞菌属具有高氯酸还原能力,这表明亚铁氧化可能耦合氯元素循环[34 ] . ...
... 光化学过程的氧化行为,现今主要报道的是厌氧光合铁氧化菌,这类细菌在厌氧条件下,以光为能源、碳酸氢盐为电子受体、Fe(Ⅱ)为唯一电子供体生成Fe(OH)3 ,合成生物量的同时氧化Fe(Ⅱ).光合型铁氧化菌绝大多数属于α变形菌纲,适宜生活在pH为6.5~7.0,主要的代表性微生物有红杆菌科(Rhodobacteraceae),可以进行有氧或兼性厌氧的代谢方式[31 ] .基因组研究进一步揭示了该菌的另一个特点:氧化Fe(Ⅱ)和固定碳的形式要先于氧气型光合作用[36 ] . ...
Microbial anaerobic Fe(II) oxidation-ecology, mechanisms and environmental implications
6
2018
... 在厌氧条件下,厌氧铁氧化菌是氧化Fe(Ⅱ)的关键成员,能将硝酸根还原为亚硝酸根,利用电子供体将硝酸根还原为NO,N2 O和N2 ,在此过程中同时进行Fe(Ⅱ)的氧化[31 ] .该反应过程依赖于硝酸盐的还原,需亚铁氧化与硝酸根耦合,从而使亚铁氧化菌参与到氮循环中.厌氧硝化菌是另一种主要的铁氧化菌,其作用方式与细菌的自养特性、氢营养类型以及是否属于化学反硝化菌有关.厌氧硝化菌主要有3种类型(图4 ):① 2 获得碳,通过Fe(II)氧化还原酶氧化Fe(II);② ③ [32 ] .图5 展示了自养型Fe(II)氧化剂氧化Fe(II)的潜在机理,细菌通过Fe(II)氧化获得一个电子,并沿电子传递链传递,硝酸盐逐步还原为NO;在胞外,NO有可能被周围群落消耗或与水合Fe(II)反应[32 ] . ...
... [32 ]. ...
... [
32 ]
<strong>Overview of three different types of interaction between nitrate-reducing bacteria and Fe</strong>(<strong>II</strong>)<sup>[<xref ref-type="bibr" rid="R32">32</xref>]</sup> Fig.4 ![]()
图5 关于<strong>Fe</strong>(<strong>II</strong>)氧化机理的假设示意图<sup>[<xref ref-type="bibr" rid="R32">32</xref>]</sup> <strong>Schematics of the current hypotheses on the mechanism of Fe</strong>(<strong>II</strong>)<strong> oxidation</strong><sup>[<xref ref-type="bibr" rid="R32">32</xref>]</sup> NADH:烟酰胺腺嘌呤二核苷酸(还原态),是还原型辅酶Ⅰ;NAD+ :烟酰胺腺嘌呤二核苷酸(氧化态),是辅酶Ⅰ ...
... [
32 ]
Fig.4 ![]()
图5 关于<strong>Fe</strong>(<strong>II</strong>)氧化机理的假设示意图<sup>[<xref ref-type="bibr" rid="R32">32</xref>]</sup> <strong>Schematics of the current hypotheses on the mechanism of Fe</strong>(<strong>II</strong>)<strong> oxidation</strong><sup>[<xref ref-type="bibr" rid="R32">32</xref>]</sup> NADH:烟酰胺腺嘌呤二核苷酸(还原态),是还原型辅酶Ⅰ;NAD+ :烟酰胺腺嘌呤二核苷酸(氧化态),是辅酶Ⅰ ...
... [
32 ]
<strong>Schematics of the current hypotheses on the mechanism of Fe</strong>(<strong>II</strong>)<strong> oxidation</strong><sup>[<xref ref-type="bibr" rid="R32">32</xref>]</sup> NADH:烟酰胺腺嘌呤二核苷酸(还原态),是还原型辅酶Ⅰ;NAD+ :烟酰胺腺嘌呤二核苷酸(氧化态),是辅酶Ⅰ ...
... [
32 ]
NADH:烟酰胺腺嘌呤二核苷酸(还原态),是还原型辅酶Ⅰ;NAD+ :烟酰胺腺嘌呤二核苷酸(氧化态),是辅酶Ⅰ ...
Fe(III) mineral formation and cell encrustation by the nitrate-dependent Fe(II)-oxidizer strain BoFeN1
1
2010
... 常见的硝酸盐依赖型厌氧铁氧化菌有嗜酸菌、嗜热古菌、脱氮硫杆菌等,分属于α-,β-,γ-及δ-变形菌纲[31 ] .Acidovorax纲的BoFeN1作为模式菌株以Fe(Ⅱ)与醋酸盐为电子供体、NO3 - 3 - . 醋酸盐的存在是因为Fe(Ⅱ)作为唯一电子供体难以维持菌株BoFeN1的连续传代培养,1 mmol/L的醋酸盐可以辅助4 mmol/L Fe(Ⅱ)氧化为Fe(Ⅲ)[33 ] .从沉积物中得到的脱氯单胞菌Dechloromonas sp. UWNR4具有亚铁氧化耦合硝酸盐还原的能力,而脱氯单胞菌属具有高氯酸还原能力,这表明亚铁氧化可能耦合氯元素循环[34 ] . ...
