Basic Research Opportunities in Earth Science
3
2001
... 地球关键带(Critical Zone, CZ)是指地球浅层岩石—土壤—大气—水—生物及人类活动相互作用,为生命系统提供支撑资源的异质的地球表层系统[1,2,3,4].土壤及其生态功能被认为是关键带的核心,但在关键带理论框架下土壤涵盖的内容更广,关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5]. ...
... 地球关键带研究是将单独研究地球表层系统的科学(地质学、生态学、水文学、土壤学、地球化学和地貌学等)在观测和模拟过程中进行有机整合的综合性科学[1,2,3,4].美国国家研究理事会在2001年出版的《地球基础科学研究的机遇》中首次正式提出地球关键带的概念[1].地球科学的“关键带”和生态学的“生态系统”概念上具有显著的一致性,其差异主要体现在垂直尺度上[5].生态系统地上部分的生命体显著影响整个岩石风化层—土壤剖面的生物地球化学过程,但大部分陆地生态系统的下边界划定通常限于1 m深度,不能完整地反映整个生态系统的结构和功能[5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... [1].地球科学的“关键带”和生态学的“生态系统”概念上具有显著的一致性,其差异主要体现在垂直尺度上[5].生态系统地上部分的生命体显著影响整个岩石风化层—土壤剖面的生物地球化学过程,但大部分陆地生态系统的下边界划定通常限于1 m深度,不能完整地反映整个生态系统的结构和功能[5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
Frontiers in Exploration of the Critical Zone
4
2006
... 地球关键带(Critical Zone, CZ)是指地球浅层岩石—土壤—大气—水—生物及人类活动相互作用,为生命系统提供支撑资源的异质的地球表层系统[1,2,3,4].土壤及其生态功能被认为是关键带的核心,但在关键带理论框架下土壤涵盖的内容更广,关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5]. ...
... 地球关键带研究是将单独研究地球表层系统的科学(地质学、生态学、水文学、土壤学、地球化学和地貌学等)在观测和模拟过程中进行有机整合的综合性科学[1,2,3,4].美国国家研究理事会在2001年出版的《地球基础科学研究的机遇》中首次正式提出地球关键带的概念[1].地球科学的“关键带”和生态学的“生态系统”概念上具有显著的一致性,其差异主要体现在垂直尺度上[5].生态系统地上部分的生命体显著影响整个岩石风化层—土壤剖面的生物地球化学过程,但大部分陆地生态系统的下边界划定通常限于1 m深度,不能完整地反映整个生态系统的结构和功能[5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... [2,3,4,5]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
Sustaining Earth's Critical Zone Basic Science and Interdisciplinary Solutions for Global Challenges
4
2013
... 地球关键带(Critical Zone, CZ)是指地球浅层岩石—土壤—大气—水—生物及人类活动相互作用,为生命系统提供支撑资源的异质的地球表层系统[1,2,3,4].土壤及其生态功能被认为是关键带的核心,但在关键带理论框架下土壤涵盖的内容更广,关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5]. ...
... 地球关键带研究是将单独研究地球表层系统的科学(地质学、生态学、水文学、土壤学、地球化学和地貌学等)在观测和模拟过程中进行有机整合的综合性科学[1,2,3,4].美国国家研究理事会在2001年出版的《地球基础科学研究的机遇》中首次正式提出地球关键带的概念[1].地球科学的“关键带”和生态学的“生态系统”概念上具有显著的一致性,其差异主要体现在垂直尺度上[5].生态系统地上部分的生命体显著影响整个岩石风化层—土壤剖面的生物地球化学过程,但大部分陆地生态系统的下边界划定通常限于1 m深度,不能完整地反映整个生态系统的结构和功能[5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... ,3,4,5]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
New Opportunities for Critical Zone Science
5
2017
... 地球关键带(Critical Zone, CZ)是指地球浅层岩石—土壤—大气—水—生物及人类活动相互作用,为生命系统提供支撑资源的异质的地球表层系统[1,2,3,4].土壤及其生态功能被认为是关键带的核心,但在关键带理论框架下土壤涵盖的内容更广,关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5]. ...
... 地球关键带研究是将单独研究地球表层系统的科学(地质学、生态学、水文学、土壤学、地球化学和地貌学等)在观测和模拟过程中进行有机整合的综合性科学[1,2,3,4].美国国家研究理事会在2001年出版的《地球基础科学研究的机遇》中首次正式提出地球关键带的概念[1].地球科学的“关键带”和生态学的“生态系统”概念上具有显著的一致性,其差异主要体现在垂直尺度上[5].生态系统地上部分的生命体显著影响整个岩石风化层—土壤剖面的生物地球化学过程,但大部分陆地生态系统的下边界划定通常限于1 m深度,不能完整地反映整个生态系统的结构和功能[5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... ,4,5]. ...
... 流域尺度上从冠层到基岩生态系统各碳库同位素(δ13C)特征以及各库之间碳流动产生的同位素分馏效应,可以辅助解析植物碳固定及分配、从地表到基岩的土壤碳库分解和转化及小流域碳迁移与平衡等生物地球化学过程与机制[4,11].稳定同位素光谱(Isotope Ratio Infrared Spectroscopy,IRIS)和质谱(Isotope Ratio Mass Spectrometry,IRMS)技术的进步使气体、固体及溶液中各种形态碳同位素的测定成为可能[11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
One physical system: Tansley's ecosystem as Earth's critical zone, Tansley review
5
2015
... 地球关键带(Critical Zone, CZ)是指地球浅层岩石—土壤—大气—水—生物及人类活动相互作用,为生命系统提供支撑资源的异质的地球表层系统[1,2,3,4].土壤及其生态功能被认为是关键带的核心,但在关键带理论框架下土壤涵盖的内容更广,关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5]. ...
... 地球关键带研究是将单独研究地球表层系统的科学(地质学、生态学、水文学、土壤学、地球化学和地貌学等)在观测和模拟过程中进行有机整合的综合性科学[1,2,3,4].美国国家研究理事会在2001年出版的《地球基础科学研究的机遇》中首次正式提出地球关键带的概念[1].地球科学的“关键带”和生态学的“生态系统”概念上具有显著的一致性,其差异主要体现在垂直尺度上[5].生态系统地上部分的生命体显著影响整个岩石风化层—土壤剖面的生物地球化学过程,但大部分陆地生态系统的下边界划定通常限于1 m深度,不能完整地反映整个生态系统的结构和功能[5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... [5].关键带科学本质上是从冠层到基岩重新认识整个生态系统的结构和功能[5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... [5],在流域尺度上强调大气和植物之间、植物和土壤之间、小流域土壤和溪流之间物质和元素循环的相互联系等[2,3,4,5]. ...
... ,5]. ...
In situ (CO2)-C-13 pulse labelling of field-grown eucalypt trees revealed the effects of potassium nutrition and throughfall exclusion on phloem transport of photosynthetic carbon
1
2016
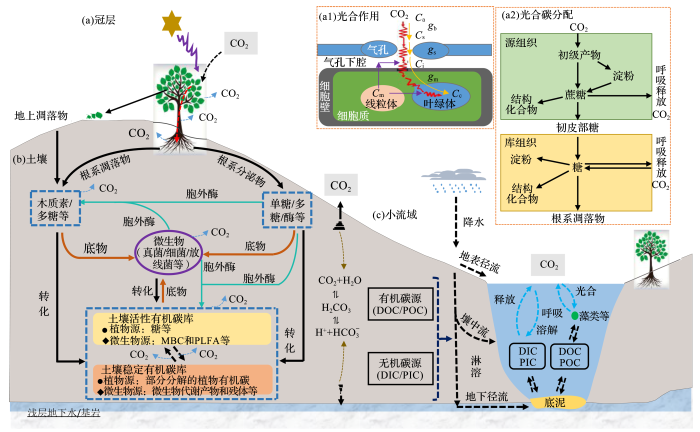
... 碳循环是地球系统物质和能量循环的核心,是地圈—生物圈—大气圈相互作用的纽带.从地球关键带视角理解生态系统碳循环的生物地球化学过程与机制,应该强调植物碳固定及分配[6]、从地表到基岩的土壤碳库分解和转化[7]以及小流域碳迁移与平衡[8]等关键环节(图1).例如,基岩风化氮输入是生态系统被忽视的重要氮源,可以增加森林生产力,进而增加生态系统碳储量[9];土壤物理侵蚀速率决定了海洋沉积物的碳沉积速率,远高于硅酸盐风化的碳吸收量[10]. ...
Persistence of soil organic matter as an ecosystem property
3
2011
... 碳循环是地球系统物质和能量循环的核心,是地圈—生物圈—大气圈相互作用的纽带.从地球关键带视角理解生态系统碳循环的生物地球化学过程与机制,应该强调植物碳固定及分配[6]、从地表到基岩的土壤碳库分解和转化[7]以及小流域碳迁移与平衡[8]等关键环节(图1).例如,基岩风化氮输入是生态系统被忽视的重要氮源,可以增加森林生产力,进而增加生态系统碳储量[9];土壤物理侵蚀速率决定了海洋沉积物的碳沉积速率,远高于硅酸盐风化的碳吸收量[10]. ...
