Geology: Understanding soil time
1
2008
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
The role of soil organic matter in sustaining soil fertility
1
1994
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
The role of soil organic matter in maintaining the productivity and yield stability of cereals in China
1
2009
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
Assessing the Productivity Function of Soils
1
2011
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
Soil Carbon: Science, Management and Policy for Multiple Benefits
1
2014
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
Biogeochemical cycles and biodiversity as key drivers of ecosystem services provided by soils
2
2015
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
... 从本源上说,土壤物质就是大小、结构和功能(反应性和生物活性)不同的、矿物质—有机质—微生物有序集合的土壤团聚体颗粒集合.其形成、发育和稳定属于土壤的生物动力学过程(biodynamics)[158],是土壤形成及相关生命过程的基础.植物等生物活动剩余物输入的有机物质,一方面作为原始材料驱动土壤有机质形成和稳定;另一方面,它们被土壤中的食物网,特别是微生物分解利用,并提供其活动的能源.而土壤团聚体的形成、稳定和更新提供和维持了有机质—微生物—生物功能的偶联关系及其稳定,是土壤发挥生物地球化学作用和生物多样性保持[6]的根本.因此,土壤有机质连续体实际上是一种保持有机质—微生物—生物活性良好协同的微域机制框架. ...
The benefits of soil carbon
1
2012
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
Advances in soil ecosystem services: Concepts, models, and applications for Earth system life support
1
2013
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
Global soil organic carbon assessment
1
2015
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
Harmonized soil property values for broad-scale modelling (WISE30sec) with estimates of global soil carbon stocks
2
2016
... 土壤有机质是促进土壤发育、提高土壤功能的最原始且最核心的驱动者,因此认识土壤有机质是土壤学的基础和核心理论范畴之一.地球表层岩石风化伊始,生命活动残留物进入土壤转化为土壤有机质,进而推动土壤形成和发育[1].土壤有机质的积累和固定改善了土壤质量并促进土壤功能,这尤其体现在农业生产力和土壤管理的可持续性[2,3,4]方面.土壤有机质作为土壤的关键组成部分,通过对土壤结构发育(biophysical)和地球生物地球化学过程(biogeochemical)的双重控制,对各种土壤过程起到启动和调节作用,保障土壤提供生物量生产、能源生产、生物多样性保持、水分蓄持及固碳减排等多种生态系统服务[5].并且,以土壤有机质为媒介的生物地球化学过程及生物多样性驱动着土壤提供多种生态系统服务[6].有充分的理由相信,土壤有机质是地球可持续发展的关键资源和自然资本[7,8],无疑是评价土壤自然价值的核心[9,10]. ...
... 全球土壤数据整合计量表明,全球土壤有机碳(平均占土壤有机质58%)储量可达1 500 Pg (1 m深)和2 200 Pg(2 m深)[10],这是大气碳储量和植被生物量碳储量的数倍.因此,土壤有机质管理在调节气候变化和生态系统(特别是农田)生产力上具有举足轻重的地位[11,12].面向日益严峻的气候变化管控和日益增长的食物需求的农业生产,土壤有机质研究一方面走向空间可分辨的、微生物机理明确的、矿物—植物交互影响的有机质过程;另一方面走向为全球治理服务的价值观和政策性管理整合分析及模型升级[13].深入认识和刻画土壤有机质的起源、组分、结构与土壤功能及服务的关系(简称结构—功能关系),解析土壤有机质组分与土壤中生物和非生物因子的相互作用关系,阐明土壤有机质结构—功能与土壤形成演化、有机碳库固定和稳定及其与生物多样性发育、保持及生态系统服务功能协同的土壤学机制,仍将是土壤学领域基础研究的核心任务[14].为此,本文梳理近20年来土壤有机质研究的基本路径,分析讨论土壤有机质研究的若干进展与关键科学认识,展望未来土壤有机质研究的前沿,以期为中国土壤学研究提供参考. ...
Soil carbon sequestration impacts on global climate change and food security
1
2004
... 全球土壤数据整合计量表明,全球土壤有机碳(平均占土壤有机质58%)储量可达1 500 Pg (1 m深)和2 200 Pg(2 m深)[10],这是大气碳储量和植被生物量碳储量的数倍.因此,土壤有机质管理在调节气候变化和生态系统(特别是农田)生产力上具有举足轻重的地位[11,12].面向日益严峻的气候变化管控和日益增长的食物需求的农业生产,土壤有机质研究一方面走向空间可分辨的、微生物机理明确的、矿物—植物交互影响的有机质过程;另一方面走向为全球治理服务的价值观和政策性管理整合分析及模型升级[13].深入认识和刻画土壤有机质的起源、组分、结构与土壤功能及服务的关系(简称结构—功能关系),解析土壤有机质组分与土壤中生物和非生物因子的相互作用关系,阐明土壤有机质结构—功能与土壤形成演化、有机碳库固定和稳定及其与生物多样性发育、保持及生态系统服务功能协同的土壤学机制,仍将是土壤学领域基础研究的核心任务[14].为此,本文梳理近20年来土壤有机质研究的基本路径,分析讨论土壤有机质研究的若干进展与关键科学认识,展望未来土壤有机质研究的前沿,以期为中国土壤学研究提供参考. ...
Climate-smart soils
1
2016
... 全球土壤数据整合计量表明,全球土壤有机碳(平均占土壤有机质58%)储量可达1 500 Pg (1 m深)和2 200 Pg(2 m深)[10],这是大气碳储量和植被生物量碳储量的数倍.因此,土壤有机质管理在调节气候变化和生态系统(特别是农田)生产力上具有举足轻重的地位[11,12].面向日益严峻的气候变化管控和日益增长的食物需求的农业生产,土壤有机质研究一方面走向空间可分辨的、微生物机理明确的、矿物—植物交互影响的有机质过程;另一方面走向为全球治理服务的价值观和政策性管理整合分析及模型升级[13].深入认识和刻画土壤有机质的起源、组分、结构与土壤功能及服务的关系(简称结构—功能关系),解析土壤有机质组分与土壤中生物和非生物因子的相互作用关系,阐明土壤有机质结构—功能与土壤形成演化、有机碳库固定和稳定及其与生物多样性发育、保持及生态系统服务功能协同的土壤学机制,仍将是土壤学领域基础研究的核心任务[14].为此,本文梳理近20年来土壤有机质研究的基本路径,分析讨论土壤有机质研究的若干进展与关键科学认识,展望未来土壤有机质研究的前沿,以期为中国土壤学研究提供参考. ...
The changing faces of soil organic matter research
1
2018
... 全球土壤数据整合计量表明,全球土壤有机碳(平均占土壤有机质58%)储量可达1 500 Pg (1 m深)和2 200 Pg(2 m深)[10],这是大气碳储量和植被生物量碳储量的数倍.因此,土壤有机质管理在调节气候变化和生态系统(特别是农田)生产力上具有举足轻重的地位[11,12].面向日益严峻的气候变化管控和日益增长的食物需求的农业生产,土壤有机质研究一方面走向空间可分辨的、微生物机理明确的、矿物—植物交互影响的有机质过程;另一方面走向为全球治理服务的价值观和政策性管理整合分析及模型升级[13].深入认识和刻画土壤有机质的起源、组分、结构与土壤功能及服务的关系(简称结构—功能关系),解析土壤有机质组分与土壤中生物和非生物因子的相互作用关系,阐明土壤有机质结构—功能与土壤形成演化、有机碳库固定和稳定及其与生物多样性发育、保持及生态系统服务功能协同的土壤学机制,仍将是土壤学领域基础研究的核心任务[14].为此,本文梳理近20年来土壤有机质研究的基本路径,分析讨论土壤有机质研究的若干进展与关键科学认识,展望未来土壤有机质研究的前沿,以期为中国土壤学研究提供参考. ...
The knowns, known unknowns and unknowns of sequestration of soil organic carbon
1
2013
... 全球土壤数据整合计量表明,全球土壤有机碳(平均占土壤有机质58%)储量可达1 500 Pg (1 m深)和2 200 Pg(2 m深)[10],这是大气碳储量和植被生物量碳储量的数倍.因此,土壤有机质管理在调节气候变化和生态系统(特别是农田)生产力上具有举足轻重的地位[11,12].面向日益严峻的气候变化管控和日益增长的食物需求的农业生产,土壤有机质研究一方面走向空间可分辨的、微生物机理明确的、矿物—植物交互影响的有机质过程;另一方面走向为全球治理服务的价值观和政策性管理整合分析及模型升级[13].深入认识和刻画土壤有机质的起源、组分、结构与土壤功能及服务的关系(简称结构—功能关系),解析土壤有机质组分与土壤中生物和非生物因子的相互作用关系,阐明土壤有机质结构—功能与土壤形成演化、有机碳库固定和稳定及其与生物多样性发育、保持及生态系统服务功能协同的土壤学机制,仍将是土壤学领域基础研究的核心任务[14].为此,本文梳理近20年来土壤有机质研究的基本路径,分析讨论土壤有机质研究的若干进展与关键科学认识,展望未来土壤有机质研究的前沿,以期为中国土壤学研究提供参考. ...
The Nature and Properties of Soils(8th ed)
1
1974
... 经典土壤学将土壤有机质定义为土壤中的生命体及其死亡的生物质残留和腐殖质.前者也可认为是非腐殖质,代表没有降解或者降解残留的植物源有机物质.腐殖质包括腐殖物质和非腐殖物质(利用和分解有机质留下的微生物源物质).在一系列不同的pH和电解质浓度下提取和分离操作的方法基础上,经典土壤腐殖质理论认为腐殖物质包含3个最基本和重要的组分:分子量较小而化学性质较活泼的富里酸,分子量较大且以杂环结构为特征的复杂胡敏酸以及高度缩合、惰性的胡敏素等.这些组分是土壤有机质中数量较多、性质稳定的复杂有机物质,其分子量可介于数百到数十万道尔顿(dalton),化学结构具有高度异质性[15]. ...
Plant Litter: Decomposition, Humus Formation, Carbon Sequestration (2nd ed)
1
2007
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
The humus profile-concept, class and reality
1
1995
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
Elements of the Nature and Properties of Soils (2nd ed)
1
2004
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
Humus Chemistry: Genesis, Composition, Reactions (2nd ed)
1
1994
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
Are the traditional concepts of the structures of humic substances realistic?