Neutrophilic, nitrate-dependent, Fe(II) oxidation by a dechloromonas species
1
2013
... 常见的硝酸盐依赖型厌氧铁氧化菌有嗜酸菌、嗜热古菌、脱氮硫杆菌等,分属于α-,β-,γ-及δ-变形菌纲[31 ] .Acidovorax纲的BoFeN1作为模式菌株以Fe(Ⅱ)与醋酸盐为电子供体、NO3 - 3 - . 醋酸盐的存在是因为Fe(Ⅱ)作为唯一电子供体难以维持菌株BoFeN1的连续传代培养,1 mmol/L的醋酸盐可以辅助4 mmol/L Fe(Ⅱ)氧化为Fe(Ⅲ)[33 ] .从沉积物中得到的脱氯单胞菌Dechloromonas sp. UWNR4具有亚铁氧化耦合硝酸盐还原的能力,而脱氯单胞菌属具有高氯酸还原能力,这表明亚铁氧化可能耦合氯元素循环[34 ] . ...
What's new is old: Resolving the identity of leptothrix ochracea using single cell genomics, pyrosequencing and fish
1
2011
... 相比于有氧时期,生物矿化主要出现在低氧或无氧环境,Fe(Ⅱ)的氧化主要依靠微生物来进行,微生物氧化Fe(Ⅱ)产生Fe(Ⅲ)矿物,一些厌氧细菌在氧化过程中可以产生矿化有机鞘来防止铁矿物的覆盖[35 ] . ...
Soil microbe mediated iron cycling and its environmental implication
3
2014
... 光化学过程的氧化行为,现今主要报道的是厌氧光合铁氧化菌,这类细菌在厌氧条件下,以光为能源、碳酸氢盐为电子受体、Fe(Ⅱ)为唯一电子供体生成Fe(OH)3 ,合成生物量的同时氧化Fe(Ⅱ).光合型铁氧化菌绝大多数属于α变形菌纲,适宜生活在pH为6.5~7.0,主要的代表性微生物有红杆菌科(Rhodobacteraceae),可以进行有氧或兼性厌氧的代谢方式[31 ] .基因组研究进一步揭示了该菌的另一个特点:氧化Fe(Ⅱ)和固定碳的形式要先于氧气型光合作用[36 ] . ...
... Fe(Ⅲ)还原通过异化铁还原菌氧化有机或无机物来进行.异化铁还原菌包含大多数的古菌和细菌,它们在厌氧环境中利用铁还原酶的催化,将电子在呼吸链上传递给末端电子受体Fe(Ⅲ)并产生其所需能量,因此异化铁还原又称铁呼吸[37 ] ,这种途径被认为是最古老的产能方式之一.异化铁还原菌采用3种方式将电子传递到不溶的Fe(Ⅲ)矿物表面:① ② ③ [36 ] . ...
... 地杆菌科(Geobacteraceae)和希瓦氏菌属(Shewanella )是研究最多的2种异化铁还原菌[40 ] .地杆菌属(Geobacter )可以氧化有机物,Fe(Ⅲ)是进行铁的异化还原的唯一电子受体[41 ] .G. sulfurreducens 的胞外电子传递主要通过C-型细胞色素OmcE和OmcS介导,其中OmcS与导电菌毛相连,在外膜孔蛋白OmpJ的帮助下,沿此路径调节电子的传递[36 ] .希瓦氏菌属中S. oneidensis MR-1研究较多,可以利用H+ 、甲酸盐和丙酮酸等进行异化铁还原.MR-1通过C-型细胞色素CymA、质膜C-型细胞色素MtrA、外膜MtrB以及外膜C-型细胞色素MtrC和OmcA来转移电子到Fe(Ⅲ),以此来进行异化铁的还原[42 ] . ...
土壤微生物铁循环及其环境意义
3
2014
... 光化学过程的氧化行为,现今主要报道的是厌氧光合铁氧化菌,这类细菌在厌氧条件下,以光为能源、碳酸氢盐为电子受体、Fe(Ⅱ)为唯一电子供体生成Fe(OH)3 ,合成生物量的同时氧化Fe(Ⅱ).光合型铁氧化菌绝大多数属于α变形菌纲,适宜生活在pH为6.5~7.0,主要的代表性微生物有红杆菌科(Rhodobacteraceae),可以进行有氧或兼性厌氧的代谢方式[31 ] .基因组研究进一步揭示了该菌的另一个特点:氧化Fe(Ⅱ)和固定碳的形式要先于氧气型光合作用[36 ] . ...