... 从地表到基岩的土壤微生物及胞外酶驱动的土壤有机碳库的分解与转化是碳生物地球化学循环的周转环节(图1b).植物通过地上凋落物、根系凋落物、根系分泌物向土壤中输入有机质,通过胞外酶的矿化和微生物同化,形成土壤有机质碳库[7,34,35].土壤微生物种群结构和酶活性的改变将会改变土壤碳库的分解及蓄积[36]. ...
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
Lateral transport of soil carbon and land-atmosphere CO2 flux induced by water erosion in China
2
2016
... 碳循环是地球系统物质和能量循环的核心,是地圈—生物圈—大气圈相互作用的纽带.从地球关键带视角理解生态系统碳循环的生物地球化学过程与机制,应该强调植物碳固定及分配[6]、从地表到基岩的土壤碳库分解和转化[7]以及小流域碳迁移与平衡[8]等关键环节(图1).例如,基岩风化氮输入是生态系统被忽视的重要氮源,可以增加森林生产力,进而增加生态系统碳储量[9];土壤物理侵蚀速率决定了海洋沉积物的碳沉积速率,远高于硅酸盐风化的碳吸收量[10]. ...
... 土壤碳损失除了以CO2气态形式直接排放以外,主要以地表径流和碳淋溶(壤中流和地下径流)的途径流失[8,66].土壤碳损失主要受CO2通量[250~1 220 g C/(m2·a)]影响,DOC淋溶引起的碳损失较少[7.0~9.4 g C/(m2·a)][66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
Increased forest ecosystem carbon and nitrogen storage from nitrogen rich bedrock
3
2011
... 碳循环是地球系统物质和能量循环的核心,是地圈—生物圈—大气圈相互作用的纽带.从地球关键带视角理解生态系统碳循环的生物地球化学过程与机制,应该强调植物碳固定及分配[6]、从地表到基岩的土壤碳库分解和转化[7]以及小流域碳迁移与平衡[8]等关键环节(图1).例如,基岩风化氮输入是生态系统被忽视的重要氮源,可以增加森林生产力,进而增加生态系统碳储量[9];土壤物理侵蚀速率决定了海洋沉积物的碳沉积速率,远高于硅酸盐风化的碳吸收量[10]. ...
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Global carbon export from the terrestrial biosphere controlled by erosion
1
2015
... 碳循环是地球系统物质和能量循环的核心,是地圈—生物圈—大气圈相互作用的纽带.从地球关键带视角理解生态系统碳循环的生物地球化学过程与机制,应该强调植物碳固定及分配[6]、从地表到基岩的土壤碳库分解和转化[7]以及小流域碳迁移与平衡[8]等关键环节(图1).例如,基岩风化氮输入是生态系统被忽视的重要氮源,可以增加森林生产力,进而增加生态系统碳储量[9];土壤物理侵蚀速率决定了海洋沉积物的碳沉积速率,远高于硅酸盐风化的碳吸收量[10]. ...
Stable Isotope Ecology
3
2013
... 流域尺度上从冠层到基岩生态系统各碳库同位素(δ13C)特征以及各库之间碳流动产生的同位素分馏效应,可以辅助解析植物碳固定及分配、从地表到基岩的土壤碳库分解和转化及小流域碳迁移与平衡等生物地球化学过程与机制[4,11].稳定同位素光谱(Isotope Ratio Infrared Spectroscopy,IRIS)和质谱(Isotope Ratio Mass Spectrometry,IRMS)技术的进步使气体、固体及溶液中各种形态碳同位素的测定成为可能[11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
... [11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
... 土壤有机质分解所释放的CO2 δ13C包含了土壤有机碳、微生物碳和微生物呼吸CO2等来源的δ13C信号[13].森林生态系统中植物生育期内根际碳释放进入土壤有机质库的数量大于地上凋落物[60],这部分根际碳将携带特殊的δ13C特征(13C贫化)进入土壤有机质库,若被微生物快速分解则将影响δ13C.土壤中木质素的δ13C相对于纤维素贫化6,土壤有机质δ13C贫化和富集的程度,依赖于纤维素和木质素之间最初的δ13C差异以及影响纤维素和木质素分解速率的因子[11].通过IRIS和箱式法技术,结合简单二元混合模型,可将土壤呼吸区分为自养呼吸和异养呼吸两部分[61];利用三元混合模型,将地中海暖温带森林地表排放CO2的源拆分为3个来源,即根、凋落物和土壤碳[62]. ...
稳定同位素生态学
3
2013
... 流域尺度上从冠层到基岩生态系统各碳库同位素(δ13C)特征以及各库之间碳流动产生的同位素分馏效应,可以辅助解析植物碳固定及分配、从地表到基岩的土壤碳库分解和转化及小流域碳迁移与平衡等生物地球化学过程与机制[4,11].稳定同位素光谱(Isotope Ratio Infrared Spectroscopy,IRIS)和质谱(Isotope Ratio Mass Spectrometry,IRMS)技术的进步使气体、固体及溶液中各种形态碳同位素的测定成为可能[11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
... [11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
... 土壤有机质分解所释放的CO2 δ13C包含了土壤有机碳、微生物碳和微生物呼吸CO2等来源的δ13C信号[13].森林生态系统中植物生育期内根际碳释放进入土壤有机质库的数量大于地上凋落物[60],这部分根际碳将携带特殊的δ13C特征(13C贫化)进入土壤有机质库,若被微生物快速分解则将影响δ13C.土壤中木质素的δ13C相对于纤维素贫化6,土壤有机质δ13C贫化和富集的程度,依赖于纤维素和木质素之间最初的δ13C差异以及影响纤维素和木质素分解速率的因子[11].通过IRIS和箱式法技术,结合简单二元混合模型,可将土壤呼吸区分为自养呼吸和异养呼吸两部分[61];利用三元混合模型,将地中海暖温带森林地表排放CO2的源拆分为3个来源,即根、凋落物和土壤碳[62]. ...
Evaluating calibration strategies for isotope ratio infrared spectroscopy for atmospheric 13CO2/12CO2 measurement
1
2013
... 流域尺度上从冠层到基岩生态系统各碳库同位素(δ13C)特征以及各库之间碳流动产生的同位素分馏效应,可以辅助解析植物碳固定及分配、从地表到基岩的土壤碳库分解和转化及小流域碳迁移与平衡等生物地球化学过程与机制[4,11].稳定同位素光谱(Isotope Ratio Infrared Spectroscopy,IRIS)和质谱(Isotope Ratio Mass Spectrometry,IRMS)技术的进步使气体、固体及溶液中各种形态碳同位素的测定成为可能[11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
Diel variations in the carbon isotope composition of respired CO2 and associated carbon sources: A review of dynamics and mechanisms
3
2011
... 流域尺度上从冠层到基岩生态系统各碳库同位素(δ13C)特征以及各库之间碳流动产生的同位素分馏效应,可以辅助解析植物碳固定及分配、从地表到基岩的土壤碳库分解和转化及小流域碳迁移与平衡等生物地球化学过程与机制[4,11].稳定同位素光谱(Isotope Ratio Infrared Spectroscopy,IRIS)和质谱(Isotope Ratio Mass Spectrometry,IRMS)技术的进步使气体、固体及溶液中各种形态碳同位素的测定成为可能[11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
... 土壤有机质分解所释放的CO2 δ13C包含了土壤有机碳、微生物碳和微生物呼吸CO2等来源的δ13C信号[13].森林生态系统中植物生育期内根际碳释放进入土壤有机质库的数量大于地上凋落物[60],这部分根际碳将携带特殊的δ13C特征(13C贫化)进入土壤有机质库,若被微生物快速分解则将影响δ13C.土壤中木质素的δ13C相对于纤维素贫化6,土壤有机质δ13C贫化和富集的程度,依赖于纤维素和木质素之间最初的δ13C差异以及影响纤维素和木质素分解速率的因子[11].通过IRIS和箱式法技术,结合简单二元混合模型,可将土壤呼吸区分为自养呼吸和异养呼吸两部分[61];利用三元混合模型,将地中海暖温带森林地表排放CO2的源拆分为3个来源,即根、凋落物和土壤碳[62]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Differentiated correction on the signal intensity dependence of GasBench Ⅱ-IRMS from blank effect and instrument nonlinear effect
1
2017
... 流域尺度上从冠层到基岩生态系统各碳库同位素(δ13C)特征以及各库之间碳流动产生的同位素分馏效应,可以辅助解析植物碳固定及分配、从地表到基岩的土壤碳库分解和转化及小流域碳迁移与平衡等生物地球化学过程与机制[4,11].稳定同位素光谱(Isotope Ratio Infrared Spectroscopy,IRIS)和质谱(Isotope Ratio Mass Spectrometry,IRMS)技术的进步使气体、固体及溶液中各种形态碳同位素的测定成为可能[11,12,13,14].与同位素技术结合,通过野外原位长期连续观测和采样、野外或室内控制实验,可以解析在小时尺度、日尺度、季节尺度和年尺度上从冠层到基岩生态系统碳循环过程及其控制机制. ...