2
2001
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
... [20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
Humic matter in soil and the environment, principles and controversies
1
2015
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
Humification and mineralization in soils
1
2005
... 土壤腐殖质形成理论先后有植物有机物降解假设(plant alteration hypothesis,植物有机物降解残留的木质素—蛋白质为核的有机大分子聚合物)、微生物合成假设(microbial synthesis hypothesis, 微生物代谢活动在其细胞内合成大分子的腐殖物质聚合物、死亡后释放到土壤中)、化学多聚化假设(chemical polymerization, 微生物产生的胺基化合物、酚类化合物等单体在土壤中发生多聚化而形成腐殖物质)以及细胞自溶假设(cell autolysis hypothesis,土壤中生物死亡后细胞自体崩解而溶析出氨基酸、糖类和苯羧酸等分子,后者多聚化形成腐殖物质)[16,17].经典腐殖质理论的核心是生物源有机物的腐殖化过程:新鲜有机质矿化—残余物二次合成—聚合与缩合—腐殖质形成.腐殖质合成的本质是以含氧芳香烃为核心,有机质降解中间产物与其聚合进一步合成大分子多聚体(往往称为二次合成),最终很可能以含氧芳香烃及其衍生物为基础,外接不同碳链的脂肪族烷烃分子而链接为巨大聚合物[18],这种大分子聚合体通过金属离子桥键结合到矿物表面,成为有机矿质复合体(被认为是腐殖物质的超分子结构本质)而稳定于土壤,使腐殖化过程具有了生态学意义[19].但这种聚合作用和桥接稳定作用的热力学过程还很不清楚,其反应并不符合能量降低规律,故不断受到土壤有机质研究者的质疑[20,21,22].Burdon[20]指出,现代分子检测技术无法直接检测出具有完整分子结构和形态的腐殖组分,假设的有机质矿化产物发生腐殖化反应的“二次合成”所需的能源来源和途径也缺乏足够的热力学证据. ...
Analytical approaches for characterizing soil organic matter
1
2000
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Comparison of labile soil organic matter fractionation techniques
2
2004
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
Indications for soil organic matter quality in soils under different management
2
2002
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
A state of the art structural concept for humic substances
2
1993
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
... [26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
New insights into organic-mineral particles: Composition, properties and models of molecular structure
1
2000
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Solid-state 13C NMR of sedimentary humic substances: New revelations on their chemical composition
1
1983
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Spectroscopic characterization of humic acid fractions isolated from soil using different extraction procedures
2
2006
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
... [29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
NMR characterization of humic acid fractions from different Philippine soils and sediments
1
1990
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Thermochemolysis of genetically different soil humic acids and their fractions obtained by tandem size exclusion chromatography-polyacrylamide gel electrophoresis
1
2006
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Chemical characterization of soil organic-matter fractions by analytical pyrolysis -Gas Chromatography-Mass spectrometry
1
1986
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Quantitative evaluation of XAD-8 and XAD-4 resins used in tandem for removing organic solutes from water
1
1992
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Leaching of Dissolved Organic Carbon (DOC) and Dissolved Organic Nitrogen (DON) in mor humus as affected by temperature and pH
1
2000
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
Historical evolution of soil organic matter concepts and their relationships with the fertility and sustainability of cropping systems
1
2007
... 20世纪90年代以来,傅里叶变换红外光谱、核磁共振和热裂解—质谱仪等光谱/波谱/色谱学结合分析技术引入土壤有机质组分研究,得以在功能团组成上认识土壤有机质的复杂性和多元性[23,24,25].Schulten等[26]提出了胡敏酸分子化学结构模型,他们采用13C-NMR波谱仪方法和热解方法推断出胡敏酸分子的核心结构是烷基芳香烃,缺少含氧官能团,并且胡敏酸有机大分子与土壤矿物质结合构成了化学结构上特定的矿质有机复合体[27].但后来的一系列提取和热解质谱分析研究[28,29],胡敏酸组分的含氧基团组分比例其实很高,显然不支持Schulten等[26]对胡敏酸分子结构的认识.Wershaw等[30]采集了菲律宾不同地点的泥炭土、耕地和湖泊沉积物样品,通过吸附—色谱分离法分离胡敏酸组分,进一步鉴定其核磁共振官能团组成,表明不同来源的胡敏酸组分都含有来自植物组织的木质素化合物、碳水化合物和脂肪族化合物,而不同样本中基团的化学位移差异代表了这些植物起源化合物的结构差异,特别是碳水化合物可以直接指示不同的植物来源.Saiz-Jimenez等[31]采用凝胶渗透层析色谱法和聚乙酰胺凝胶电泳耦合法对灰土、软土、始成土和老成土的胡敏酸进行分组,进一步通过对其热解(thermolysis)产物进行气质联用分析,表明胡敏酸热解产物的电泳移动性组分结构在不同土壤间相似,强电泳移动性组分主要是植物脂肪酸和蜡质正构烷烃,中等电泳移动性的组分是一定程度降解的木质素,而木质素大分子富集于弱电泳性组分,这种组分结构的组成成分并不因土壤发生改变而改变.Saiz-Jimenez等[32]采用13C-CPMAS-NMR技术对草地土壤和灌木林土壤的比较分析表明,2种土壤的胡敏酸、富里酸和胡敏素等组分的分子组成结构存在相似性,尽管富里酸的芳构化度低于胡敏酸,但胡敏酸与胡敏素的芳构度接近.这些均指示了胡敏酸和胡敏素并不一定是2种不同类型的腐殖物质,而组成简单的富里酸更不是胡敏酸的降构产物.20世纪90年代初,XAD-8树脂(常与XAD-4联用) 应用于分离制取水溶性天然有机质或土壤中可溶性有机质,这个物理迁移性和化学反应性都很活泼的组分并不只含有亲水性有机酸,居然还含有疏水性(芳香碳)有机酸与中性有机物,当然分子量及组成比例变化范围很宽[29,33,34].由此看来,腐殖质学说的胡敏酸、富里酸及胡敏素等核心组分都包含酚和部分氧化的木质素单体复合构成的网状结构,它们的差异仅可能有以下2个原因:吸附或结合于这种结构表面的羧酸类、脂肪类等有机功能团的相对丰度不同;在土壤中的结合状态不同,在土壤中并不存在腐殖物质大分子单体.总之,21世纪日益活跃的分子结构研究,并不支持土壤有机质由大分子化合物组成的腐殖质学说,尤其是不能肯定土壤中存在持久稳定的“腐殖物质”分子[35]. ...
The macromolecular organic composition of plant and microbial residues as inputs to soil organic matter
1
2002
... 与腐殖质学说相交存在的另一种观点是,土壤有机质是植物来源有机物的残留物.20世纪30年代,已经有不同分解阶段的植物残留有机物的化学组成资料.到20世纪末,对于土壤中不同植物来源有机分子的存在及其分解性已经有详尽的资料[36].这些未分解、部分分解和正在分解转化中的有机组分构成了瞬时的土壤有机质主体,当然它们处于连续的动态变化中.之前研究认为,真菌促进了新鲜有机物分解而使其残留物成为土壤有机质.例如,植物蛋白质降解产生的氨基酸在土壤中残留而形成了稳定的含氮有机物质.后来,这种有机质残留被认为是土壤微生物在完成分解后“选择性保留”于土壤的结果[37,38,39],是土壤微生物的“自私行为”.作为优先性分解组分,来自植物残体和叶片的有机物质快速分解损失,而来自根部的植物有机质倾向于保留在土壤中[40],使土壤表现出对根有机物的“偏好”(亲合性).Rasse等[41]指出,根系来源的有机化合物实际上是土壤有机质积累和固定的主要来源,他们认为来自根系的有机质比来自地上部的有机质更易被土壤固定而保留于土壤中,这是因为根系活动通过真菌菌根、根毛和分泌物的矿质(金属)结合、团聚体物理保护等机制来促进根系源物质的优先残留保持,而不是由稳定性化学转化而决定的.上述的植物源芳香化合物的可溶性组分被矿质优先吸附和固定也符合选择性残留保持机制. ...
Nitrogen and lignin control of hardwood leaf litter decomposition dynamics
2
1982
... 与腐殖质学说相交存在的另一种观点是,土壤有机质是植物来源有机物的残留物.20世纪30年代,已经有不同分解阶段的植物残留有机物的化学组成资料.到20世纪末,对于土壤中不同植物来源有机分子的存在及其分解性已经有详尽的资料[36].这些未分解、部分分解和正在分解转化中的有机组分构成了瞬时的土壤有机质主体,当然它们处于连续的动态变化中.之前研究认为,真菌促进了新鲜有机物分解而使其残留物成为土壤有机质.例如,植物蛋白质降解产生的氨基酸在土壤中残留而形成了稳定的含氮有机物质.后来,这种有机质残留被认为是土壤微生物在完成分解后“选择性保留”于土壤的结果[37,38,39],是土壤微生物的“自私行为”.作为优先性分解组分,来自植物残体和叶片的有机物质快速分解损失,而来自根部的植物有机质倾向于保留在土壤中[40],使土壤表现出对根有机物的“偏好”(亲合性).Rasse等[41]指出,根系来源的有机化合物实际上是土壤有机质积累和固定的主要来源,他们认为来自根系的有机质比来自地上部的有机质更易被土壤固定而保留于土壤中,这是因为根系活动通过真菌菌根、根毛和分泌物的矿质(金属)结合、团聚体物理保护等机制来促进根系源物质的优先残留保持,而不是由稳定性化学转化而决定的.上述的植物源芳香化合物的可溶性组分被矿质优先吸附和固定也符合选择性残留保持机制. ...