... Fe(Ⅲ)还原通过异化铁还原菌氧化有机或无机物来进行.异化铁还原菌包含大多数的古菌和细菌,它们在厌氧环境中利用铁还原酶的催化,将电子在呼吸链上传递给末端电子受体Fe(Ⅲ)并产生其所需能量,因此异化铁还原又称铁呼吸[37 ] ,这种途径被认为是最古老的产能方式之一.异化铁还原菌采用3种方式将电子传递到不溶的Fe(Ⅲ)矿物表面:① ② ③ [36 ] . ...
... 地杆菌科(Geobacteraceae)和希瓦氏菌属(Shewanella )是研究最多的2种异化铁还原菌[40 ] .地杆菌属(Geobacter )可以氧化有机物,Fe(Ⅲ)是进行铁的异化还原的唯一电子受体[41 ] .G. sulfurreducens 的胞外电子传递主要通过C-型细胞色素OmcE和OmcS介导,其中OmcS与导电菌毛相连,在外膜孔蛋白OmpJ的帮助下,沿此路径调节电子的传递[36 ] .希瓦氏菌属中S. oneidensis MR-1研究较多,可以利用H+ 、甲酸盐和丙酮酸等进行异化铁还原.MR-1通过C-型细胞色素CymA、质膜C-型细胞色素MtrA、外膜MtrB以及外膜C-型细胞色素MtrC和OmcA来转移电子到Fe(Ⅲ),以此来进行异化铁的还原[42 ] . ...
Influence of secondary iron-oxide mineralization induced by dissimilatory iron reduction bacteria on fraction transformation of heavy metals in soil
1
2018
... Fe(Ⅲ)还原通过异化铁还原菌氧化有机或无机物来进行.异化铁还原菌包含大多数的古菌和细菌,它们在厌氧环境中利用铁还原酶的催化,将电子在呼吸链上传递给末端电子受体Fe(Ⅲ)并产生其所需能量,因此异化铁还原又称铁呼吸[37 ] ,这种途径被认为是最古老的产能方式之一.异化铁还原菌采用3种方式将电子传递到不溶的Fe(Ⅲ)矿物表面:① ② ③ [36 ] . ...
异化铁还原诱导次生铁矿对土壤重金属形态转化的影响
1
2018
... Fe(Ⅲ)还原通过异化铁还原菌氧化有机或无机物来进行.异化铁还原菌包含大多数的古菌和细菌,它们在厌氧环境中利用铁还原酶的催化,将电子在呼吸链上传递给末端电子受体Fe(Ⅲ)并产生其所需能量,因此异化铁还原又称铁呼吸[37 ] ,这种途径被认为是最古老的产能方式之一.异化铁还原菌采用3种方式将电子传递到不溶的Fe(Ⅲ)矿物表面:① ② ③ [36 ] . ...
Dissimilatory Fe(III) and Mn(IV) reduction
1
2004
... 从系统发育学的角度看,异化铁还原菌在细菌和古菌中广泛分布,在细菌域主要分布在8个门:栖热袍菌门(Thermotogae)、热脱硫杆菌门(Thermo desulfobacteria)、异常球菌—栖热菌门(Deinococcus-Thermus)、铁还原杆菌门(Deferribacteres)、变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)以及酸杆菌门(Acidobacteria).古菌域主要分布在7个目:热变形菌目(Hermoproteales)、硫还原古球菌目(Desulfurococcales)、硫化叶菌目(Sulfolobales)、甲烷球菌目(Methanococales)、热球菌目(Thermococcales)、古生球菌目(Archaeoglobales)和甲烷嗜高温菌目(Methanopyrales)[38 ] .到目前为止,已有超过71种海洋铁还原菌被鉴定,广泛存在于革兰氏阳性菌和阴性菌中[39 ] . ...
Iron-reducing bacteria and iron nanostructures
1
2017
... 从系统发育学的角度看,异化铁还原菌在细菌和古菌中广泛分布,在细菌域主要分布在8个门:栖热袍菌门(Thermotogae)、热脱硫杆菌门(Thermo desulfobacteria)、异常球菌—栖热菌门(Deinococcus-Thermus)、铁还原杆菌门(Deferribacteres)、变形菌门(Proteobacteria)、厚壁菌门(Firmicutes)、放线菌门(Actinobacteria)以及酸杆菌门(Acidobacteria).古菌域主要分布在7个目:热变形菌目(Hermoproteales)、硫还原古球菌目(Desulfurococcales)、硫化叶菌目(Sulfolobales)、甲烷球菌目(Methanococales)、热球菌目(Thermococcales)、古生球菌目(Archaeoglobales)和甲烷嗜高温菌目(Methanopyrales)[38 ] .到目前为止,已有超过71种海洋铁还原菌被鉴定,广泛存在于革兰氏阳性菌和阴性菌中[39 ] . ...