Seasonal variations drive short-term dynamics and partitioning of recently assimilated carbon in the foliage of adult beech and pine
1
2017
... 植物光合CO2供应和需求以及光合碳分配过程是碳生物地球化学循环的起始环节(图1a).光合作用是陆地生态系统碳循环的起始阶段,其形成的碳水化合物用于维持植物呼吸、生长和储存等[15]. ...
Extremely thick cell walls and low mesophyll conductance: Welcome to the world of ancient living
1
2017
... 群落中不同植物具有不同的叶片形态、解剖结构以及养分特征,维持相对较高的光合速率,以维持植物存活、生长以及群落结构.叶片气孔导度(gs)、叶肉导度(gm)和最大羧化速率(Vcmax)共同影响植物光合速率(A),且各因子对A的相对贡献率受树种及样地条件的影响[16].以往研究假设gm是无限的,认为A仅受gs和Vcmax的影响[17].然而,gm是有限的,且种间差异很大,这种差异与叶片解剖结构相关[18].将IRIS技术与传统叶片CO2响应测定技术结合可以获取A,gs,gm和Vcmax等参数,量化植物光合CO2供应和需求关系[19]. ...
The role of mesophyll conductance in the economics of nitrogen and water use in photosynthesis
1
2014
... 群落中不同植物具有不同的叶片形态、解剖结构以及养分特征,维持相对较高的光合速率,以维持植物存活、生长以及群落结构.叶片气孔导度(gs)、叶肉导度(gm)和最大羧化速率(Vcmax)共同影响植物光合速率(A),且各因子对A的相对贡献率受树种及样地条件的影响[16].以往研究假设gm是无限的,认为A仅受gs和Vcmax的影响[17].然而,gm是有限的,且种间差异很大,这种差异与叶片解剖结构相关[18].将IRIS技术与传统叶片CO2响应测定技术结合可以获取A,gs,gm和Vcmax等参数,量化植物光合CO2供应和需求关系[19]. ...
Mesophyll diffusion conductance to CO2: An unappreciated central player in photosynthesis
1
2012
... 群落中不同植物具有不同的叶片形态、解剖结构以及养分特征,维持相对较高的光合速率,以维持植物存活、生长以及群落结构.叶片气孔导度(gs)、叶肉导度(gm)和最大羧化速率(Vcmax)共同影响植物光合速率(A),且各因子对A的相对贡献率受树种及样地条件的影响[16].以往研究假设gm是无限的,认为A仅受gs和Vcmax的影响[17].然而,gm是有限的,且种间差异很大,这种差异与叶片解剖结构相关[18].将IRIS技术与传统叶片CO2响应测定技术结合可以获取A,gs,gm和Vcmax等参数,量化植物光合CO2供应和需求关系[19]. ...
Photosynthetic carbon isotope discrimination and its relationship to the carbon isotope signals of stem, soil and ecosystem respiration
1
2010
... 群落中不同植物具有不同的叶片形态、解剖结构以及养分特征,维持相对较高的光合速率,以维持植物存活、生长以及群落结构.叶片气孔导度(gs)、叶肉导度(gm)和最大羧化速率(Vcmax)共同影响植物光合速率(A),且各因子对A的相对贡献率受树种及样地条件的影响[16].以往研究假设gm是无限的,认为A仅受gs和Vcmax的影响[17].然而,gm是有限的,且种间差异很大,这种差异与叶片解剖结构相关[18].将IRIS技术与传统叶片CO2响应测定技术结合可以获取A,gs,gm和Vcmax等参数,量化植物光合CO2供应和需求关系[19]. ...
Photosynthetic limitations in two Antarctic vascular plants: Importance of leaf anatomical traits and Rubisco kinetic parameters
1
2017
... 植物通过权衡叶片光合过程中CO2供应(gs和gm)和需求(Vcmax)的关系,维持相对较高的光合速率[20].CO2供应和需求权衡关系对植物适应环境、充分利用资源以及分化生态位非常重要.gs和gm对A的限制作用受叶片及叶肉组织结构等因素的影响[21,22].Vcmax对A的影响不仅受叶片内部CO2浓度梯度的影响,还受叶片养分条件的影响[21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
Cell-level anatomical characteristics explain high mesophyll conductance and photosynthetic capacity in sclerophyllous Mediterranean oaks
2
2017
... 植物通过权衡叶片光合过程中CO2供应(gs和gm)和需求(Vcmax)的关系,维持相对较高的光合速率[20].CO2供应和需求权衡关系对植物适应环境、充分利用资源以及分化生态位非常重要.gs和gm对A的限制作用受叶片及叶肉组织结构等因素的影响[21,22].Vcmax对A的影响不仅受叶片内部CO2浓度梯度的影响,还受叶片养分条件的影响[21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
... [21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
The strategies of water-carbon regulation of plants in a subtropical primary forest on karst soils in China
1
2018
... 植物通过权衡叶片光合过程中CO2供应(gs和gm)和需求(Vcmax)的关系,维持相对较高的光合速率[20].CO2供应和需求权衡关系对植物适应环境、充分利用资源以及分化生态位非常重要.gs和gm对A的限制作用受叶片及叶肉组织结构等因素的影响[21,22].Vcmax对A的影响不仅受叶片内部CO2浓度梯度的影响,还受叶片养分条件的影响[21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
More nitrogen partition in structural proteins and decreased photosynthetic nitrogen-use efficiency of Pinus massoniana under in situ polluted stress
1
2011
... 植物通过权衡叶片光合过程中CO2供应(gs和gm)和需求(Vcmax)的关系,维持相对较高的光合速率[20].CO2供应和需求权衡关系对植物适应环境、充分利用资源以及分化生态位非常重要.gs和gm对A的限制作用受叶片及叶肉组织结构等因素的影响[21,22].Vcmax对A的影响不仅受叶片内部CO2浓度梯度的影响,还受叶片养分条件的影响[21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
Soil parent material—A major driver of plant nutrient limitations in terrestrial ecosystems
1
2017
... 植物通过权衡叶片光合过程中CO2供应(gs和gm)和需求(Vcmax)的关系,维持相对较高的光合速率[20].CO2供应和需求权衡关系对植物适应环境、充分利用资源以及分化生态位非常重要.gs和gm对A的限制作用受叶片及叶肉组织结构等因素的影响[21,22].Vcmax对A的影响不仅受叶片内部CO2浓度梯度的影响,还受叶片养分条件的影响[21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
Evolution of Photosynthesis
1
2011
... 植物通过权衡叶片光合过程中CO2供应(gs和gm)和需求(Vcmax)的关系,维持相对较高的光合速率[20].CO2供应和需求权衡关系对植物适应环境、充分利用资源以及分化生态位非常重要.gs和gm对A的限制作用受叶片及叶肉组织结构等因素的影响[21,22].Vcmax对A的影响不仅受叶片内部CO2浓度梯度的影响,还受叶片养分条件的影响[21].植物通过不同的叶片经济策略来适应土壤的养分条件[23].不同地区A受氮(N)或磷(P)限制,或N-P协同限制,其中N-P协同限制占主导地位[24].除了叶片N和P元素,叶片矿物元素(钙、镁、钾、钠等)对A也存在调控作用[25]. ...
Carbohydrate storage enhances seedling shade and stress tolerance in a neotropical forest
2
2007
... 了解不同物种植物在不同时期非结构性碳水化合物(Non-Structural Carbohydrate,NSC)和结构性碳水化合物(Structural Carbohydrate,SC)动态及权衡关系对植物体内碳平衡的影响,是揭示植物生长和维持机制的关键[26].植物储存的NSC可以用于植物春季萌发生长、冬季植物维持呼吸、缓解水分胁迫等[27].多年生植物NSC的储存特性可以反映植物自身生长及其对环境的适应策略[28].由于常绿树种储存了更多的NSC维持植物体内的碳平衡,通常常绿树种NSC浓度差异较落叶树种NSC浓度差异小[29].落叶树种在早春叶片还没有萌芽阶段,从永久性结构组织中活化NSC供应植物生长和呼吸,从而导致储存器官中NSC含量降低[30].NSC季节变异可以表征植物对水分、光照、温度和养分条件的适应策略[26].在干旱胁迫条件下植物会利用储存的NSC满足植物维持代谢需要、水势适应、组织修复以及应对病原体灾害[28]. ...
... [26].在干旱胁迫条件下植物会利用储存的NSC满足植物维持代谢需要、水势适应、组织修复以及应对病原体灾害[28]. ...
Non-structural carbohydrate pools in a tropical forest
1
2005
... 了解不同物种植物在不同时期非结构性碳水化合物(Non-Structural Carbohydrate,NSC)和结构性碳水化合物(Structural Carbohydrate,SC)动态及权衡关系对植物体内碳平衡的影响,是揭示植物生长和维持机制的关键[26].植物储存的NSC可以用于植物春季萌发生长、冬季植物维持呼吸、缓解水分胁迫等[27].多年生植物NSC的储存特性可以反映植物自身生长及其对环境的适应策略[28].由于常绿树种储存了更多的NSC维持植物体内的碳平衡,通常常绿树种NSC浓度差异较落叶树种NSC浓度差异小[29].落叶树种在早春叶片还没有萌芽阶段,从永久性结构组织中活化NSC供应植物生长和呼吸,从而导致储存器官中NSC含量降低[30].NSC季节变异可以表征植物对水分、光照、温度和养分条件的适应策略[26].在干旱胁迫条件下植物会利用储存的NSC满足植物维持代谢需要、水势适应、组织修复以及应对病原体灾害[28]. ...