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Predicting long-term patterns of mass loss, nitrogen dynamics, and soil organic matter formation from initial fine litter chemistry in temperate forest ecosystems
1
1990
... 与腐殖质学说相交存在的另一种观点是,土壤有机质是植物来源有机物的残留物.20世纪30年代,已经有不同分解阶段的植物残留有机物的化学组成资料.到20世纪末,对于土壤中不同植物来源有机分子的存在及其分解性已经有详尽的资料[36].这些未分解、部分分解和正在分解转化中的有机组分构成了瞬时的土壤有机质主体,当然它们处于连续的动态变化中.之前研究认为,真菌促进了新鲜有机物分解而使其残留物成为土壤有机质.例如,植物蛋白质降解产生的氨基酸在土壤中残留而形成了稳定的含氮有机物质.后来,这种有机质残留被认为是土壤微生物在完成分解后“选择性保留”于土壤的结果[37,38,39],是土壤微生物的“自私行为”.作为优先性分解组分,来自植物残体和叶片的有机物质快速分解损失,而来自根部的植物有机质倾向于保留在土壤中[40],使土壤表现出对根有机物的“偏好”(亲合性).Rasse等[41]指出,根系来源的有机化合物实际上是土壤有机质积累和固定的主要来源,他们认为来自根系的有机质比来自地上部的有机质更易被土壤固定而保留于土壤中,这是因为根系活动通过真菌菌根、根毛和分泌物的矿质(金属)结合、团聚体物理保护等机制来促进根系源物质的优先残留保持,而不是由稳定性化学转化而决定的.上述的植物源芳香化合物的可溶性组分被矿质优先吸附和固定也符合选择性残留保持机制. ...
Evidence for non-selective preservation of organic matter in sinking marine particles
1
2001
... 与腐殖质学说相交存在的另一种观点是,土壤有机质是植物来源有机物的残留物.20世纪30年代,已经有不同分解阶段的植物残留有机物的化学组成资料.到20世纪末,对于土壤中不同植物来源有机分子的存在及其分解性已经有详尽的资料[36].这些未分解、部分分解和正在分解转化中的有机组分构成了瞬时的土壤有机质主体,当然它们处于连续的动态变化中.之前研究认为,真菌促进了新鲜有机物分解而使其残留物成为土壤有机质.例如,植物蛋白质降解产生的氨基酸在土壤中残留而形成了稳定的含氮有机物质.后来,这种有机质残留被认为是土壤微生物在完成分解后“选择性保留”于土壤的结果[37,38,39],是土壤微生物的“自私行为”.作为优先性分解组分,来自植物残体和叶片的有机物质快速分解损失,而来自根部的植物有机质倾向于保留在土壤中[40],使土壤表现出对根有机物的“偏好”(亲合性).Rasse等[41]指出,根系来源的有机化合物实际上是土壤有机质积累和固定的主要来源,他们认为来自根系的有机质比来自地上部的有机质更易被土壤固定而保留于土壤中,这是因为根系活动通过真菌菌根、根毛和分泌物的矿质(金属)结合、团聚体物理保护等机制来促进根系源物质的优先残留保持,而不是由稳定性化学转化而决定的.上述的植物源芳香化合物的可溶性组分被矿质优先吸附和固定也符合选择性残留保持机制. ...
Stabilization and destabilization of soil organic matter: Mechanisms and controls
1
1996
... 与腐殖质学说相交存在的另一种观点是,土壤有机质是植物来源有机物的残留物.20世纪30年代,已经有不同分解阶段的植物残留有机物的化学组成资料.到20世纪末,对于土壤中不同植物来源有机分子的存在及其分解性已经有详尽的资料[36].这些未分解、部分分解和正在分解转化中的有机组分构成了瞬时的土壤有机质主体,当然它们处于连续的动态变化中.之前研究认为,真菌促进了新鲜有机物分解而使其残留物成为土壤有机质.例如,植物蛋白质降解产生的氨基酸在土壤中残留而形成了稳定的含氮有机物质.后来,这种有机质残留被认为是土壤微生物在完成分解后“选择性保留”于土壤的结果[37,38,39],是土壤微生物的“自私行为”.作为优先性分解组分,来自植物残体和叶片的有机物质快速分解损失,而来自根部的植物有机质倾向于保留在土壤中[40],使土壤表现出对根有机物的“偏好”(亲合性).Rasse等[41]指出,根系来源的有机化合物实际上是土壤有机质积累和固定的主要来源,他们认为来自根系的有机质比来自地上部的有机质更易被土壤固定而保留于土壤中,这是因为根系活动通过真菌菌根、根毛和分泌物的矿质(金属)结合、团聚体物理保护等机制来促进根系源物质的优先残留保持,而不是由稳定性化学转化而决定的.上述的植物源芳香化合物的可溶性组分被矿质优先吸附和固定也符合选择性残留保持机制. ...
Is soil carbon mostly root carbon? Mechanisms for a specific stabilization
2
2005
... 与腐殖质学说相交存在的另一种观点是,土壤有机质是植物来源有机物的残留物.20世纪30年代,已经有不同分解阶段的植物残留有机物的化学组成资料.到20世纪末,对于土壤中不同植物来源有机分子的存在及其分解性已经有详尽的资料[36].这些未分解、部分分解和正在分解转化中的有机组分构成了瞬时的土壤有机质主体,当然它们处于连续的动态变化中.之前研究认为,真菌促进了新鲜有机物分解而使其残留物成为土壤有机质.例如,植物蛋白质降解产生的氨基酸在土壤中残留而形成了稳定的含氮有机物质.后来,这种有机质残留被认为是土壤微生物在完成分解后“选择性保留”于土壤的结果[37,38,39],是土壤微生物的“自私行为”.作为优先性分解组分,来自植物残体和叶片的有机物质快速分解损失,而来自根部的植物有机质倾向于保留在土壤中[40],使土壤表现出对根有机物的“偏好”(亲合性).Rasse等[41]指出,根系来源的有机化合物实际上是土壤有机质积累和固定的主要来源,他们认为来自根系的有机质比来自地上部的有机质更易被土壤固定而保留于土壤中,这是因为根系活动通过真菌菌根、根毛和分泌物的矿质(金属)结合、团聚体物理保护等机制来促进根系源物质的优先残留保持,而不是由稳定性化学转化而决定的.上述的植物源芳香化合物的可溶性组分被矿质优先吸附和固定也符合选择性残留保持机制. ...
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Refractory organic carbon in particle-size fractions of arable soils I: Distribution of refractory carbon between the size fractions
1
2002
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
What is recalcitrant soil organic matter?
2
2010
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
Contribution of lignin and polysaccharides to the refractory carbon pool in C-depleted arable soils
2
2003
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Labile, recalcitrant, and inert organic matter in Mediterranean forest soils
1
2007
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
Does the acid hydrolysis-incubation method measure meaningful soil organic carbon pools?
1
2006
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
A new conceptual model for the fate of lignin in decomposing plant litter
1
2011
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
Review of mechanisms and quantification of priming effects
2
2000
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Oxidoreductase enzymes liberated by plant roots and their effects on soil humic material
1
1999
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
Decomposition of carbon-14-labeled organic amendments and humic acids in a long-term field experiment
1
2009
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
Stabilization of organic matter in temperate soils: Mechanisms and their relevance under different soil conditions—A review
2
2006
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass-a new perspective: The biology of the Regulatory Gate
1
2008
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
'Mineralization of native soil organic matter is not regulated by the size, activity or composition of the soil microbial biomass — A new perspective' [Soil Biology & Biochemistry 40, 61-73]
1
2009
... 腐殖化理论指向土壤有机质持留存在的终极机制是腐殖物质的分子化学稳定性,但不易被微生物矿化分解,成为抗性(refractory)有机质,特别是有机质损失而亏缺的农业土壤中残余的有机质部分[42].随着有机质的输入,不稳定组分首先被土壤中的微生物利用,而稳定性组分只有在不稳定组分消耗殆尽时才有可能被微生物利用,因而在土壤中更长期保存下来成为老碳[43],即微生物的“喜新厌旧”使土壤有机质总体上更新变慢.即使是这样,在森林土壤中,大分子的木质素化合物可能仍会选择性地保留在大土壤颗粒(数百微米的团聚体)中成为抗性组分,而农业利用下的抗性组分主要在细颗粒(例如粉粒/黏粒级复合体)中,其中选择性地保留了分解有机质产生的微生物多糖[44].腐殖物质化学稳定说曾在土壤学研究中盛极一时,还提出了多种操作方法(如6 mol/L HCl提取组分法)用以表征土壤有机质特别是腐殖物质的化学稳定性[45,46].不过,在合适的条件下,特定的微生物区系对化学惰性明显的“腐殖物质”大分子化合物的分解可能比预想的更快,这看起来就像是土壤微生物遇到一个伴侣时的“兴奋互动”.例如,在植物残体分解的早期,在真菌参与下木质素大分子的分解较快,特别是小分子有机质碳源的存在“激励”微生物矿化了木质素,这在微生物个体可以到达的所有微域都可能发生[47].这种情况其实属于土壤微生物的“激发效应”,微生物喜好的特定基质就像触发剂(trigger molecule),一旦加入就可能导致某些不易分解有机质的共代谢而加快分解[48].类似的,Gramss等[49]将森林植被黑钙土的碱性腐殖质提取物(Humus Extract,HE)暴露于36种真菌和9种细菌的液体培养基液中培养,发现其降解速率惊人,这显然与腐殖质理论中胡敏素生物学稳定性相悖,当然这也属于新食物对土壤微生物的“诱饵效应”.相似的,Tatzber等[50]进行了14C同位素标记有机物在不同种植制土壤的田间分解试验,用1 mol/L NaOH 提取的腐殖质组分的14C同位素丰度都随时间迅速下降,并在所有供试土壤中相似,他们计算出的标记有机物腐殖化形成的腐殖质更新周期在10年左右,即腐殖质分子仍是可以快速分解的,这不能支持腐殖化是有机质的终极稳定过程的认识.在任一时刻,土壤有机质均是不稳定和相对稳定的化学组分同时存在而构成的整体[51].Kemmitt等[52]提出了“调节门假设”(Regulatory Gate hypothesis),认为腐殖质的分解矿化可能包含腐殖物质组分(分子)“出露”(exposure)而可被分解(即暴露于微生物,分解的有效性)以及“捕获”而被微生物食用消化(酶分解)等2个步骤,而出露可能是取决于土壤非生物因素的限速步骤.这个折中的假设,既肯定了土壤腐殖质的化学稳定性,又承认了腐殖质的保护(即不转化为可分解状态)是有机质不被微生物分解的原因.不过,这种观点仍受到土壤生物化学界的强烈反对[53]. ...