Microbial Fe(III) reduction in subsurface environments
1
2010
... 地杆菌科(Geobacteraceae)和希瓦氏菌属(Shewanella )是研究最多的2种异化铁还原菌[40 ] .地杆菌属(Geobacter )可以氧化有机物,Fe(Ⅲ)是进行铁的异化还原的唯一电子受体[41 ] .G. sulfurreducens 的胞外电子传递主要通过C-型细胞色素OmcE和OmcS介导,其中OmcS与导电菌毛相连,在外膜孔蛋白OmpJ的帮助下,沿此路径调节电子的传递[36 ] .希瓦氏菌属中S. oneidensis MR-1研究较多,可以利用H+ 、甲酸盐和丙酮酸等进行异化铁还原.MR-1通过C-型细胞色素CymA、质膜C-型细胞色素MtrA、外膜MtrB以及外膜C-型细胞色素MtrC和OmcA来转移电子到Fe(Ⅲ),以此来进行异化铁的还原[42 ] . ...
NanoSIMS imaging of extracellular electron transport processes during microbial iron (III) reduction
1
2018
... 地杆菌科(Geobacteraceae)和希瓦氏菌属(Shewanella )是研究最多的2种异化铁还原菌[40 ] .地杆菌属(Geobacter )可以氧化有机物,Fe(Ⅲ)是进行铁的异化还原的唯一电子受体[41 ] .G. sulfurreducens 的胞外电子传递主要通过C-型细胞色素OmcE和OmcS介导,其中OmcS与导电菌毛相连,在外膜孔蛋白OmpJ的帮助下,沿此路径调节电子的传递[36 ] .希瓦氏菌属中S. oneidensis MR-1研究较多,可以利用H+ 、甲酸盐和丙酮酸等进行异化铁还原.MR-1通过C-型细胞色素CymA、质膜C-型细胞色素MtrA、外膜MtrB以及外膜C-型细胞色素MtrC和OmcA来转移电子到Fe(Ⅲ),以此来进行异化铁的还原[42 ] . ...
The roles of outer membrane cytochromes of Shewanella and Geobacter in extracellular electron transfer
1
2009
... 地杆菌科(Geobacteraceae)和希瓦氏菌属(Shewanella )是研究最多的2种异化铁还原菌[40 ] .地杆菌属(Geobacter )可以氧化有机物,Fe(Ⅲ)是进行铁的异化还原的唯一电子受体[41 ] .G. sulfurreducens 的胞外电子传递主要通过C-型细胞色素OmcE和OmcS介导,其中OmcS与导电菌毛相连,在外膜孔蛋白OmpJ的帮助下,沿此路径调节电子的传递[36 ] .希瓦氏菌属中S. oneidensis MR-1研究较多,可以利用H+ 、甲酸盐和丙酮酸等进行异化铁还原.MR-1通过C-型细胞色素CymA、质膜C-型细胞色素MtrA、外膜MtrB以及外膜C-型细胞色素MtrC和OmcA来转移电子到Fe(Ⅲ),以此来进行异化铁的还原[42 ] . ...
Review on the mechanism and development of ferric iron microbial reduction
2
2006
... 综上所述,我们可以看出Fe(Ⅲ) 还原菌物种多样,生化机制复杂,这与它们在生态系统中的作用息息相关.Fe(Ⅲ) 还原菌既是碳循环的驱动者,也是氮循环的重要参与者.以Geobacter 和Clostridia 为例,前者参与NO3 - [43 ] .探究Fe(Ⅲ)还原菌在生态系统碳和氮循环中的意义将推动我们对海洋铁代谢菌的深入了解.遗憾的是,尽管目前已经从环境中分离得到多种Fe(Ⅲ) 还原菌,但 Fe(Ⅲ)还原菌的全基因组基因信息尚未获得,后续需借助测序平台和现代分子生物学技术拓展对 Fe(Ⅲ)还原菌分子层面的认识[43 ] . ...
... [43 ]. ...
Fe(Ⅲ)微生物还原机理及其研究进展
2
2006
... 综上所述,我们可以看出Fe(Ⅲ) 还原菌物种多样,生化机制复杂,这与它们在生态系统中的作用息息相关.Fe(Ⅲ) 还原菌既是碳循环的驱动者,也是氮循环的重要参与者.以Geobacter 和Clostridia 为例,前者参与NO3 - [43 ] .探究Fe(Ⅲ)还原菌在生态系统碳和氮循环中的意义将推动我们对海洋铁代谢菌的深入了解.遗憾的是,尽管目前已经从环境中分离得到多种Fe(Ⅲ) 还原菌,但 Fe(Ⅲ)还原菌的全基因组基因信息尚未获得,后续需借助测序平台和现代分子生物学技术拓展对 Fe(Ⅲ)还原菌分子层面的认识[43 ] . ...
... [43 ]. ...