The fate of recently fixed carbon after drought release: Towards unravelling C storage regulation in Tilia platyphyllos and Pinus sylvestris
6
2017
... 了解不同物种植物在不同时期非结构性碳水化合物(Non-Structural Carbohydrate,NSC)和结构性碳水化合物(Structural Carbohydrate,SC)动态及权衡关系对植物体内碳平衡的影响,是揭示植物生长和维持机制的关键[26].植物储存的NSC可以用于植物春季萌发生长、冬季植物维持呼吸、缓解水分胁迫等[27].多年生植物NSC的储存特性可以反映植物自身生长及其对环境的适应策略[28].由于常绿树种储存了更多的NSC维持植物体内的碳平衡,通常常绿树种NSC浓度差异较落叶树种NSC浓度差异小[29].落叶树种在早春叶片还没有萌芽阶段,从永久性结构组织中活化NSC供应植物生长和呼吸,从而导致储存器官中NSC含量降低[30].NSC季节变异可以表征植物对水分、光照、温度和养分条件的适应策略[26].在干旱胁迫条件下植物会利用储存的NSC满足植物维持代谢需要、水势适应、组织修复以及应对病原体灾害[28]. ...
... [28]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
... ,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
... [28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Non-structural carbon compounds in temperate forest trees
1
2003
... 了解不同物种植物在不同时期非结构性碳水化合物(Non-Structural Carbohydrate,NSC)和结构性碳水化合物(Structural Carbohydrate,SC)动态及权衡关系对植物体内碳平衡的影响,是揭示植物生长和维持机制的关键[26].植物储存的NSC可以用于植物春季萌发生长、冬季植物维持呼吸、缓解水分胁迫等[27].多年生植物NSC的储存特性可以反映植物自身生长及其对环境的适应策略[28].由于常绿树种储存了更多的NSC维持植物体内的碳平衡,通常常绿树种NSC浓度差异较落叶树种NSC浓度差异小[29].落叶树种在早春叶片还没有萌芽阶段,从永久性结构组织中活化NSC供应植物生长和呼吸,从而导致储存器官中NSC含量降低[30].NSC季节变异可以表征植物对水分、光照、温度和养分条件的适应策略[26].在干旱胁迫条件下植物会利用储存的NSC满足植物维持代谢需要、水势适应、组织修复以及应对病原体灾害[28]. ...
Non-structural carbohydrates and nitrogen dynamics in mediterranean sub-shrubs: An analysis of the functional role of overwintering leaves
1
2007
... 了解不同物种植物在不同时期非结构性碳水化合物(Non-Structural Carbohydrate,NSC)和结构性碳水化合物(Structural Carbohydrate,SC)动态及权衡关系对植物体内碳平衡的影响,是揭示植物生长和维持机制的关键[26].植物储存的NSC可以用于植物春季萌发生长、冬季植物维持呼吸、缓解水分胁迫等[27].多年生植物NSC的储存特性可以反映植物自身生长及其对环境的适应策略[28].由于常绿树种储存了更多的NSC维持植物体内的碳平衡,通常常绿树种NSC浓度差异较落叶树种NSC浓度差异小[29].落叶树种在早春叶片还没有萌芽阶段,从永久性结构组织中活化NSC供应植物生长和呼吸,从而导致储存器官中NSC含量降低[30].NSC季节变异可以表征植物对水分、光照、温度和养分条件的适应策略[26].在干旱胁迫条件下植物会利用储存的NSC满足植物维持代谢需要、水势适应、组织修复以及应对病原体灾害[28]. ...
Belowground carbon trade among tall trees in a temperate forest
7
2016
... 除光合作用合成碳水化合物外,植物间地下碳交换是植物获取碳源的另外一种重要途径,特别是在环境胁迫或人为干扰条件下,地下碳交换的重要性已凸显.已有研究结果表明,地下碳交换的方向和量级等存在很大差异[31,32].通过对挪威云杉大树进行长期连续的原位标记研究发现,地下碳交换在细的根总碳量中占比达到40[31],而对花旗松幼苗开展的控制条件下脉冲标记研究的结果表明,地下碳交换只占到整个标记期间总碳量的0.1[32].将IRIS技术与冠层透明箱脉冲标记试验结合可以理解碳产生、分配、转移和释放过程的动态变化,解析环境和生理要素等的影响机制[33]. ...
... [31],而对花旗松幼苗开展的控制条件下脉冲标记研究的结果表明,地下碳交换只占到整个标记期间总碳量的0.1[32].将IRIS技术与冠层透明箱脉冲标记试验结合可以理解碳产生、分配、转移和释放过程的动态变化,解析环境和生理要素等的影响机制[33]. ...
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
... ,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
... ,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Transfer of 13C between paired Douglas-fir seedlings reveals plant kinship effects and uptake of exudates by ectomycorrhizas
2
2017
... 除光合作用合成碳水化合物外,植物间地下碳交换是植物获取碳源的另外一种重要途径,特别是在环境胁迫或人为干扰条件下,地下碳交换的重要性已凸显.已有研究结果表明,地下碳交换的方向和量级等存在很大差异[31,32].通过对挪威云杉大树进行长期连续的原位标记研究发现,地下碳交换在细的根总碳量中占比达到40[31],而对花旗松幼苗开展的控制条件下脉冲标记研究的结果表明,地下碳交换只占到整个标记期间总碳量的0.1[32].将IRIS技术与冠层透明箱脉冲标记试验结合可以理解碳产生、分配、转移和释放过程的动态变化,解析环境和生理要素等的影响机制[33]. ...
... [32].将IRIS技术与冠层透明箱脉冲标记试验结合可以理解碳产生、分配、转移和释放过程的动态变化,解析环境和生理要素等的影响机制[33]. ...
Pulse-labelling trees to study carbon allocation dynamics: A review of methods, current knowledge and future prospects
1
2012
... 除光合作用合成碳水化合物外,植物间地下碳交换是植物获取碳源的另外一种重要途径,特别是在环境胁迫或人为干扰条件下,地下碳交换的重要性已凸显.已有研究结果表明,地下碳交换的方向和量级等存在很大差异[31,32].通过对挪威云杉大树进行长期连续的原位标记研究发现,地下碳交换在细的根总碳量中占比达到40[31],而对花旗松幼苗开展的控制条件下脉冲标记研究的结果表明,地下碳交换只占到整个标记期间总碳量的0.1[32].将IRIS技术与冠层透明箱脉冲标记试验结合可以理解碳产生、分配、转移和释放过程的动态变化,解析环境和生理要素等的影响机制[33]. ...
Soil enzymes in a changing environment: Current knowledge and future directions
2
2013
... 从地表到基岩的土壤微生物及胞外酶驱动的土壤有机碳库的分解与转化是碳生物地球化学循环的周转环节(图1b).植物通过地上凋落物、根系凋落物、根系分泌物向土壤中输入有机质,通过胞外酶的矿化和微生物同化,形成土壤有机质碳库[7,34,35].土壤微生物种群结构和酶活性的改变将会改变土壤碳库的分解及蓄积[36]. ...
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
Microbiology: The life beneath our feet
2
2013
... 从地表到基岩的土壤微生物及胞外酶驱动的土壤有机碳库的分解与转化是碳生物地球化学循环的周转环节(图1b).植物通过地上凋落物、根系凋落物、根系分泌物向土壤中输入有机质,通过胞外酶的矿化和微生物同化,形成土壤有机质碳库[7,34,35].土壤微生物种群结构和酶活性的改变将会改变土壤碳库的分解及蓄积[36]. ...
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
Altered soil microbial community at elevated CO2 leads to loss of soil carbon
1
2007
... 从地表到基岩的土壤微生物及胞外酶驱动的土壤有机碳库的分解与转化是碳生物地球化学循环的周转环节(图1b).植物通过地上凋落物、根系凋落物、根系分泌物向土壤中输入有机质,通过胞外酶的矿化和微生物同化,形成土壤有机质碳库[7,34,35].土壤微生物种群结构和酶活性的改变将会改变土壤碳库的分解及蓄积[36]. ...