SOM fractionation methods: Relevance to functional pools and to stabilization mechanisms
1
2007
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
Old and stable soil organic matter is not necessarily chemically recalcitrant: Implications for modeling concepts and temperature sensitivity
1
2011
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
Similar response of labile and resistant soil organic matter pools to changes in temperature
2
2005
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
Black carbon decomposition and incorporation into soil microbial biomass estimated by 14C labeling
1
2009
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
Soil organic matter turnover is governed by accessibility not recalcitrance
3
2012
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... [58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Advances in understanding the molecular structure of soil organic matter: Implications for interactions in the environment
2
2010
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
Dynamic molecular structure of plant biomass-derived black carbon (biochar)
2
2010
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
How relevant is recalcitrance for the stabilization of organic matter in soils?
1
2010
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
The Nature and Properties of Soils (14th Edition)
3
2008
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... [
62]和(
b)
[63]的演变
<strong>Concept update of soil organic matter and the composition </strong>(<strong>a</strong>)<sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup>和(<strong>b</strong>)<sup>[<xref ref-type="bibr" rid="R63">63</xref>]</sup>Fig.1![]() <strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化
<strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
... [
62]和(
b)
[63]Fig.1![]() <strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化
<strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
The Nature and Properties of Soils (15th Edition)
5
2017
... 21世纪以来,腐殖质组分抗微生物分解的内在(化学)稳定性经典学说越来越被诟病.Kleber[43] 强烈质疑腐殖质学说,认为腐殖质组分抗分解概念是模糊和抽象的,严重误导了土壤碳循环与气候变化的反馈研究,某些组分的固有抗分解性不是其分子性质所决定的,而是受到微生境条件下的生物可达性及/酶反应动力学的制约.例如北美草原土壤农耕后残留的土壤有机碳,更多地稳定于黏粒和细粉砂组分的复合体中,有机碳的14C绝对年龄可能在百年水平[54],他们不是因为腐殖化形成的老碳而残留,而是因为与细颗粒结合,免于分解而保持[55].气候变化研究中经常报道有机质分解的温度敏感性,也支持了是分解条件而不是分解基质的性质决定有机质抗分解性的观点.例如,Fang等[56]实验报道老碳和新碳对升温下微生物分解同样敏感.后来的研究[57]进一步证实14C同位素指示的森林土壤中新碳也可能比老碳中的有机质芳构化更强,而烷基化分子相对更少.Dungait等[58]讨论了现行碳模型的有机质动力学模拟问题,指出已有研究资料并不能支持有机质可分解性可以预测有机质的变化,而复杂分子结构的腐殖质抗微生物分解的化学稳定说不能解释有机质的稳定性.分子检测技术的进步使我们认识到许多土壤中含有大量可分解的有机质.我们不得不认为,探寻化学稳定碳来诠释固碳,可能使土壤固碳机制的探索走向歧途[58].而认识土壤碳不被微生物利用的保护机制既能更好地模拟碳动态,又能用于土壤管理.由此看来,腐殖质学说的化学结构稳定性不但在化学本质上受到质疑,在解释土壤有机质稳定机制上也日益显得苍白而难以为继.基于有机质物理化学本质的腐殖质学说可能误导了对土壤有机质过程以及生态系统碳循环的理解,需要新的角度和技术方法重新构建土壤有机质理论[59,60,61].这些认识进展已经反映在国外权威教科书《The Nature and Properties of Soils》第15版对其第14版中关于土壤有机质概念及其组成结构的修正上(图1)[62,63]. ...
... [
63]的演变
<strong>Concept update of soil organic matter and the composition </strong>(<strong>a</strong>)<sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup>和(<strong>b</strong>)<sup>[<xref ref-type="bibr" rid="R63">63</xref>]</sup>Fig.1![]() <strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化
<strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
... [
63]
Fig.1![]() <strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化
<strong>2.3</strong> 有机碳结合保护学说诠释土壤有机质稳定化20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
... 在生态系统中,无论来自地上部还是地下部生物生命活动留下的残余物或者代谢产物,在土壤中随时随地处于不同程度微生物接触而降解的过程,因此瞬时的土壤有机质应该是所有曾经及正在土壤中活动的生命代谢产物或有机体残余,仅是不同程度微生物降解的化合物的集合,构成了土壤有机质系统.不同土壤所处生态系统演替过程各异,作为有机质输入来源的植物种类及其比例差异在不同生态系统土壤中差异很大,欧洲学界将此抽象为腐殖质系统及其形态类型(humus system and forms) (这里的腐殖质指土壤中呈暗色的稳定有机质[63]在土壤景观中的不同生态关系,而不是基于化学提取的传统腐殖质概念)[134].故而任何一种土壤中,植物天然生物标志物分子丰度及组成都可能反映特定系统的土壤有机质特点,更可以用来追溯有机质来源或者生态系统演化历程.除了前述的植物生物标志物外,养殖动物粪便有机质中的生物标志物系列也已建立,采用高温气质联用检测可以检测到施用奶牛粪便土壤中的动物生物标志物5β-stanols (n~ n ×102 ng/g),从而可以追踪养殖废弃物对土壤的有机污染[135].当然,土壤中还可能存在其他来源有机物质,如农药、抗生素和农膜等有机化合物污染,这些分子可以采用气相色谱法直接检测出来.早在20世纪80年代初,就有学者报道土壤中人为源化合物(污染化合物,xenobiotics)多达30多种,其在土壤有机质中可能高达10%~30%[136]. ...
Core issues and research progresses of soil science of C sequestration
1
2007
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
固碳土壤学的核心科学问题与研究进展
1
2007
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
A profile of international climate change science in the past one hundred years
1
2018
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
国际气候变化科学百年研究态势分析
1
2018
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
Stabilization of soil organic matter: Association with minerals or chemical recalcitrance?
2
2006
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Soil macroaggregate turnover and microaggregate formation: A mechanism for C sequestration under no-tillage agriculture
2
2000
... 20世纪末以来,减缓气候变化的国际需求推动了农业固碳减排研究及其技术发展,这反过来拉动了土壤有机质稳定和积累机制的研究,成为一个时期土壤学研究的前沿和热点领域[64,65].经典土壤腐殖质理论的核心是芳构化大分子聚合成为化学稳定性的复杂有机多聚体,形成土壤中稳定性有机质碳库.相反,十多年来的土壤固碳与生态系统功能的研究,越来越倾向于有机质的矿物—团聚体保护/稳定机制,而不是其复杂的化学转化,来诠释土壤有机质的固定和稳定[51,66,67].基于强电解质提取的腐殖质学说,其实只是肯定了复杂有机质与土壤中组分结合稳定的存在状态. ...
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
A model of the physical protection of organic matter in soils
1
1997
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
Amount and composition of clay-associated soil organic matter in a range of kaolinitic and smectitic soils
1
2001
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
Clay mineral composition modifies decomposition and sequestration of organic carbon and nitrogen in fine soil fractions
1
2015
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
Carbon losses from all soils across England and Wales 1978-2003
1
2005
... 早期的黏粒保护理论认为,有机物是被黏粒矿物吸附到表面从而表现为稳定碳的存在[68],这其实也是腐殖质学说的强电解质提取方法的依据:土壤有机物质是与反应性矿物结合的复杂集合体.不同萃取条件对黏粒有机复合体破坏程度不同,提取出的土壤胡敏酸的结构特性也不同.使用传统碱提取剂提取的胡敏酸含有的脂肪族化合物和苯酚类物质比树脂法提取出的胡敏酸多[24],似乎是与黏粒矿物质结合越紧密,有机质分子芳构化越高,分子量越大,实际上可能是强电解质下有机物的“假象”缩合,而黏粒中结合的有机分子可能是亲水性小的有机组分.采自不同国家的热带生态系统土壤的分析表明,黏粒矿物结合态有机质的多少与矿物类型无关,而高岭石矿物趋向于结合富集多糖类有机质分子,2∶1的蒙皂石则趋向于结合较多的芳香族有机分子[69],其可能是参与分解有机质的微生物生物体及其代谢产物残留[70].反之,不与矿质紧密结合的颗粒态碳,在土壤中呈较为活泼的游离态有机质,也并不一定是分解程度低或者生物活性低的年轻碳.Six等[67]对美国不同生态区农业土壤常规耕作和免耕体系中土壤有机质分布进行了团聚体分组研究,免耕土壤中表现为大团聚体内微团聚体间或其中的颗粒态有机质的积累,13C同位素丰度分析也揭示了这些颗粒态碳可能是非微生物来源的老碳,它们在土壤中的稳定受矿质—有机质复合过程影响的团聚体保护.这说明,土壤中有机质的分解不是有机碳组分的化学稳定性变化的问题,而是土壤中失去保护的问题.例如,在微生物有机质分解实验中,老碳可能比新碳对升温更敏感[56];又如,处于北温带的英国,土壤有机质碳在升温加快的1990—2000年间快速损失[71]. ...
Organic Matter
1
... 同理,可溶性有机质也不一定是结构简单而易分解的新碳,还可能是老碳.在土壤腐殖质理论中,可溶性组分是最活泼的有机组分,常被认为是有机质初级分解中释放的简单化合物,例如纤维素等植物源有机物质分解的有机酸[72].采用分组结合13C核磁共振法对同是火山岩发育的不同年代的夏威夷土壤有机质的研究[73,74],揭示出最持久性的矿质结合碳成分是部分氧化的植物源芳香化合物,这些固定的稳定有机质来自含有植物源芳香烃化合物的可溶性有机质,其通过矿物表面的选择性吸附而被矿质结合固定.这种认识后来进一步被纳米颗粒同步辐射(Nano-SIMS)微域研究所证实[75].采用Nano-SIMS结合微域同位素探针,揭示了培养条件下玉米秸秆初级分解产物优先结合于有机矿质复合的团聚体表面,而不是被单个黏粒矿物表面所结合.他们认为,新碳的固定和稳定化可能并不是化学转化的稳定性有机物所致,而是植物源有机质被团聚体结合稳定之故. ...