Quantification of dissolved iron sources to the North Atlantic Ocean
1
2014
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Elusive marine nitrogen fixation
1
2016
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Local versus basin-scale limitation of marine nitrogen fixation
1
2014
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Ligand-enhanced abiotic iron oxidation and the effects of chemical versus biological iron cycling in anoxic environments
2
2013
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
... [47 ].铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Coupled dynamics of iron and phosphorus in sediments of an oligotrophic coastal basin and the impact of anaerobic oxidation of methane
2
2013
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
... 与磷的关系中也揭示了铁循环与磷元素之间的复杂联系.大西洋的尘埃输入和水团活动控制着固氮生物在磷限制与铁限制之间的转换[54 ] .模型研究显示,由于分层的加强,表层水体中大量营养元素的垂直供应减少,而气溶胶铁的输入增加,铁与养分供应比的增大会影响Fe-P关系,从而对浮游植物生长和多样性产生影响[55 ] .在低纬度海域中,铁来自海洋上方而磷来自下方,因此,这2种营养物质的传递容易受到不同水团的影响[55 ] .随着气溶胶铁输入的增加以及分层强化造成的磷供应的减少,可能会打破Fe/P化学计量比和供应比的平衡,从而大幅降低固氮细菌的增长、氮固定效率以及铁、磷和碳的输出[55 ] .Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
Interactive effects of ocean acidification and nitrogen limitation on two bloom-forming dinoflagellate species
1
2016
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Exploring records of Saharan dust transport and hurricanes in the Caribbean and Gulf of Mexico over recent millennia
1
2017
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Research progress and application status of seawater eutrophication evaluation methods
1
2018
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
海水富营养化评价方法的研究进展与应用现状
1
2018
... 铁作为一种限制性因子,它的生态作用除了自身属性,更重要的是体现在与其他元素的耦合关系上[44 ] .铁与氮的作用最先在赤道大西洋海域研究中报道,该研究发现固氮生物的生产力受氮元素的限制,当铁不作为限制因素时,固氮生物可正常生长[45 ] ;相反,当铁浓度不足时,固氮生物生长受限,这使得低纬度海域中尘埃铁成为维持海洋氮库的重要因素[46 ] .低纬度海区尘埃铁的供应有助于维持海洋的固氮存量,而高纬度地区沉积物和热液铁源的供应推动了铁与氮循环和海面氮气的交换[8 ] .异化铁还原、铁氨氧化、反硝化过程是海洋缺氧环境中Fe-N循环的重要方式.通过异化铁还原、铁氨氧化将环境中的Fe(Ⅲ)还原为Fe(Ⅱ),而Fe(Ⅱ)在反硝化过程中又会作为电子供体被氧化成Fe(Ⅲ),伴随着NH4 + 3 - 4 + 3 - [47 ] .最近研究发现海洋中还存在一类特殊的Fe(Ⅱ)氧化——NO3 - 3 - [47 ] .铁在海洋中的另一种重要用途是帮助浮游植物获取氮,从而导致了Fe-N共限制的产生.有研究表明,由于海洋分层强化、酸化驱动的硝化抑制以及脱氧作用驱动的反硝化作用的增强导致上层水体的硝酸盐浓度下降,使得浮游植物对氮源的依赖性增强[48 ,49 ] .在东墨西哥湾,近岸撒哈拉沙漠灰尘的沉降缓解了该海域铁的限值,使具有固氮能力的海藻(T. erythraeum )在硝酸盐限制的条件下仍可大量繁殖;T. erythraeum 的排泄物中有50% G. breve 利用这些DON大量繁殖,最终导致藻华灾害的发生,影响已有的生态平衡[50 ,51 ] . ...
Why are biotic iron pools uniform across high-and low-iron pelagic ecosystems?
1
2015
... 除了氮,铁与碳元素的关系也被关注,浮游植物的光合作用是固定大气CO2 的主要方式,将无机碳转化为有机碳是食物网碳循环的主要推手,铁的利用与再生为该网络的运行提供了驱动力(图6 )[9 ] .浮游植物、细菌和病毒的铁再生作用在铁的生物供应中扮演着“中转站”的作用[52 ] .有研究发现,再生铁有利于提升浮游植物的固碳效率,铁的吸收对于固碳速率有着根本的影响[24 ] .加利福尼亚海域44个站点设计的铁添加实验,证明了铁限制浮游植物的生长,主要因为钝化了体内有机碳的合成[53 ] . ...
Spatial and temporal variability in phytoplankton iron limitation along the California coast and consequences for Si, N, and C biogeochemistry
1
2003
... 除了氮,铁与碳元素的关系也被关注,浮游植物的光合作用是固定大气CO2 的主要方式,将无机碳转化为有机碳是食物网碳循环的主要推手,铁的利用与再生为该网络的运行提供了驱动力(图6 )[9 ] .浮游植物、细菌和病毒的铁再生作用在铁的生物供应中扮演着“中转站”的作用[52 ] .有研究发现,再生铁有利于提升浮游植物的固碳效率,铁的吸收对于固碳速率有着根本的影响[24 ] .加利福尼亚海域44个站点设计的铁添加实验,证明了铁限制浮游植物的生长,主要因为钝化了体内有机碳的合成[53 ] . ...