The carbon balance of terrestrial ecosystems in China
1
2009
... 土壤有机碳库变化导致陆地碳循环评估具有很大的不确定性[37].凋落物的分解速率受到底物的化学成分、参与分解过程的微生物特征、环境条件(温度、水分等)及酶活性影响[38].稳定性植物分解的残体及腐殖化作用形成的腐殖质,其含量可占土壤碳库的70~80.土壤活体微生物量碳约占土壤总有机碳库的1[39],微生物残体可直接贡献土壤稳定碳库[40].土壤中微生物死亡残体量可达土壤微生物活体生物量的40倍,其对土壤总有机碳贡献率可达25~40[40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
Soil extracellular enzyme activities, soil carbon and nitrogen storage under nitrogen fertilization: A meta-analysis
2
2016
... 土壤有机碳库变化导致陆地碳循环评估具有很大的不确定性[37].凋落物的分解速率受到底物的化学成分、参与分解过程的微生物特征、环境条件(温度、水分等)及酶活性影响[38].稳定性植物分解的残体及腐殖化作用形成的腐殖质,其含量可占土壤碳库的70~80.土壤活体微生物量碳约占土壤总有机碳库的1[39],微生物残体可直接贡献土壤稳定碳库[40].土壤中微生物死亡残体量可达土壤微生物活体生物量的40倍,其对土壤总有机碳贡献率可达25~40[40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Nitrogen fertilization decreases forest soil fungal and bacterial biomass in three long-term experiments
1
2006
... 土壤有机碳库变化导致陆地碳循环评估具有很大的不确定性[37].凋落物的分解速率受到底物的化学成分、参与分解过程的微生物特征、环境条件(温度、水分等)及酶活性影响[38].稳定性植物分解的残体及腐殖化作用形成的腐殖质,其含量可占土壤碳库的70~80.土壤活体微生物量碳约占土壤总有机碳库的1[39],微生物残体可直接贡献土壤稳定碳库[40].土壤中微生物死亡残体量可达土壤微生物活体生物量的40倍,其对土壤总有机碳贡献率可达25~40[40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
The importance of anabolism in microbial control over soil carbon storage
2
2017
... 土壤有机碳库变化导致陆地碳循环评估具有很大的不确定性[37].凋落物的分解速率受到底物的化学成分、参与分解过程的微生物特征、环境条件(温度、水分等)及酶活性影响[38].稳定性植物分解的残体及腐殖化作用形成的腐殖质,其含量可占土壤碳库的70~80.土壤活体微生物量碳约占土壤总有机碳库的1[39],微生物残体可直接贡献土壤稳定碳库[40].土壤中微生物死亡残体量可达土壤微生物活体生物量的40倍,其对土壤总有机碳贡献率可达25~40[40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
... [40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
Soil carbon sequestration and land‐use change: Processes and potential
1
2000
... 土壤有机碳库变化导致陆地碳循环评估具有很大的不确定性[37].凋落物的分解速率受到底物的化学成分、参与分解过程的微生物特征、环境条件(温度、水分等)及酶活性影响[38].稳定性植物分解的残体及腐殖化作用形成的腐殖质,其含量可占土壤碳库的70~80.土壤活体微生物量碳约占土壤总有机碳库的1[39],微生物残体可直接贡献土壤稳定碳库[40].土壤中微生物死亡残体量可达土壤微生物活体生物量的40倍,其对土壤总有机碳贡献率可达25~40[40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
Soil organic carbon sequestration rates by tillage and crop rotation
1
2002
... 土壤有机碳库变化导致陆地碳循环评估具有很大的不确定性[37].凋落物的分解速率受到底物的化学成分、参与分解过程的微生物特征、环境条件(温度、水分等)及酶活性影响[38].稳定性植物分解的残体及腐殖化作用形成的腐殖质,其含量可占土壤碳库的70~80.土壤活体微生物量碳约占土壤总有机碳库的1[39],微生物残体可直接贡献土壤稳定碳库[40].土壤中微生物死亡残体量可达土壤微生物活体生物量的40倍,其对土壤总有机碳贡献率可达25~40[40].虽然普遍认同生态系统是开放的系统,但以往地下生态系统科学研究中土壤深度都比较浅.Post等[41]和West等[42]综述了土地利用变化对土壤有机质的影响,通常土壤采样深度的中值为20 cm,约90的土壤采样深度小于30 cm. ...
Surficial gains and subsoil losses of soil carbon and nitrogen during secondary forest development
1
2014
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Limits to soil carbon stability: Deep, ancient soil carbon decomposition stimulated by new labile organic inputs
1
2016
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Ectomycorrhizae in a soil-weathered granitic bedrock regolith: Linking matrix resources to plants
1
2005
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Changes in substrate availability drive carbon cycle response to chronic warming
1
2017
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Storage and stability of organic carbon in soils as related to depth, occlusion within aggregates, and attachment to minerals
1
2013
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Increasing molecular structural complexity and decreasing nitrogen availability depress the mineralization of organic matter in subtropical forest soils
1
2017
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
Changes in the temperature sensitivity of SOM decomposition with grassland succession: Implications for soil C sequestration
1
2013
... 通常下层土壤体积大,且与表层有机碳周转机制不同,其变化规律可能对区域和全球碳循环产生显著影响[43].土壤深度是一个重要的生物和环境梯度,不同深度土壤物理、化学和生物属性及土壤有机碳质量具有显著的差异[44].植物根系是岩石化学风化的主要驱动力,菌根能够穿透长石和角闪石,可能通过有机酸溶解,释放岩石内的养分,供生物吸收[45].深层岩石的化学组成能够对植物生物量和生态系统自身演化产生巨大的影响[9].不同来源有机质矿化速率受微生物量、胞外酶活性、底物数量和质量以及环境条件影响[38,46].随着土壤深度增加,土壤有机质的化学组成、周转时间、降解程度和微生物环境会发生明显改变[47].通常深层土壤有机质含有更多结构复杂和难降解的惰性土壤有机碳组分,土壤有机质矿化速率会降低[48].将IRIS与箱式法技术结合,可在模拟昼夜变化模式下测定土壤碳矿化速率并解析激发效应过程与机制[49]. ...
The contentious nature of soil organic matter
1
2015
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
Advances of soil microbiology in the last decade in China
1
2013
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
中国土壤微生物学研究10年回顾
1
2013
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
Tree species select diverse soil fungal communities expressing different sets of lignocellulolytic enzyme-encoding genes
1
2016
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
Spatial distribution and catalytic mechanisms of β-glucosidase activity at the root-soil interface
1
2016
... 土壤碳库储量和稳定性与微生物活性显著相关,且不同类群的微生物对不同类型有机质分解的贡献存在差异,研究岩石—土壤—生物过程必须考虑土壤微生物群落的功能性作用[50].土壤微生物常被比作有机质中碳与养分元素循环的“转化器”,是控制有机质周转的重要因子[7,51],而土壤酶活性被作为表征有机质中碳氮磷分解的敏感指标[34].土壤微生物群落组成的变化受土壤深度影响比受地理位置和植被类型影响更显著.不同树种,由于其凋落物的质量不同,导致土壤中的真菌功能群不同,分泌的木质纤维素酶不同,影响着土壤碳循环过程[52].土壤根际热点区的碳循环相关酶活性比非根际土壤高近20倍[53]. ...
Contrasting responses of phosphatase kinetic parameters to nitrogen and phosphorus additions in forest soils
1
2018
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
Nitrogen functional gene activity in soil profiles under progressive vegetative recovery after abandonment of agriculture at the Puding Karst Critical Zone Observatory, SW China
1
2018
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
Illumina metabarcoding of a soil fungal community
3
2013
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Improved performance of the PacBio SMRT technology for 16S rDNA sequencing
3
2014
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
DNA stable-isotope probing
3
2007
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Soil biogeochemical processes within the critical zone
2
2007
... 土壤中绝大多数微生物多样性与功能的认识还不清楚,被视为“生物暗物质”[35,54].以往研究多集中在对不同土壤深度、植被类型等对氮循环功能微生物的影响方向[55],目前对碳循环功能微生物丰度与种群多样性在基岩—土壤连续体分布特征还不清楚.随着稳定同位素示踪[31]、高通量测序[56]、三代测序技术[57]和DNA—稳定同位素探针[58]等技术应用,使针对土壤碳库分解和转化的微生物驱动机制研究成为可能,宏基因组学旨在从土壤样本的基因组数据中筛选功能和结构信息,可用于确定物种种类、代谢功能和特定生物地球化学过程之间的关联[59]. ...
... 土壤碳损失除了以CO2气态形式直接排放以外,主要以地表径流和碳淋溶(壤中流和地下径流)的途径流失[8,66].土壤碳损失主要受CO2通量[250~1 220 g C/(m2·a)]影响,DOC淋溶引起的碳损失较少[7.0~9.4 g C/(m2·a)][66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
Roots and associated fungi drive long-term carbon sequestration in boreal forest
1
2013
... 土壤有机质分解所释放的CO2 δ13C包含了土壤有机碳、微生物碳和微生物呼吸CO2等来源的δ13C信号[13].森林生态系统中植物生育期内根际碳释放进入土壤有机质库的数量大于地上凋落物[60],这部分根际碳将携带特殊的δ13C特征(13C贫化)进入土壤有机质库,若被微生物快速分解则将影响δ13C.土壤中木质素的δ13C相对于纤维素贫化6,土壤有机质δ13C贫化和富集的程度,依赖于纤维素和木质素之间最初的δ13C差异以及影响纤维素和木质素分解速率的因子[11].通过IRIS和箱式法技术,结合简单二元混合模型,可将土壤呼吸区分为自养呼吸和异养呼吸两部分[61];利用三元混合模型,将地中海暖温带森林地表排放CO2的源拆分为3个来源,即根、凋落物和土壤碳[62]. ...