Biogeochemistry of mineral-organic associations across a long-term mineralogical soil gradient (0.3-4100 kyr), Hawaiian Islands
1
2009
... 同理,可溶性有机质也不一定是结构简单而易分解的新碳,还可能是老碳.在土壤腐殖质理论中,可溶性组分是最活泼的有机组分,常被认为是有机质初级分解中释放的简单化合物,例如纤维素等植物源有机物质分解的有机酸[72].采用分组结合13C核磁共振法对同是火山岩发育的不同年代的夏威夷土壤有机质的研究[73,74],揭示出最持久性的矿质结合碳成分是部分氧化的植物源芳香化合物,这些固定的稳定有机质来自含有植物源芳香烃化合物的可溶性有机质,其通过矿物表面的选择性吸附而被矿质结合固定.这种认识后来进一步被纳米颗粒同步辐射(Nano-SIMS)微域研究所证实[75].采用Nano-SIMS结合微域同位素探针,揭示了培养条件下玉米秸秆初级分解产物优先结合于有机矿质复合的团聚体表面,而不是被单个黏粒矿物表面所结合.他们认为,新碳的固定和稳定化可能并不是化学转化的稳定性有机物所致,而是植物源有机质被团聚体结合稳定之故. ...
Controls on carbon storage and weathering in volcanic soils across a high-elevation climate gradient on Mauna Kea, Hawaii
1
2016
... 同理,可溶性有机质也不一定是结构简单而易分解的新碳,还可能是老碳.在土壤腐殖质理论中,可溶性组分是最活泼的有机组分,常被认为是有机质初级分解中释放的简单化合物,例如纤维素等植物源有机物质分解的有机酸[72].采用分组结合13C核磁共振法对同是火山岩发育的不同年代的夏威夷土壤有机质的研究[73,74],揭示出最持久性的矿质结合碳成分是部分氧化的植物源芳香化合物,这些固定的稳定有机质来自含有植物源芳香烃化合物的可溶性有机质,其通过矿物表面的选择性吸附而被矿质结合固定.这种认识后来进一步被纳米颗粒同步辐射(Nano-SIMS)微域研究所证实[75].采用Nano-SIMS结合微域同位素探针,揭示了培养条件下玉米秸秆初级分解产物优先结合于有机矿质复合的团聚体表面,而不是被单个黏粒矿物表面所结合.他们认为,新碳的固定和稳定化可能并不是化学转化的稳定性有机物所致,而是植物源有机质被团聚体结合稳定之故. ...
Submicron structures provide preferential spots for carbon and nitrogen sequestration in soils
1
2014
... 同理,可溶性有机质也不一定是结构简单而易分解的新碳,还可能是老碳.在土壤腐殖质理论中,可溶性组分是最活泼的有机组分,常被认为是有机质初级分解中释放的简单化合物,例如纤维素等植物源有机物质分解的有机酸[72].采用分组结合13C核磁共振法对同是火山岩发育的不同年代的夏威夷土壤有机质的研究[73,74],揭示出最持久性的矿质结合碳成分是部分氧化的植物源芳香化合物,这些固定的稳定有机质来自含有植物源芳香烃化合物的可溶性有机质,其通过矿物表面的选择性吸附而被矿质结合固定.这种认识后来进一步被纳米颗粒同步辐射(Nano-SIMS)微域研究所证实[75].采用Nano-SIMS结合微域同位素探针,揭示了培养条件下玉米秸秆初级分解产物优先结合于有机矿质复合的团聚体表面,而不是被单个黏粒矿物表面所结合.他们认为,新碳的固定和稳定化可能并不是化学转化的稳定性有机物所致,而是植物源有机质被团聚体结合稳定之故. ...
Pyrogenic carbon in soils: A literature-based inventory and a global estimation of its content in soil organic carbon and stocks
1
2016
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
Terrestrial pyrogenic carbon export to fluvial ecosystems: Lessons learned from the White Nile watershed of East Africa
1
2016
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
Industrialization of biochar from biomass pyrolysis: A new option for straw burning ban and green agriculture of China
1
2015
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
热裂解生物质炭产业化: 秸秆禁烧与绿色农业新途径
1
2015
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
Anaerobic microsites have an unaccounted role in soil carbon stabilization
1
2017
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
Persistence of soil organic matter as an ecosystem property
1
2011
... 此外,热解残留碳(pyrogenic carbon),常被称为炭黑或黑碳(black carbon),曾被认为是土壤有机质中最持久的碳组分[76],在某些发生过自然大火的森林土壤有机碳中其比例可能高达60%,其中也检测出多糖等简单的单个有机组分[60,77],这与前述农业土壤中分解残留的抗性碳中有微生物多糖残留不谋而合.很多研究表明,生物质炭(biochar)这样的颗粒态有机碳,实际上是多种复杂生物源有机物质的总称,而不是一类结构复杂和稳定的有机化合物[78],被归类于新定义的腐殖质的一种类型[63].最近,Keiluweit等[79]采用氧饱和溶液扩散装置研究了与不同细度石英砂混合的土壤有机质的矿化,表明氧气的进入程度控制着土壤有机质矿化速率.因而提出,即使在好气土壤中,微细土壤颗粒的有机质矿化因氧气难以进入而受阻,使脂肪酸和糖类等不被分解而存留.年轻(活性)有机质得到较长时间滞留(更新速率降低)的土壤机制是由厌氧微生境保护引起的,而与基质的易矿化性无关,它属于持留机制(persistence)而不是化学上的固定机制(bound or sequestered).这提示土壤固碳更多地属于生物物理过程(biophysical),而不是化学形态的转化或分子稳定化过程.这与先前关于土壤固碳是还原态碳积累的研究[59]异曲同工.前述的复杂有机质组分有条件的抗分解性,实质上也是土壤(团聚体)中矿物、孔隙和微生物复杂组合结构对这些不同有机质分子组分的特定(选择性)保护,土壤物理化学保护机制在土壤有机质稳定性中的作用比过去所认为的更重要[80].因此,探析土壤有机质本质需要更多地基于生命分子鉴定的分子土壤有机质认识和检测,特别是分析分子组成与存在特点. ...
Basis of a humeomics science: Chemical fractionation and molecular characterization of humic biosuprastructures
4
2011
... 应用检测技术进行物质结构分析的前提是待测物质中组分组成尽量富集或单一纯净.“腐殖质”成分的复杂性并不能满足各组分的分子结构鉴定.为细致地研究腐殖质各组分的分子组成,模拟腐殖质由单一的简单组分与土壤物质形成不同结合力的复杂组分,采用模拟结合力的有机提取剂分离特定的腐殖质组分一直是腐殖质学说坚守者的努力方向.2011年Piccolo团队首次提出了“腐殖质组学(Humeomics)”概念及其方法框架[81].腐殖质组学是基于有机溶剂萃取的连续性分离法,通过与三氟化硼在甲醇体系中的酯交换、甲醇的碱性水解、使用HI裂解醚和糖苷键合等一系列反应,土壤中不同结合状态的有机分子被连续提取和释放,通过气相色谱鉴定反应释放出的腐殖质组分分子,最后表征土壤腐殖质的分子组成结构和特点(图2)[82].这种腐殖质组学法分离得到的有机组分可以偶联于核磁共振波谱分析、气质联用分析和电喷雾静电场轨道阱组合式高分辨质谱联用分析系统(ESI-Orbitrap-MS),从而获得高分辨率的腐殖质分子组成信息[83]. ...
... 土壤腐殖质组学的连续提取分离法程序框架(据参考文献[
81]修改)
<strong>Procedure framework of sequential extraction and separation of soil humus</strong>(<strong>modified after reference </strong>[<xref ref-type="bibr" rid="R81">81</xref>])Fig.2![]()
这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
... [
81])
Fig.2![]()
这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
... 这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
A molecular zoom into soil Humeome by a direct sequential chemical fractionation of soil
2
2017
... 应用检测技术进行物质结构分析的前提是待测物质中组分组成尽量富集或单一纯净.“腐殖质”成分的复杂性并不能满足各组分的分子结构鉴定.为细致地研究腐殖质各组分的分子组成,模拟腐殖质由单一的简单组分与土壤物质形成不同结合力的复杂组分,采用模拟结合力的有机提取剂分离特定的腐殖质组分一直是腐殖质学说坚守者的努力方向.2011年Piccolo团队首次提出了“腐殖质组学(Humeomics)”概念及其方法框架[81].腐殖质组学是基于有机溶剂萃取的连续性分离法,通过与三氟化硼在甲醇体系中的酯交换、甲醇的碱性水解、使用HI裂解醚和糖苷键合等一系列反应,土壤中不同结合状态的有机分子被连续提取和释放,通过气相色谱鉴定反应释放出的腐殖质组分分子,最后表征土壤腐殖质的分子组成结构和特点(图2)[82].这种腐殖质组学法分离得到的有机组分可以偶联于核磁共振波谱分析、气质联用分析和电喷雾静电场轨道阱组合式高分辨质谱联用分析系统(ESI-Orbitrap-MS),从而获得高分辨率的腐殖质分子组成信息[83]. ...
... 这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
Humeomics: A key to unravel the humusic pentagram
3
2018
... 应用检测技术进行物质结构分析的前提是待测物质中组分组成尽量富集或单一纯净.“腐殖质”成分的复杂性并不能满足各组分的分子结构鉴定.为细致地研究腐殖质各组分的分子组成,模拟腐殖质由单一的简单组分与土壤物质形成不同结合力的复杂组分,采用模拟结合力的有机提取剂分离特定的腐殖质组分一直是腐殖质学说坚守者的努力方向.2011年Piccolo团队首次提出了“腐殖质组学(Humeomics)”概念及其方法框架[81].腐殖质组学是基于有机溶剂萃取的连续性分离法,通过与三氟化硼在甲醇体系中的酯交换、甲醇的碱性水解、使用HI裂解醚和糖苷键合等一系列反应,土壤中不同结合状态的有机分子被连续提取和释放,通过气相色谱鉴定反应释放出的腐殖质组分分子,最后表征土壤腐殖质的分子组成结构和特点(图2)[82].这种腐殖质组学法分离得到的有机组分可以偶联于核磁共振波谱分析、气质联用分析和电喷雾静电场轨道阱组合式高分辨质谱联用分析系统(ESI-Orbitrap-MS),从而获得高分辨率的腐殖质分子组成信息[83]. ...