Seasonal ITCZ migration dynamically controls the location of the (sub)tropical Atlantic biogeochemical divide
1
2014
... 与磷的关系中也揭示了铁循环与磷元素之间的复杂联系.大西洋的尘埃输入和水团活动控制着固氮生物在磷限制与铁限制之间的转换[54 ] .模型研究显示,由于分层的加强,表层水体中大量营养元素的垂直供应减少,而气溶胶铁的输入增加,铁与养分供应比的增大会影响Fe-P关系,从而对浮游植物生长和多样性产生影响[55 ] .在低纬度海域中,铁来自海洋上方而磷来自下方,因此,这2种营养物质的传递容易受到不同水团的影响[55 ] .随着气溶胶铁输入的增加以及分层强化造成的磷供应的减少,可能会打破Fe/P化学计量比和供应比的平衡,从而大幅降低固氮细菌的增长、氮固定效率以及铁、磷和碳的输出[55 ] .Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
Marine phytoplankton and the changing ocean iron cycle
4
2016
... 与磷的关系中也揭示了铁循环与磷元素之间的复杂联系.大西洋的尘埃输入和水团活动控制着固氮生物在磷限制与铁限制之间的转换[54 ] .模型研究显示,由于分层的加强,表层水体中大量营养元素的垂直供应减少,而气溶胶铁的输入增加,铁与养分供应比的增大会影响Fe-P关系,从而对浮游植物生长和多样性产生影响[55 ] .在低纬度海域中,铁来自海洋上方而磷来自下方,因此,这2种营养物质的传递容易受到不同水团的影响[55 ] .随着气溶胶铁输入的增加以及分层强化造成的磷供应的减少,可能会打破Fe/P化学计量比和供应比的平衡,从而大幅降低固氮细菌的增长、氮固定效率以及铁、磷和碳的输出[55 ] .Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
... [55 ].随着气溶胶铁输入的增加以及分层强化造成的磷供应的减少,可能会打破Fe/P化学计量比和供应比的平衡,从而大幅降低固氮细菌的增长、氮固定效率以及铁、磷和碳的输出[55 ] .Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
... [55 ].Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
... 总结海洋铁循环的运作方式及其与N/C/P循环的关系,我们可以看到低纬度地区中尘埃铁的主导地位与高纬度地区中沉积物和热液铁源的作用不同.高纬度地区的铁供应推动了海洋碳循环,而低纬度地区的尘埃供应有助于维持海洋的固氮存量.微量元素铁的分布相比主导元素更为复杂,其中的驱动因素有深海铁源、颗粒铁沉降、铁的再生与清除、不同水团的影响以及远程运输的配体特性等.综合现有知识,Tagliabue等[8 ] 勾勒了一幅海洋铁循环示意图(图7 ).在高纬度地区,热液和沉积物是铁的主要来源[55 ] .混有铁元素的水流和混合作用是促进生物活性的主要基础,其通过微生物介导的铁再生来维持.高纬度地区的铁通常有限,因此配体以及溶解态络合物可能会潜沉和朝赤道方向输送(图7 ).相反,低纬度地区,大气沉降构成了铁的主要来源,如果尘埃供应量低同时上升流消失,随之而来的铁胁迫会导致固氮率降低[57 ] .低纬度地区的铁受到沉降有机物中再生铁与成岩颗粒铁之间的平衡关系的影响[58 ] .铁的清除率与未络合铁的数量密切相关,低纬度地区的铁可能会受到含有饱和配体的高纬度水的俯冲和赤道流的远程控制[59 ] .这一观点符合太平洋和大西洋的情形,因为两者的经向环流均以高纬度深水团为主.目前,针对铁的精细循环有过多种模型,但不同算法和维度的结果差异甚大,这些差异导致计算出的铁停留时间范围不一致,从几年到几百年不等,这种不确定性干扰了对铁的深度预测.为了减少这种不确定性,需要从新的海洋调查数据中提取更多的铁循环过程和控制因素等信息,以代入更精确的运算模型,求取外部铁供应和内部铁循环的周转时间. ...