Measuring the 13C content of soil‐respired CO2 using a novel open chamber system
1
2008
... 土壤有机质分解所释放的CO2 δ13C包含了土壤有机碳、微生物碳和微生物呼吸CO2等来源的δ13C信号[13].森林生态系统中植物生育期内根际碳释放进入土壤有机质库的数量大于地上凋落物[60],这部分根际碳将携带特殊的δ13C特征(13C贫化)进入土壤有机质库,若被微生物快速分解则将影响δ13C.土壤中木质素的δ13C相对于纤维素贫化6,土壤有机质δ13C贫化和富集的程度,依赖于纤维素和木质素之间最初的δ13C差异以及影响纤维素和木质素分解速率的因子[11].通过IRIS和箱式法技术,结合简单二元混合模型,可将土壤呼吸区分为自养呼吸和异养呼吸两部分[61];利用三元混合模型,将地中海暖温带森林地表排放CO2的源拆分为3个来源,即根、凋落物和土壤碳[62]. ...
In Dual-Chamber Measurements of δ13C of Soil-respired CO2 Partitioned Using a Field-based Three End-member Model
1
2012
... 土壤有机质分解所释放的CO2 δ13C包含了土壤有机碳、微生物碳和微生物呼吸CO2等来源的δ13C信号[13].森林生态系统中植物生育期内根际碳释放进入土壤有机质库的数量大于地上凋落物[60],这部分根际碳将携带特殊的δ13C特征(13C贫化)进入土壤有机质库,若被微生物快速分解则将影响δ13C.土壤中木质素的δ13C相对于纤维素贫化6,土壤有机质δ13C贫化和富集的程度,依赖于纤维素和木质素之间最初的δ13C差异以及影响纤维素和木质素分解速率的因子[11].通过IRIS和箱式法技术,结合简单二元混合模型,可将土壤呼吸区分为自养呼吸和异养呼吸两部分[61];利用三元混合模型,将地中海暖温带森林地表排放CO2的源拆分为3个来源,即根、凋落物和土壤碳[62]. ...
A connection to deep groundwater alters ecosystem carbon fluxes and budgets: Example from a Costa Rican rainforest
1
2013
... 土壤和小流域水体有机和无机碳的迁移过程是碳生物地球化学循环的迁移环节(图1c),主要包括:以地表径流和碳淋溶(壤中流、地下径流)的途径进入流域水体[63];进入流域水体的碳在光合/呼吸、溶解/脱溶、底泥释放/沉积等过程的影响下随流域水体流出流域[64].土壤和小流域水体中的碳主要包括溶解有机碳(Dissolved Organic Carbon,DOC)、颗粒有机碳(Particulate Organic Carbon,POC)、溶解无机碳(Dissolved Inorganic Carbon,DIC)和颗粒无机碳(Particulate Inorganic Carbon,PIC)[65]. ...
Riverine carbon fluxes to the South China Sea
1
2017
... 土壤和小流域水体有机和无机碳的迁移过程是碳生物地球化学循环的迁移环节(图1c),主要包括:以地表径流和碳淋溶(壤中流、地下径流)的途径进入流域水体[63];进入流域水体的碳在光合/呼吸、溶解/脱溶、底泥释放/沉积等过程的影响下随流域水体流出流域[64].土壤和小流域水体中的碳主要包括溶解有机碳(Dissolved Organic Carbon,DOC)、颗粒有机碳(Particulate Organic Carbon,POC)、溶解无机碳(Dissolved Inorganic Carbon,DIC)和颗粒无机碳(Particulate Inorganic Carbon,PIC)[65]. ...
Terrestrial and fluvial carbon fluxes in a tropical watershed: Nyong Basin, Cameroon
1
2009
... 土壤和小流域水体有机和无机碳的迁移过程是碳生物地球化学循环的迁移环节(图1c),主要包括:以地表径流和碳淋溶(壤中流、地下径流)的途径进入流域水体[63];进入流域水体的碳在光合/呼吸、溶解/脱溶、底泥释放/沉积等过程的影响下随流域水体流出流域[64].土壤和小流域水体中的碳主要包括溶解有机碳(Dissolved Organic Carbon,DOC)、颗粒有机碳(Particulate Organic Carbon,POC)、溶解无机碳(Dissolved Inorganic Carbon,DIC)和颗粒无机碳(Particulate Inorganic Carbon,PIC)[65]. ...
Climatic and landscape influences on soil moisture are primary determinants of soil carbon fluxes in seasonally snow-covered forest ecosystems
2
2015
... 土壤碳损失除了以CO2气态形式直接排放以外,主要以地表径流和碳淋溶(壤中流和地下径流)的途径流失[8,66].土壤碳损失主要受CO2通量[250~1 220 g C/(m2·a)]影响,DOC淋溶引起的碳损失较少[7.0~9.4 g C/(m2·a)][66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
... [66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
Equilibration of the terrestrial water, nitrogen, and carbon cycles: Advocating a health threshold for carbon storage
2
2013
... 土壤碳损失除了以CO2气态形式直接排放以外,主要以地表径流和碳淋溶(壤中流和地下径流)的途径流失[8,66].土壤碳损失主要受CO2通量[250~1 220 g C/(m2·a)]影响,DOC淋溶引起的碳损失较少[7.0~9.4 g C/(m2·a)][66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
... 水文过程通过激发碳在剖面、坡地以及流域的物理和化学过程交换,控制着陆地生态系统碳水平输出和生物地球化学循环[67].例如,Johnson等[75]研究表明亚马逊源头溪流的CO2通量代表了深层土壤呼吸的显著周转过程.在湿润季节,深层土壤(2~8 m)CO2浓度的剖面分布表明CO2气体向土壤深层扩散,地下CO2随水出露地表后并未全部排放到大气中,而是以可溶性CO2的形式随水向下游运输,从而对下游地区碳周转产生影响.目前整个小流域尺度水土过程的碳流失通量较难直接观测[76].定量估算水土过程的碳流失通量的基本方法是通过监测研究区域的土壤理化性质、植物生长状况、径流和泥沙携带碳含量等进行估算.随径流运移的流域水体碳输出通量可通过碳含量与流量的乘积计算得到,而流域水体通过光合/呼吸、溶解/脱溶、底泥释放/沉积等过程释放的CO2较难准确直接测定[77].利用δ13C结合相关地球化学参数,如二氧化碳分压(pCO2),DIC和HCO的浓度及阴阳离子等参量反演各端元的相对贡献己经成为研究河流碳的控制因素的重要手段[78]. ...
Cold winter soils enhance dissolved organic carbon concentrations in soil and stream water
1
2010
... 土壤碳损失除了以CO2气态形式直接排放以外,主要以地表径流和碳淋溶(壤中流和地下径流)的途径流失[8,66].土壤碳损失主要受CO2通量[250~1 220 g C/(m2·a)]影响,DOC淋溶引起的碳损失较少[7.0~9.4 g C/(m2·a)][66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
Old carbon mobilized
1
2015
... 土壤碳损失除了以CO2气态形式直接排放以外,主要以地表径流和碳淋溶(壤中流和地下径流)的途径流失[8,66].土壤碳损失主要受CO2通量[250~1 220 g C/(m2·a)]影响,DOC淋溶引起的碳损失较少[7.0~9.4 g C/(m2·a)][66].不同土地利用类型及坡面位置,影响着水土界面的水文和生物地球化学循环过程,进而影响通过水土过程流入流域水体的有机和无机碳[67].水流过土壤的孔隙网络,与多种暴露的土壤表面相接触,影响溶质进入或者移出溶液,在小流域尺度影响着地表水和地下水的质量[59].土壤表层中DOC和DIC与土壤碳含量、微生物活性、水文径流和土壤扰动有关[68].不同土地利用类型产生和输出的泥沙及碳流失量有很大差别,合理的土地利用方式可改善土壤结构,增加土壤对外界环境变化的抵抗力,而不合理的土地利用方式则会导致土壤质量下降,加速侵蚀[69]. ...
Dissolved carbon leaching from soil is a crucial component of the net ecosystem carbon balance
1
2011
... 土壤碳淋溶在生态系统碳平衡估算中占据重要比例,忽视碳淋溶的碳平衡模型存在很大的不确定性[70].碳淋溶过程的控制机制可分为物理化学机制和微生物机制.控制土壤碳淋溶过程的物理化学机制,包括土壤酸碱性和土壤次生矿物对DOC的吸附解吸和优先流等因素[71].土壤酸化导致的土壤DOC淋溶增加对溪流DOC的贡献可达46~126[72];而在富含Fe和Al的土壤中,土壤对溶解有机质的吸附作用不可忽视,但土壤对DOC的吸附存在饱和阈值[73].传统研究认为降水与土壤水充分混合后再形成径流,而基于δD和δ18O相关性研究表明,降水进入土壤后可分为植物吸收利用的束缚水以及自由移动形成径流的自由水,影响碳淋溶的迁移过程[74].微生物对土壤碳淋溶过程的控制体现在对植物源碳和土壤碳库的分解与转化过程中.植物源碳大部分被土壤固持补充土壤碳库,较小部分溶解在土壤水中淋溶流失,导致土壤淋溶液中DOC仍主要来源于土壤碳库,植物源碳与土壤碳库在淋溶过程中发生了动态交换[73]. ...