... 这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
... 腐殖质组学的可贵之处是表征了“腐殖质”有机物组分间的化学连接方式以及不同结合力所保持组分的比例(相当于微生物community structure).采用这种方法分析表明,腐殖质分子既可以以疏水键合黏附于黏土矿物表面,也可以通过共价键合结合于无定形有机铁化合物,从而成为稳定的碳组分.腐殖质组学还认为,腐殖质中脂肪族有机分子可能主要来自植物酯类[86],而不是细菌代谢产物,当然后者正是当前研究热点,多有争论[87,88].正因如此,腐殖质组学方法原理上有可能说明土壤有机质的结构/功能关系,例如组分分布与碳固定的关系以及组分比率变化与土壤中生物活性及反应性的关系[83,89]. ...
Unveiling the molecular composition of the unextractable soil organic fraction (humin) by humeomics
1
2015
... 这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
The molecular properties of biochar carbon released in dilute acidic solution and its effects on maize seed germination
1
2017
... 这套方法体系在传统的腐殖质碱提取法基础上,试图将所有土壤有机质组分能连续地分离纯化,继而通过有机组分的高分辨率分子鉴定来详细解析各分离组分的腐殖质分子组成,以获得比传统碱提取技术更多的分子组成信息.然而,基于土壤胡敏酸的腐殖质组学组分所获得的分子组成,并不能表征全部土壤有机质分子组成信息.为此,Drosos等[82,83]提出了修正的提取分离方法程序(图2),代替上述方法中对胡敏酸的提取,第一步首先对土壤进行0.1 mol/L HCl溶解提取,并采用传统碱溶液提取法对上述组学法第四步提取后的残留物(RES 4)再次提取,获得残留有机组分(residual organic matter,RESOM)及最终残渣(total residue,REST),其他提取剂及流程与先前的方法保持不变[81].改进后的腐殖质组学技术能够直接应用于原始土壤样品,可最大程度地提取和分离出更多的土壤有机质组分,获得令人满意的土壤有机质总回收率[84].不但获得了第一个游离态水溶性(其实是弱酸溶性)组分(AQU1),并对残留组分进行进一步分离,获得了更多的土壤有机质组分分子组成信息,继而通过不同组分的质量组成及其比例[类似于微生物基因的丰度、结构及多样性,如对不同组分进行van Krevelen作图(图3)],可以探究土壤中不同组分对总有机质的贡献[85]. ...
Advances in humeomics: Enhanced structural identification of humic molecules after size fractionation of a soil humic acid
1
2012
... 腐殖质组学的可贵之处是表征了“腐殖质”有机物组分间的化学连接方式以及不同结合力所保持组分的比例(相当于微生物community structure).采用这种方法分析表明,腐殖质分子既可以以疏水键合黏附于黏土矿物表面,也可以通过共价键合结合于无定形有机铁化合物,从而成为稳定的碳组分.腐殖质组学还认为,腐殖质中脂肪族有机分子可能主要来自植物酯类[86],而不是细菌代谢产物,当然后者正是当前研究热点,多有争论[87,88].正因如此,腐殖质组学方法原理上有可能说明土壤有机质的结构/功能关系,例如组分分布与碳固定的关系以及组分比率变化与土壤中生物活性及反应性的关系[83,89]. ...
SOM genesis: Microbial biomass as a significant source
1
2012
... 腐殖质组学的可贵之处是表征了“腐殖质”有机物组分间的化学连接方式以及不同结合力所保持组分的比例(相当于微生物community structure).采用这种方法分析表明,腐殖质分子既可以以疏水键合黏附于黏土矿物表面,也可以通过共价键合结合于无定形有机铁化合物,从而成为稳定的碳组分.腐殖质组学还认为,腐殖质中脂肪族有机分子可能主要来自植物酯类[86],而不是细菌代谢产物,当然后者正是当前研究热点,多有争论[87,88].正因如此,腐殖质组学方法原理上有可能说明土壤有机质的结构/功能关系,例如组分分布与碳固定的关系以及组分比率变化与土壤中生物活性及反应性的关系[83,89]. ...
Direct evidence for microbial-derived soil organic matter formation and its ecophysiological controls
1
2016
... 腐殖质组学的可贵之处是表征了“腐殖质”有机物组分间的化学连接方式以及不同结合力所保持组分的比例(相当于微生物community structure).采用这种方法分析表明,腐殖质分子既可以以疏水键合黏附于黏土矿物表面,也可以通过共价键合结合于无定形有机铁化合物,从而成为稳定的碳组分.腐殖质组学还认为,腐殖质中脂肪族有机分子可能主要来自植物酯类[86],而不是细菌代谢产物,当然后者正是当前研究热点,多有争论[87,88].正因如此,腐殖质组学方法原理上有可能说明土壤有机质的结构/功能关系,例如组分分布与碳固定的关系以及组分比率变化与土壤中生物活性及反应性的关系[83,89]. ...
Linking organic matter chemistry with soil aggregate stability: Insight from 13C NMR spectroscopy
1
2018
... 腐殖质组学的可贵之处是表征了“腐殖质”有机物组分间的化学连接方式以及不同结合力所保持组分的比例(相当于微生物community structure).采用这种方法分析表明,腐殖质分子既可以以疏水键合黏附于黏土矿物表面,也可以通过共价键合结合于无定形有机铁化合物,从而成为稳定的碳组分.腐殖质组学还认为,腐殖质中脂肪族有机分子可能主要来自植物酯类[86],而不是细菌代谢产物,当然后者正是当前研究热点,多有争论[87,88].正因如此,腐殖质组学方法原理上有可能说明土壤有机质的结构/功能关系,例如组分分布与碳固定的关系以及组分比率变化与土壤中生物活性及反应性的关系[83,89]. ...
Air Pollutants and the Leaf Cuticle
2
1994
... 直至20世纪末,生命特征化合物(分子)鉴定技术不断成熟,并逐渐用以探究土壤有机质的本质[90].了解有机质分子分布,需要辨析生命来源的有机物随着分解的分子变化.在正常土壤中,土壤有机质来源于植物残体、根系及其分泌物,以及土壤微生物生物体及其代谢产物.这些是不同分子大小和碳结构的多糖、单宁、蜡质、脂类、木质素和蛋白质等类群的有机物质.土壤中存在的这些有机物质可能因来源和降解程度的差异在分子结构及碳链长度上带有特定生命信息,称为有机质的分子生物标志物(biomarker molecules)[91,92].鉴定这些分子的丰度和组成,可能为探索土壤有机质生命来源、追溯生态系统演变提供了一种新的技术途径[91,92,93,94]. ...
... 注:据参考文献[90,92,97]等综合 ...
Occurrence of biomarkers and straight-chain biopolymers in humin: Implication for the origin of soil organic matter
2
1998
... 直至20世纪末,生命特征化合物(分子)鉴定技术不断成熟,并逐渐用以探究土壤有机质的本质[90].了解有机质分子分布,需要辨析生命来源的有机物随着分解的分子变化.在正常土壤中,土壤有机质来源于植物残体、根系及其分泌物,以及土壤微生物生物体及其代谢产物.这些是不同分子大小和碳结构的多糖、单宁、蜡质、脂类、木质素和蛋白质等类群的有机物质.土壤中存在的这些有机物质可能因来源和降解程度的差异在分子结构及碳链长度上带有特定生命信息,称为有机质的分子生物标志物(biomarker molecules)[91,92].鉴定这些分子的丰度和组成,可能为探索土壤有机质生命来源、追溯生态系统演变提供了一种新的技术途径[91,92,93,94]. ...
... [91,92,93,94]. ...
What happens to terrestrial organic matter in the ocean?
5
1997
... 直至20世纪末,生命特征化合物(分子)鉴定技术不断成熟,并逐渐用以探究土壤有机质的本质[90].了解有机质分子分布,需要辨析生命来源的有机物随着分解的分子变化.在正常土壤中,土壤有机质来源于植物残体、根系及其分泌物,以及土壤微生物生物体及其代谢产物.这些是不同分子大小和碳结构的多糖、单宁、蜡质、脂类、木质素和蛋白质等类群的有机物质.土壤中存在的这些有机物质可能因来源和降解程度的差异在分子结构及碳链长度上带有特定生命信息,称为有机质的分子生物标志物(biomarker molecules)[91,92].鉴定这些分子的丰度和组成,可能为探索土壤有机质生命来源、追溯生态系统演变提供了一种新的技术途径[91,92,93,94]. ...
... ,92,93,94]. ...
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... [92]. ...
... 注:据参考文献[90,92,97]等综合 ...
Combining biomarker with stable isotope analyses for assessing the transformation and turnover of soil organic matter
1
2008
... 直至20世纪末,生命特征化合物(分子)鉴定技术不断成熟,并逐渐用以探究土壤有机质的本质[90].了解有机质分子分布,需要辨析生命来源的有机物随着分解的分子变化.在正常土壤中,土壤有机质来源于植物残体、根系及其分泌物,以及土壤微生物生物体及其代谢产物.这些是不同分子大小和碳结构的多糖、单宁、蜡质、脂类、木质素和蛋白质等类群的有机物质.土壤中存在的这些有机物质可能因来源和降解程度的差异在分子结构及碳链长度上带有特定生命信息,称为有机质的分子生物标志物(biomarker molecules)[91,92].鉴定这些分子的丰度和组成,可能为探索土壤有机质生命来源、追溯生态系统演变提供了一种新的技术途径[91,92,93,94]. ...
Potential applications of cutin-derived CuO reaction products for discriminating vascular plant sources in natural environments
3
1990
... 直至20世纪末,生命特征化合物(分子)鉴定技术不断成熟,并逐渐用以探究土壤有机质的本质[90].了解有机质分子分布,需要辨析生命来源的有机物随着分解的分子变化.在正常土壤中,土壤有机质来源于植物残体、根系及其分泌物,以及土壤微生物生物体及其代谢产物.这些是不同分子大小和碳结构的多糖、单宁、蜡质、脂类、木质素和蛋白质等类群的有机物质.土壤中存在的这些有机物质可能因来源和降解程度的差异在分子结构及碳链长度上带有特定生命信息,称为有机质的分子生物标志物(biomarker molecules)[91,92].鉴定这些分子的丰度和组成,可能为探索土壤有机质生命来源、追溯生态系统演变提供了一种新的技术途径[91,92,93,94]. ...