Limitation and synergistic effect of iron and phosphorus on the fulminant proliferation of prodinoflagellates and cryptophyta
1
2006
... 与磷的关系中也揭示了铁循环与磷元素之间的复杂联系.大西洋的尘埃输入和水团活动控制着固氮生物在磷限制与铁限制之间的转换[54 ] .模型研究显示,由于分层的加强,表层水体中大量营养元素的垂直供应减少,而气溶胶铁的输入增加,铁与养分供应比的增大会影响Fe-P关系,从而对浮游植物生长和多样性产生影响[55 ] .在低纬度海域中,铁来自海洋上方而磷来自下方,因此,这2种营养物质的传递容易受到不同水团的影响[55 ] .随着气溶胶铁输入的增加以及分层强化造成的磷供应的减少,可能会打破Fe/P化学计量比和供应比的平衡,从而大幅降低固氮细菌的增长、氮固定效率以及铁、磷和碳的输出[55 ] .Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
铁和磷对原甲藻和隐藻暴发性增殖的限制与协同影响
1
2006
... 与磷的关系中也揭示了铁循环与磷元素之间的复杂联系.大西洋的尘埃输入和水团活动控制着固氮生物在磷限制与铁限制之间的转换[54 ] .模型研究显示,由于分层的加强,表层水体中大量营养元素的垂直供应减少,而气溶胶铁的输入增加,铁与养分供应比的增大会影响Fe-P关系,从而对浮游植物生长和多样性产生影响[55 ] .在低纬度海域中,铁来自海洋上方而磷来自下方,因此,这2种营养物质的传递容易受到不同水团的影响[55 ] .随着气溶胶铁输入的增加以及分层强化造成的磷供应的减少,可能会打破Fe/P化学计量比和供应比的平衡,从而大幅降低固氮细菌的增长、氮固定效率以及铁、磷和碳的输出[55 ] .Slomp等[48 ] 研究发现铁结合磷是波罗的海中重要的沉积相,Fe(II)-P矿物是其中重要的磷沉积.室内控制实验中,有学者也从微观尺度研究了铁、磷对原甲藻(Prorocentrummicans Ehrenber )、 隐藻(Cryptomonas sp.)生长的限制作用和协同影响.结果表明,铁缺乏会造成藻细胞叶绿体和线粒体发生扭曲和变形,从而导致藻类生长速度减慢;铁和磷的交互作用对藻类生长具有正态分布性,藻细胞中磷与铁的最佳摩尔比为356∶ 1 [56 ] . ...
Nitrogen fixation in the eastern Atlantic reaches similar levels in the Southern and Northern Hemisphere
1
2017
... 总结海洋铁循环的运作方式及其与N/C/P循环的关系,我们可以看到低纬度地区中尘埃铁的主导地位与高纬度地区中沉积物和热液铁源的作用不同.高纬度地区的铁供应推动了海洋碳循环,而低纬度地区的尘埃供应有助于维持海洋的固氮存量.微量元素铁的分布相比主导元素更为复杂,其中的驱动因素有深海铁源、颗粒铁沉降、铁的再生与清除、不同水团的影响以及远程运输的配体特性等.综合现有知识,Tagliabue等[8 ] 勾勒了一幅海洋铁循环示意图(图7 ).在高纬度地区,热液和沉积物是铁的主要来源[55 ] .混有铁元素的水流和混合作用是促进生物活性的主要基础,其通过微生物介导的铁再生来维持.高纬度地区的铁通常有限,因此配体以及溶解态络合物可能会潜沉和朝赤道方向输送(图7 ).相反,低纬度地区,大气沉降构成了铁的主要来源,如果尘埃供应量低同时上升流消失,随之而来的铁胁迫会导致固氮率降低[57 ] .低纬度地区的铁受到沉降有机物中再生铁与成岩颗粒铁之间的平衡关系的影响[58 ] .铁的清除率与未络合铁的数量密切相关,低纬度地区的铁可能会受到含有饱和配体的高纬度水的俯冲和赤道流的远程控制[59 ] .这一观点符合太平洋和大西洋的情形,因为两者的经向环流均以高纬度深水团为主.目前,针对铁的精细循环有过多种模型,但不同算法和维度的结果差异甚大,这些差异导致计算出的铁停留时间范围不一致,从几年到几百年不等,这种不确定性干扰了对铁的深度预测.为了减少这种不确定性,需要从新的海洋调查数据中提取更多的铁循环过程和控制因素等信息,以代入更精确的运算模型,求取外部铁供应和内部铁循环的周转时间. ...
The biogeochemical cycle of iron in the ocean
1
2010
... 总结海洋铁循环的运作方式及其与N/C/P循环的关系,我们可以看到低纬度地区中尘埃铁的主导地位与高纬度地区中沉积物和热液铁源的作用不同.高纬度地区的铁供应推动了海洋碳循环,而低纬度地区的尘埃供应有助于维持海洋的固氮存量.微量元素铁的分布相比主导元素更为复杂,其中的驱动因素有深海铁源、颗粒铁沉降、铁的再生与清除、不同水团的影响以及远程运输的配体特性等.综合现有知识,Tagliabue等[8 ] 勾勒了一幅海洋铁循环示意图(图7 ).在高纬度地区,热液和沉积物是铁的主要来源[55 ] .混有铁元素的水流和混合作用是促进生物活性的主要基础,其通过微生物介导的铁再生来维持.高纬度地区的铁通常有限,因此配体以及溶解态络合物可能会潜沉和朝赤道方向输送(图7 ).相反,低纬度地区,大气沉降构成了铁的主要来源,如果尘埃供应量低同时上升流消失,随之而来的铁胁迫会导致固氮率降低[57 ] .低纬度地区的铁受到沉降有机物中再生铁与成岩颗粒铁之间的平衡关系的影响[58 ] .铁的清除率与未络合铁的数量密切相关,低纬度地区的铁可能会受到含有饱和配体的高纬度水的俯冲和赤道流的远程控制[59 ] .这一观点符合太平洋和大西洋的情形,因为两者的经向环流均以高纬度深水团为主.目前,针对铁的精细循环有过多种模型,但不同算法和维度的结果差异甚大,这些差异导致计算出的铁停留时间范围不一致,从几年到几百年不等,这种不确定性干扰了对铁的深度预测.为了减少这种不确定性,需要从新的海洋调查数据中提取更多的铁循环过程和控制因素等信息,以代入更精确的运算模型,求取外部铁供应和内部铁循环的周转时间. ...