Abiotic solubilization of soil organic matter, a less-seen aspect of dissolved organic matter production
1
2012
... 土壤碳淋溶在生态系统碳平衡估算中占据重要比例,忽视碳淋溶的碳平衡模型存在很大的不确定性[70].碳淋溶过程的控制机制可分为物理化学机制和微生物机制.控制土壤碳淋溶过程的物理化学机制,包括土壤酸碱性和土壤次生矿物对DOC的吸附解吸和优先流等因素[71].土壤酸化导致的土壤DOC淋溶增加对溪流DOC的贡献可达46~126[72];而在富含Fe和Al的土壤中,土壤对溶解有机质的吸附作用不可忽视,但土壤对DOC的吸附存在饱和阈值[73].传统研究认为降水与土壤水充分混合后再形成径流,而基于δD和δ18O相关性研究表明,降水进入土壤后可分为植物吸收利用的束缚水以及自由移动形成径流的自由水,影响碳淋溶的迁移过程[74].微生物对土壤碳淋溶过程的控制体现在对植物源碳和土壤碳库的分解与转化过程中.植物源碳大部分被土壤固持补充土壤碳库,较小部分溶解在土壤水中淋溶流失,导致土壤淋溶液中DOC仍主要来源于土壤碳库,植物源碳与土壤碳库在淋溶过程中发生了动态交换[73]. ...
Acidity controls on dissolved organic carbon mobility in organic soils
1
2012
... 土壤碳淋溶在生态系统碳平衡估算中占据重要比例,忽视碳淋溶的碳平衡模型存在很大的不确定性[70].碳淋溶过程的控制机制可分为物理化学机制和微生物机制.控制土壤碳淋溶过程的物理化学机制,包括土壤酸碱性和土壤次生矿物对DOC的吸附解吸和优先流等因素[71].土壤酸化导致的土壤DOC淋溶增加对溪流DOC的贡献可达46~126[72];而在富含Fe和Al的土壤中,土壤对溶解有机质的吸附作用不可忽视,但土壤对DOC的吸附存在饱和阈值[73].传统研究认为降水与土壤水充分混合后再形成径流,而基于δD和δ18O相关性研究表明,降水进入土壤后可分为植物吸收利用的束缚水以及自由移动形成径流的自由水,影响碳淋溶的迁移过程[74].微生物对土壤碳淋溶过程的控制体现在对植物源碳和土壤碳库的分解与转化过程中.植物源碳大部分被土壤固持补充土壤碳库,较小部分溶解在土壤水中淋溶流失,导致土壤淋溶液中DOC仍主要来源于土壤碳库,植物源碳与土壤碳库在淋溶过程中发生了动态交换[73]. ...
The dynamic exchange of dissolved organic matter percolating through six diverse soils
2
2014
... 土壤碳淋溶在生态系统碳平衡估算中占据重要比例,忽视碳淋溶的碳平衡模型存在很大的不确定性[70].碳淋溶过程的控制机制可分为物理化学机制和微生物机制.控制土壤碳淋溶过程的物理化学机制,包括土壤酸碱性和土壤次生矿物对DOC的吸附解吸和优先流等因素[71].土壤酸化导致的土壤DOC淋溶增加对溪流DOC的贡献可达46~126[72];而在富含Fe和Al的土壤中,土壤对溶解有机质的吸附作用不可忽视,但土壤对DOC的吸附存在饱和阈值[73].传统研究认为降水与土壤水充分混合后再形成径流,而基于δD和δ18O相关性研究表明,降水进入土壤后可分为植物吸收利用的束缚水以及自由移动形成径流的自由水,影响碳淋溶的迁移过程[74].微生物对土壤碳淋溶过程的控制体现在对植物源碳和土壤碳库的分解与转化过程中.植物源碳大部分被土壤固持补充土壤碳库,较小部分溶解在土壤水中淋溶流失,导致土壤淋溶液中DOC仍主要来源于土壤碳库,植物源碳与土壤碳库在淋溶过程中发生了动态交换[73]. ...
... [73]. ...
Global separation of plant transpiration from groundwater and streamflow
1
2015
... 土壤碳淋溶在生态系统碳平衡估算中占据重要比例,忽视碳淋溶的碳平衡模型存在很大的不确定性[70].碳淋溶过程的控制机制可分为物理化学机制和微生物机制.控制土壤碳淋溶过程的物理化学机制,包括土壤酸碱性和土壤次生矿物对DOC的吸附解吸和优先流等因素[71].土壤酸化导致的土壤DOC淋溶增加对溪流DOC的贡献可达46~126[72];而在富含Fe和Al的土壤中,土壤对溶解有机质的吸附作用不可忽视,但土壤对DOC的吸附存在饱和阈值[73].传统研究认为降水与土壤水充分混合后再形成径流,而基于δD和δ18O相关性研究表明,降水进入土壤后可分为植物吸收利用的束缚水以及自由移动形成径流的自由水,影响碳淋溶的迁移过程[74].微生物对土壤碳淋溶过程的控制体现在对植物源碳和土壤碳库的分解与转化过程中.植物源碳大部分被土壤固持补充土壤碳库,较小部分溶解在土壤水中淋溶流失,导致土壤淋溶液中DOC仍主要来源于土壤碳库,植物源碳与土壤碳库在淋溶过程中发生了动态交换[73]. ...
CO2 efflux from Amazonian headwater streams represents a significant fate for deep soil respiration
1
2008
... 水文过程通过激发碳在剖面、坡地以及流域的物理和化学过程交换,控制着陆地生态系统碳水平输出和生物地球化学循环[67].例如,Johnson等[75]研究表明亚马逊源头溪流的CO2通量代表了深层土壤呼吸的显著周转过程.在湿润季节,深层土壤(2~8 m)CO2浓度的剖面分布表明CO2气体向土壤深层扩散,地下CO2随水出露地表后并未全部排放到大气中,而是以可溶性CO2的形式随水向下游运输,从而对下游地区碳周转产生影响.目前整个小流域尺度水土过程的碳流失通量较难直接观测[76].定量估算水土过程的碳流失通量的基本方法是通过监测研究区域的土壤理化性质、植物生长状况、径流和泥沙携带碳含量等进行估算.随径流运移的流域水体碳输出通量可通过碳含量与流量的乘积计算得到,而流域水体通过光合/呼吸、溶解/脱溶、底泥释放/沉积等过程释放的CO2较难准确直接测定[77].利用δ13C结合相关地球化学参数,如二氧化碳分压(pCO2),DIC和HCO的浓度及阴阳离子等参量反演各端元的相对贡献己经成为研究河流碳的控制因素的重要手段[78]. ...
Modeling denitrification in terrestrial and aquatic ecosystems at regional scales
2
2006
... 水文过程通过激发碳在剖面、坡地以及流域的物理和化学过程交换,控制着陆地生态系统碳水平输出和生物地球化学循环[67].例如,Johnson等[75]研究表明亚马逊源头溪流的CO2通量代表了深层土壤呼吸的显著周转过程.在湿润季节,深层土壤(2~8 m)CO2浓度的剖面分布表明CO2气体向土壤深层扩散,地下CO2随水出露地表后并未全部排放到大气中,而是以可溶性CO2的形式随水向下游运输,从而对下游地区碳周转产生影响.目前整个小流域尺度水土过程的碳流失通量较难直接观测[76].定量估算水土过程的碳流失通量的基本方法是通过监测研究区域的土壤理化性质、植物生长状况、径流和泥沙携带碳含量等进行估算.随径流运移的流域水体碳输出通量可通过碳含量与流量的乘积计算得到,而流域水体通过光合/呼吸、溶解/脱溶、底泥释放/沉积等过程释放的CO2较难准确直接测定[77].利用δ13C结合相关地球化学参数,如二氧化碳分压(pCO2),DIC和HCO的浓度及阴阳离子等参量反演各端元的相对贡献己经成为研究河流碳的控制因素的重要手段[78]. ...
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
A review of CO2 and associated carbon dynamics in headwater streams: A global perspective: Carbon dioxide in headwater streams
1
2017
... 水文过程通过激发碳在剖面、坡地以及流域的物理和化学过程交换,控制着陆地生态系统碳水平输出和生物地球化学循环[67].例如,Johnson等[75]研究表明亚马逊源头溪流的CO2通量代表了深层土壤呼吸的显著周转过程.在湿润季节,深层土壤(2~8 m)CO2浓度的剖面分布表明CO2气体向土壤深层扩散,地下CO2随水出露地表后并未全部排放到大气中,而是以可溶性CO2的形式随水向下游运输,从而对下游地区碳周转产生影响.目前整个小流域尺度水土过程的碳流失通量较难直接观测[76].定量估算水土过程的碳流失通量的基本方法是通过监测研究区域的土壤理化性质、植物生长状况、径流和泥沙携带碳含量等进行估算.随径流运移的流域水体碳输出通量可通过碳含量与流量的乘积计算得到,而流域水体通过光合/呼吸、溶解/脱溶、底泥释放/沉积等过程释放的CO2较难准确直接测定[77].利用δ13C结合相关地球化学参数,如二氧化碳分压(pCO2),DIC和HCO的浓度及阴阳离子等参量反演各端元的相对贡献己经成为研究河流碳的控制因素的重要手段[78]. ...