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... [94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
A review of some important families of refractory macromolecules: Composition, origin, and fate in soils and sediments
1
2001
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
A comparison of plant and microbial biomarkers in grassland soils from the Prairie Ecozone of Canada
2
2005
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... 通过近10多年来的发展,湿化学法结合气相色谱—质谱联用仪法,即土壤中植物标志物提取—萃取—浓缩法及GC/MS定性/定量鉴定法联合技术已经基本完善,可满足土壤有机质生命来源分子的追踪[96,97].Masoom等[132]提出了综合多态核磁共振分析法来探讨天然土壤有机质的分子存在状态,这种分析揭示了土壤有机质基本上是生命代谢或降解过程留下的有机大分子,土水界面主要是亲水的脂类和碳水化合物,而土壤(颗粒)内部是疏水的木质素和微生物代谢产物,后者又可以吸附和固定复杂的疏水有机分子.这也为理解疏水有机分子的固定是土壤固碳机制之一提供了帮助[133],揭示了生命来源的有机分子在土壤粒子空间的连续分布谱系,成为理解土壤有机质整体的新视角. ...
Sources and composition of hydrolysable aliphatic lipids and phenols in soils from western Canada
5
2006
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... [97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
... 注:据参考文献[90,92,97]等综合 ...
... 通过近10多年来的发展,湿化学法结合气相色谱—质谱联用仪法,即土壤中植物标志物提取—萃取—浓缩法及GC/MS定性/定量鉴定法联合技术已经基本完善,可满足土壤有机质生命来源分子的追踪[96,97].Masoom等[132]提出了综合多态核磁共振分析法来探讨天然土壤有机质的分子存在状态,这种分析揭示了土壤有机质基本上是生命代谢或降解过程留下的有机大分子,土水界面主要是亲水的脂类和碳水化合物,而土壤(颗粒)内部是疏水的木质素和微生物代谢产物,后者又可以吸附和固定复杂的疏水有机分子.这也为理解疏水有机分子的固定是土壤固碳机制之一提供了帮助[133],揭示了生命来源的有机分子在土壤粒子空间的连续分布谱系,成为理解土壤有机质整体的新视角. ...
Fate of lignins in soils: A review
1
2010
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
The lignin component of humic substances: Distribution among soil and sedimentary humic, fulvic, and base-insoluble fractions
1
1984
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
A reassessment of the sources and importance of land-derived organic matter in surface sediments from the Gulf of Mexico
2
1998
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
Biomarker signature in tropical wetland: Lignin Phenol Vegetation Index (LPVI) and its implications for reconstructing the paleoenvironment
3
2004
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... ,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
Sensing the ups and downs of Las Vegas: InSAR reveals structural control of land subsidence and aquifer-system deformation
2
1999
... 植物来源的脂类物质和木质素可稳定存在于土壤中,角质(cutins)和木栓质(suberins)中不同碳链长度分子的组成或特定分子的比例常用来研究土壤中有机质的植物来源及其降解程度[94,95,96,97].作为导管植物的生物大分子,木质素降解的特征单体包括香兰基(Vanillyls, 代号V)、丁香基(Syringyls, 代号S)和肉桂基(Cinnamyls,代号C),常用作木本生物标志物[92].丁香基/香兰基值(S/V)和肉桂基/香兰基值(C/ V)常用于推断土壤中木质素来源的植被类型和量化不同植被来源的有机质对土壤碳库的贡献[94,98,99,100],因而可指示土地利用变化导致的有机质组成变化.与上述简单的比率不同,Tarep等[101]提出了木质素酚类植被指数(Lignin Phenol Vegetation Index,LPVI)综合算式,来衡量3种木质素单体组分的相对贡献.LPVI值在被子植物和裸子植物的木本组织、非木本组织的木质素间没有交叠,可更好地分辨出土壤有机质中组成复杂的混合物的植物来源[97,101].木质素单体的酚酸分子与其各自对应的醛分子的比值(Ad/Al),会随木质素分解而升高,故也可以用于作为分析土壤中木质素分解程度的替代指标[102].但是,这些比值在森林土壤有机质中的范围较耕地和草地宽,间接地提示了森林土壤有机质起源的多样性和复杂性以及土壤有机质具有连续的分解程度[92]. ...
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
Strengthening the soil organic carbon pool by increasing contributions from recalcitrant aliphatic bio (macro) molecules
1
2007
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
Chemistry, biochemistry, and function of suberin and associated waxes
1
1989
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
Cutin and suberin in temperate forest soils
1
1993
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
The poly (phenolic) domain of potato suberin: A non-lignin cell wall bio-polymer
3
2001
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
... [106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Suberin research in the genomics era—New interest for an old polymer
2
2011
... 角质素化合物是非晶态的大分子生物共聚酯,其成分主要是羧酸(n-alkanoic acids)、中链羟基烃酸(midchain ω-hydroxy-alkanoic acids)和C16单/双羟基酸,并含有一定数量以酯键结合的环氧基团[103].木栓质化合物则主要来自于植物的根系和树皮,包括脂肪族聚酯、多酚类化合物以及长链羟基烃酸(链长度≥C20)[104,105,106].角质/木栓质替代脂肪酸(suberin/cutin-derived substituted fatty acids, Biomarkers-SFA)是相对稳定的植物源生物标志物[101],在C3与C4植物下,土壤有机质中20~30个碳长度的中长链羧酸(如n-C22,n-C24和n-C26)分布模式迥异[102],因而可以用来探讨植物地下部/地上部的有机质来源[107].角质素类和木栓质类化合物的组成成分中含有双键或多个羟基的羟基酸,都比ω-羟基烃酸易分解.因此,ω-C16/ΣC16(ω-C16是16-羟基十六酸的丰度,而ΣC16是16-羟基十六酸、十六碳二酸、7或8-羟基十六碳二酸和10,16-二羟基十六酸等分子的丰度之和)和ω-C18 /ΣC18(ω-C18是18-羟基十八酸丰度,而ΣC18是18-羟基十八酸、十八碳二酸、7或8-羟基十八碳二酸、10,18-二羟基十八酸等分子的丰度和)可作为指示角质素类物质和木栓质类物质降解程度的指标.ω-C16 /ΣC16值和ω-C18 /ΣC18值都会随角质素或木栓质类物的降解而增大[100],在土壤剖面中也随深度而递增[97],故可用来指示有机质生物的稳定性. ...
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Source and turnover of organic matter in agricultural soils derived from n-alkane/n-carboxylic acid compositions and C-isotope signatures
1
2004
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Lignin phenols and cutin-and suberin-derived aliphatic monomers as biomarkers for stand history, SOM source, and turnover
2
2010
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
... ,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Apoplastic polyesters in Arabidopsis surface tissues—A typical suberin and a particular cutin
2
2005
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
... [110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Suberin: A promising renewable resource for novel macromolecular materials
1
2006
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Can cutin and suberin biomarkers be used to trace shoot and root-derived organic matter?A molecular and isotopic approach
2
2011
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
... [112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Molecular dynamics of shoot vs. root biomarkers in an agricultural soil estimated by natural abundance 13C labelling
2
2010
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
... [113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Distribution and decomposition pattern of cutin and suberin in forest soils
1
1989
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Wax-deficient anther1 is involved in cuticle and wax production in rice anther walls and is required for pollen development
1
2006
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Root-derived short-chain suberin diacids from rice and rape seed in a paddy soil under rice cultivar treatments
2
2015
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
... [116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
113(G3)
1
2008
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Sources of plant‐derived carbon and stability of organic matter in soil: Implications for global change
1
2009
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Increased coniferous needle inputs accelerate decomposition of soil carbon in an old-growth forest
1
2009
... 一般认为,木栓质中的脂肪二酸(di-acids)来自单子叶植物根系[108,109],常作为农作物植物根系生物标志物(root biomarker)[106].而角质,特别是其中的链羟基羧酸(mid-chain hydroxy carboxylic acids)来自地上部茎叶组织,常作为地上部生物标志物(shoot biomarker)[107].作为土壤中稳定的植物源有机化合物[106],检测这些替代性脂肪酸的相对丰度,可以区分根和叶片对土壤有机质的输入和稳定,并评估植物地上和地下部对土壤有机质的贡献[41,109,110,111,112,113,114].例如,拟南芥(Arabidopsis)根系中脂肪二酸由C16,C18,C18(1),C20,C22和C24组成[110],但水稻根系脂肪二酸则主要由C16和C18构成[115],在不同品种种植下它们的相对丰度会产生差异[116];相对于森林土壤具有丰富的木栓质来源的有机质,角质二羧酸脂肪酸提供了鉴定和分析农田土壤有机质分子的途径[112].Mendez-Millan等[113]提出了采用气质联用技术鉴定禾本科C3/C4作物几丁质和木栓质脂肪族化合物的方法,可以定量区分地上部和地下部、玉米和小麦输入有机质的贡献.在水稻土,通过选择性提取脂肪酸分子,也可以定性鉴别水稻不同品种根系输入的土壤有机质的贡献[116].土壤有机质生物标志物分子组成变化有助于了解土地利用变化下有机质变化的本质[117],并帮助认识应对气候变化的土壤固碳的生物机制[118,119].目前为止土壤有机质分子组成研究中报道的生物标志物及其鉴定综合展示于表1. ...