A ventilation- based framework to explain the regeneration-scavenging balance of iron in the ocean
1
2015
... 总结海洋铁循环的运作方式及其与N/C/P循环的关系,我们可以看到低纬度地区中尘埃铁的主导地位与高纬度地区中沉积物和热液铁源的作用不同.高纬度地区的铁供应推动了海洋碳循环,而低纬度地区的尘埃供应有助于维持海洋的固氮存量.微量元素铁的分布相比主导元素更为复杂,其中的驱动因素有深海铁源、颗粒铁沉降、铁的再生与清除、不同水团的影响以及远程运输的配体特性等.综合现有知识,Tagliabue等[8 ] 勾勒了一幅海洋铁循环示意图(图7 ).在高纬度地区,热液和沉积物是铁的主要来源[55 ] .混有铁元素的水流和混合作用是促进生物活性的主要基础,其通过微生物介导的铁再生来维持.高纬度地区的铁通常有限,因此配体以及溶解态络合物可能会潜沉和朝赤道方向输送(图7 ).相反,低纬度地区,大气沉降构成了铁的主要来源,如果尘埃供应量低同时上升流消失,随之而来的铁胁迫会导致固氮率降低[57 ] .低纬度地区的铁受到沉降有机物中再生铁与成岩颗粒铁之间的平衡关系的影响[58 ] .铁的清除率与未络合铁的数量密切相关,低纬度地区的铁可能会受到含有饱和配体的高纬度水的俯冲和赤道流的远程控制[59 ] .这一观点符合太平洋和大西洋的情形,因为两者的经向环流均以高纬度深水团为主.目前,针对铁的精细循环有过多种模型,但不同算法和维度的结果差异甚大,这些差异导致计算出的铁停留时间范围不一致,从几年到几百年不等,这种不确定性干扰了对铁的深度预测.为了减少这种不确定性,需要从新的海洋调查数据中提取更多的铁循环过程和控制因素等信息,以代入更精确的运算模型,求取外部铁供应和内部铁循环的周转时间. ...
Objective estimates of mantle 3 He in the ocean and implications for constraining the deep ocean circulation
1
2017
... (1)生物监测与物理观测的耦合,加强以稳定同位素为代表的示踪量化.以往采用了3 He来识别大洋中脊的铁羽流[60 ] ,并推导出铁和氦的热液通量关系.此外,在一些“GEOTRACES”区段上测定了镭的衰变[61 ] ,并且可将其与溶解铁数据集相关联,估算出了铁边界数值.未来的研究中,稳定同位素法结合海洋涡旋的测定是一个重要方向,可以用于确定铁的浓度在传输间的动态变化,还可以有效规避远程信号与交错过程的信号混杂[8 ] .在这种情形下铁的分布特征具有“指纹化”,可用来追踪铁的循环路径[8 ] .此外,结合物理场景(水团、密度或涡旋)的考虑,还有助于定量关键过程的速率,并建立铁与主要营养素之间的偶联或去偶联关系,有利于全球模型参数化的统一. ...
The Ra-226-Ba relationship in the North Atlantic during GEOTRACES-GA01
1
2018
... (1)生物监测与物理观测的耦合,加强以稳定同位素为代表的示踪量化.以往采用了3 He来识别大洋中脊的铁羽流[60 ] ,并推导出铁和氦的热液通量关系.此外,在一些“GEOTRACES”区段上测定了镭的衰变[61 ] ,并且可将其与溶解铁数据集相关联,估算出了铁边界数值.未来的研究中,稳定同位素法结合海洋涡旋的测定是一个重要方向,可以用于确定铁的浓度在传输间的动态变化,还可以有效规避远程信号与交错过程的信号混杂[8 ] .在这种情形下铁的分布特征具有“指纹化”,可用来追踪铁的循环路径[8 ] .此外,结合物理场景(水团、密度或涡旋)的考虑,还有助于定量关键过程的速率,并建立铁与主要营养素之间的偶联或去偶联关系,有利于全球模型参数化的统一. ...






 甘公网安备62010202000687
甘公网安备62010202000687{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}