Origin and cycling of riverine inorganic carbon in the Sava River watershed (Slovenia) inferred from major solutes and stable carbon isotopes
1
2007
... 水文过程通过激发碳在剖面、坡地以及流域的物理和化学过程交换,控制着陆地生态系统碳水平输出和生物地球化学循环[67].例如,Johnson等[75]研究表明亚马逊源头溪流的CO2通量代表了深层土壤呼吸的显著周转过程.在湿润季节,深层土壤(2~8 m)CO2浓度的剖面分布表明CO2气体向土壤深层扩散,地下CO2随水出露地表后并未全部排放到大气中,而是以可溶性CO2的形式随水向下游运输,从而对下游地区碳周转产生影响.目前整个小流域尺度水土过程的碳流失通量较难直接观测[76].定量估算水土过程的碳流失通量的基本方法是通过监测研究区域的土壤理化性质、植物生长状况、径流和泥沙携带碳含量等进行估算.随径流运移的流域水体碳输出通量可通过碳含量与流量的乘积计算得到,而流域水体通过光合/呼吸、溶解/脱溶、底泥释放/沉积等过程释放的CO2较难准确直接测定[77].利用δ13C结合相关地球化学参数,如二氧化碳分压(pCO2),DIC和HCO的浓度及阴阳离子等参量反演各端元的相对贡献己经成为研究河流碳的控制因素的重要手段[78]. ...
A future perspective on North America's freshwater ecosystems
1
2000
... 水库等设施的修建将会导致流域水体流速变缓,水体的携带能力下降,由强动力条件下的“河流侵蚀搬运作用”变成“湖泊沉积作用”,大量径流携带的碳沉积在库区内[79],其形成的水域环境有利于浮游植物和水生植物的生长,进而影响流域水体的碳通量[80].当流域中湖泊、水库等水体较多时,将会减缓水流速度,提高停滞时间,同时也减缓了有机碳的氧化分解速度[81].滞留的有机质受蚀搬运时释放大量溶解性物质,使流域水体中DOC/POC比值保持在10以上,接近湖水/水库的比值特征[82].Li等[83]以全球11个内陆河流下游封闭湖泊沉积物数据为基础,推测全球DIC沉积速率为0.152 Pg/a,与深海碳埋藏量相当. ...
Carbon sequestration in a large hydroelectric reservoir: An integrative seismic approach
1
2014
... 水库等设施的修建将会导致流域水体流速变缓,水体的携带能力下降,由强动力条件下的“河流侵蚀搬运作用”变成“湖泊沉积作用”,大量径流携带的碳沉积在库区内[79],其形成的水域环境有利于浮游植物和水生植物的生长,进而影响流域水体的碳通量[80].当流域中湖泊、水库等水体较多时,将会减缓水流速度,提高停滞时间,同时也减缓了有机碳的氧化分解速度[81].滞留的有机质受蚀搬运时释放大量溶解性物质,使流域水体中DOC/POC比值保持在10以上,接近湖水/水库的比值特征[82].Li等[83]以全球11个内陆河流下游封闭湖泊沉积物数据为基础,推测全球DIC沉积速率为0.152 Pg/a,与深海碳埋藏量相当. ...
Geochemical characteristics and fluxes of organic carbon in a human-disturbed mountainous river (the Luodingjiang River) of the Zhujiang (Pearl River), China
1
2009
... 水库等设施的修建将会导致流域水体流速变缓,水体的携带能力下降,由强动力条件下的“河流侵蚀搬运作用”变成“湖泊沉积作用”,大量径流携带的碳沉积在库区内[79],其形成的水域环境有利于浮游植物和水生植物的生长,进而影响流域水体的碳通量[80].当流域中湖泊、水库等水体较多时,将会减缓水流速度,提高停滞时间,同时也减缓了有机碳的氧化分解速度[81].滞留的有机质受蚀搬运时释放大量溶解性物质,使流域水体中DOC/POC比值保持在10以上,接近湖水/水库的比值特征[82].Li等[83]以全球11个内陆河流下游封闭湖泊沉积物数据为基础,推测全球DIC沉积速率为0.152 Pg/a,与深海碳埋藏量相当. ...
Concentrations and characteristics of organic carbon in surface water in Arizona: Influence of urbanization
1
2000
... 水库等设施的修建将会导致流域水体流速变缓,水体的携带能力下降,由强动力条件下的“河流侵蚀搬运作用”变成“湖泊沉积作用”,大量径流携带的碳沉积在库区内[79],其形成的水域环境有利于浮游植物和水生植物的生长,进而影响流域水体的碳通量[80].当流域中湖泊、水库等水体较多时,将会减缓水流速度,提高停滞时间,同时也减缓了有机碳的氧化分解速度[81].滞留的有机质受蚀搬运时释放大量溶解性物质,使流域水体中DOC/POC比值保持在10以上,接近湖水/水库的比值特征[82].Li等[83]以全球11个内陆河流下游封闭湖泊沉积物数据为基础,推测全球DIC沉积速率为0.152 Pg/a,与深海碳埋藏量相当. ...
Substantial inorganic carbon sink in closed drainage basins globally
1
2017
... 水库等设施的修建将会导致流域水体流速变缓,水体的携带能力下降,由强动力条件下的“河流侵蚀搬运作用”变成“湖泊沉积作用”,大量径流携带的碳沉积在库区内[79],其形成的水域环境有利于浮游植物和水生植物的生长,进而影响流域水体的碳通量[80].当流域中湖泊、水库等水体较多时,将会减缓水流速度,提高停滞时间,同时也减缓了有机碳的氧化分解速度[81].滞留的有机质受蚀搬运时释放大量溶解性物质,使流域水体中DOC/POC比值保持在10以上,接近湖水/水库的比值特征[82].Li等[83]以全球11个内陆河流下游封闭湖泊沉积物数据为基础,推测全球DIC沉积速率为0.152 Pg/a,与深海碳埋藏量相当. ...
The response of photosynthesis and stomatal conductance to rising [CO2]: Mechanisms and environmental interactions
1
2007
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
Probing deep weathering in the Shale Hills Critical Zone Observatory, Pennsylvania (USA): The hypothesis of nested chemical reaction fronts in the subsurface
1
2013
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
Continental-scale decrease in net primary productivity in streams due to climate warming
1
2018
... 目前,地球关键带过程与功能是国家自然科学基金委员会地球科学部重点项目领域,其中关键带物质和元素循环的生物地球化学过程、机制及其生态功能是资助方向之一.陆地生态系统对碳的吸收、迁移与转化主要通过垂直尺度与水平尺度的碳输入和输出[84].在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识小流域碳迁移过程与平衡机制[2,3,4].Brantley等[85]提出测量和模拟整个流域上生态系统—大气—地表下岩石间的水、能量和碳交换是地球关键带的关键科学问题之一.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳平衡过程及控制机制研究提供了新的契机,例如,能够揭示从地表到基岩土壤与溪流DOC浓度和性质的差异及其对生态系统碳平衡的贡献,以及土壤DOC产生的微生物调控机制和降水径流、酸碱度及次生矿物等对DOC迁移的影响机制.生物地球化学过程模型通过综合分析观测数据和总结规律,在理论知识验证的基础上,对不同气候、土地利用类型、土壤类型等条件下的物质循环过程进行定量估算,为定量估算流域尺度生态系统碳平衡以及碳利用效率提供了有效工具[76,86]. ...
Whole-tree distribution and temporal variation of non-structural carbohydrates in broadleaf evergreen trees
2
2018
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
... ,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...
Allocation mechanisms of non-structural carbohydrates of robinia pseudoacacia l. Seedlings in response to drought and waterlogging
1
2018
... 在地球关键带科学的框架下,应加强在流域尺度上从冠层到基岩系统认识碳循环的生物地球化学过程、机制及生态功能,系统阐明生态系统植物碳固定及分配机制、从地表到基岩的土壤碳库分解和转化机制及小流域碳迁移过程与机制.稳定同位素[28,31]和土壤微生物[56,57,58]等新技术的进步为生态系统碳迁移和转化过程及控制机制研究提供了新的契机,例如,利用针对植物不同器官的野外原位采样[87,88]以及野外自然条件和室内控制条件下的13C(14C)连续或脉冲标记方法[13,28,31],可以研究植物体内SC和NSC之间分配的权衡关系、植物间地下碳交换过程以及从地表到基岩的土壤碳分解过程和机制;通过设置不同的环境胁迫(如干旱和光照等)和人为干扰(如火烧和砍伐等)条件,探讨生态系统内部碳迁移与转化过程与控制机制[28,31,87];揭示基岩风化养分输入等对生态系统碳过程的影响[9]等. ...





{kind=link}
{kind=link}