Temperature sensitivity of black carbon decomposition and oxidation
1
2010
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
SOC enhancement in three major types of paddy soils in a long term agro-ecosystem experiment in South China. III, Structural variation of two paddy soils of particular organic matter of two paddy soils
2
2009
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
三种南方典型水稻土长期试验下有机碳积累机制研究Ⅲ.两种水稻土颗粒有机质结构特征的变化
2
2009
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Water extract from straw biochar used for plant growth promotion: An initial test
2
2016
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Approaches and limits for accurate Mass characterization of large biomolecules
1
2008
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Germination tests for assessing biochar quality
1
2012
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Characterization of pyrogenic black carbon by desorption atmospheric pressure photoionization Fourier transform ion cyclotron resonance mass spectrometry
1
2012
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Longitudinal shifts in dissolved organic matter chemogeography and chemodiversity within headwater streams: A river continuum reprise
2
2015
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
... [126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
HRMAS NMR spectroscopy applications in agriculture
1
2017
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Pyrolysis temperature-dependent release of dissolved organic carbon from plant, manure, and biorefinery wastes
1
2013
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Characterization of biochar-derived dissolved organic matter using UV-visible absorption and excitation-emission fluorescence spectroscopies
1
2014
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Persistence of soil organic matter in eroding versus depositional landform positions
1
2012
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
OMDY: A new model of organic matter decomposition based on biomolecular content as assessed by 13C-CPMAS-NMR
1
2016
... 以往分子有机质研究多是采用固态红外光谱或者固态核磁共振波谱直接对土壤样品进行有机质官能团组成分析[120].而对颗粒态有机质[121]或者溶解性有机质[122]的分子分析需进行提取制样,结合气质联用分析可以达到定量表征的要求;近年来,基于质谱—气质联用技术的静电场轨道阱组合式质谱分析(electrospray ionization-Orbitrap-mass spectrometry,ESI-MS ESI-Orbitrap-MS)[123]、热解吸气质联用(thermal desorption system gas chromatography-mass spectrometry, TDS-GC-MS)[124]、傅立叶变换回旋共振质谱(Fourier transform ion cyclotron resonance mass spectrometry, FTICR-MS)[125,126],构成了土壤有机质波谱化和定量化分析方法,获得高分辨率有机质组成分子信息[127,128,129].这些高分辨率技术可以用于土地利用及管理变化下,外源有机物质进入土壤后的去向及其固定途径的解析研究.土壤有机质分子的自身结构及其化学性质强烈受制于有机质的降解和稳定机制,有机质分子本身的碳氮比、烷基碳和芳香碳比例等指标都可以指示有机质的分解抑或稳定程度[37].因此,在很长的一段时间内,有机质的分子结构性质被用来预测有机质在土壤中的分解速率[25],常被认为属于有机质的品质范畴(SOM quality),这也是研究者们试图解析土壤有机质分子结构的动力之一.例如,蛋白质和部分糖类等易分解有机质分子在土壤中的周转时间仅数十年[126,130],但可在土壤稳定性碳库中检出[44].同样,普遍认为化学性质稳定的木质素,在土壤中的周转周期要比其他有机质分子短[58].越来越多的检出研究证据表明,土壤有机质的保持和稳定还可能与生物有机分子的分子间相互作用及保护有关.正是基于生物标志物有机分子的降解规律,Incerti等[131]开发出新的有机质分解模型——OMDY,尽管还未得到足够关注. ...
Soil organic matter in its native state: Unravelling the most complex biomaterial on earth
1
2016
... 通过近10多年来的发展,湿化学法结合气相色谱—质谱联用仪法,即土壤中植物标志物提取—萃取—浓缩法及GC/MS定性/定量鉴定法联合技术已经基本完善,可满足土壤有机质生命来源分子的追踪[96,97].Masoom等[132]提出了综合多态核磁共振分析法来探讨天然土壤有机质的分子存在状态,这种分析揭示了土壤有机质基本上是生命代谢或降解过程留下的有机大分子,土水界面主要是亲水的脂类和碳水化合物,而土壤(颗粒)内部是疏水的木质素和微生物代谢产物,后者又可以吸附和固定复杂的疏水有机分子.这也为理解疏水有机分子的固定是土壤固碳机制之一提供了帮助[133],揭示了生命来源的有机分子在土壤粒子空间的连续分布谱系,成为理解土壤有机质整体的新视角. ...
Increased soil organic carbon sequestration through hydrophobic protection by humic substances
1
2002
... 通过近10多年来的发展,湿化学法结合气相色谱—质谱联用仪法,即土壤中植物标志物提取—萃取—浓缩法及GC/MS定性/定量鉴定法联合技术已经基本完善,可满足土壤有机质生命来源分子的追踪[96,97].Masoom等[132]提出了综合多态核磁共振分析法来探讨天然土壤有机质的分子存在状态,这种分析揭示了土壤有机质基本上是生命代谢或降解过程留下的有机大分子,土水界面主要是亲水的脂类和碳水化合物,而土壤(颗粒)内部是疏水的木质素和微生物代谢产物,后者又可以吸附和固定复杂的疏水有机分子.这也为理解疏水有机分子的固定是土壤固碳机制之一提供了帮助[133],揭示了生命来源的有机分子在土壤粒子空间的连续分布谱系,成为理解土壤有机质整体的新视角. ...
Humusica 1, article 1: Essential bases-Vocabulary
2
2017
... 在生态系统中,无论来自地上部还是地下部生物生命活动留下的残余物或者代谢产物,在土壤中随时随地处于不同程度微生物接触而降解的过程,因此瞬时的土壤有机质应该是所有曾经及正在土壤中活动的生命代谢产物或有机体残余,仅是不同程度微生物降解的化合物的集合,构成了土壤有机质系统.不同土壤所处生态系统演替过程各异,作为有机质输入来源的植物种类及其比例差异在不同生态系统土壤中差异很大,欧洲学界将此抽象为腐殖质系统及其形态类型(humus system and forms) (这里的腐殖质指土壤中呈暗色的稳定有机质[63]在土壤景观中的不同生态关系,而不是基于化学提取的传统腐殖质概念)[134].故而任何一种土壤中,植物天然生物标志物分子丰度及组成都可能反映特定系统的土壤有机质特点,更可以用来追溯有机质来源或者生态系统演化历程.除了前述的植物生物标志物外,养殖动物粪便有机质中的生物标志物系列也已建立,采用高温气质联用检测可以检测到施用奶牛粪便土壤中的动物生物标志物5β-stanols (n~ n ×102 ng/g),从而可以追踪养殖废弃物对土壤的有机污染[135].当然,土壤中还可能存在其他来源有机物质,如农药、抗生素和农膜等有机化合物污染,这些分子可以采用气相色谱法直接检测出来.早在20世纪80年代初,就有学者报道土壤中人为源化合物(污染化合物,xenobiotics)多达30多种,其在土壤有机质中可能高达10%~30%[136]. ...
... 表征和认识土壤有机质的本质,首先是探析生态系统中伴随土壤形成和发育的生命活动足迹(不同生物起源有机分子的丰度和结构),其次是土壤中有机质与生命多样性保持的关系(如有机质分子保持与土壤生物选择性适应及利用的关系),最后才可以揭示土壤有机质的土壤功能与生态系统服务(如土壤固碳).鉴于土壤是陆地最大的生物多样性储库且食物多样性是食物链多样性的基础,土壤有机质将具有比微生物更大的多样性.因此,土壤有机质质量至少应该与土壤微生物的食物多样性和土壤团聚体稳定性相联系,这也从生态学角度提示了有机质分子研究的重要性.我们认为,土壤有机质在土壤中不仅是连续体,而且作为土壤核心的团聚体—有机质—微生物—生命活动的耦合结构,亦应呈连续体的分布系统(图4),特别是在长期发育演化而稳定的土壤中.在这方面,近10年来欧洲学术界对土壤腐殖质系统与腐殖质形态的大陆尺度的对比和整合研究[134],有可能对土壤与生命发育的关系提供更前沿的视角.这对未来分子有机质的研究和认识具有重要的借鉴作用. ...
Combined quantification of faecal sterols, stanols, stanones and bile acids in soils and terrestrial sediments by gas chromatography-mass spectrometry
1
2012
... 在生态系统中,无论来自地上部还是地下部生物生命活动留下的残余物或者代谢产物,在土壤中随时随地处于不同程度微生物接触而降解的过程,因此瞬时的土壤有机质应该是所有曾经及正在土壤中活动的生命代谢产物或有机体残余,仅是不同程度微生物降解的化合物的集合,构成了土壤有机质系统.不同土壤所处生态系统演替过程各异,作为有机质输入来源的植物种类及其比例差异在不同生态系统土壤中差异很大,欧洲学界将此抽象为腐殖质系统及其形态类型(humus system and forms) (这里的腐殖质指土壤中呈暗色的稳定有机质[63]在土壤景观中的不同生态关系,而不是基于化学提取的传统腐殖质概念)[134].故而任何一种土壤中,植物天然生物标志物分子丰度及组成都可能反映特定系统的土壤有机质特点,更可以用来追溯有机质来源或者生态系统演化历程.除了前述的植物生物标志物外,养殖动物粪便有机质中的生物标志物系列也已建立,采用高温气质联用检测可以检测到施用奶牛粪便土壤中的动物生物标志物5β-stanols (n~ n ×102 ng/g),从而可以追踪养殖废弃物对土壤的有机污染[135].当然,土壤中还可能存在其他来源有机物质,如农药、抗生素和农膜等有机化合物污染,这些分子可以采用气相色谱法直接检测出来.早在20世纪80年代初,就有学者报道土壤中人为源化合物(污染化合物,xenobiotics)多达30多种,其在土壤有机质中可能高达10%~30%[136]. ...
Incorporation of xenobiotics into soil humus
1
1983
... 在生态系统中,无论来自地上部还是地下部生物生命活动留下的残余物或者代谢产物,在土壤中随时随地处于不同程度微生物接触而降解的过程,因此瞬时的土壤有机质应该是所有曾经及正在土壤中活动的生命代谢产物或有机体残余,仅是不同程度微生物降解的化合物的集合,构成了土壤有机质系统.不同土壤所处生态系统演替过程各异,作为有机质输入来源的植物种类及其比例差异在不同生态系统土壤中差异很大,欧洲学界将此抽象为腐殖质系统及其形态类型(humus system and forms) (这里的腐殖质指土壤中呈暗色的稳定有机质[63]在土壤景观中的不同生态关系,而不是基于化学提取的传统腐殖质概念)[134].故而任何一种土壤中,植物天然生物标志物分子丰度及组成都可能反映特定系统的土壤有机质特点,更可以用来追溯有机质来源或者生态系统演化历程.除了前述的植物生物标志物外,养殖动物粪便有机质中的生物标志物系列也已建立,采用高温气质联用检测可以检测到施用奶牛粪便土壤中的动物生物标志物5β-stanols (n~ n ×102 ng/g),从而可以追踪养殖废弃物对土壤的有机污染[135].当然,土壤中还可能存在其他来源有机物质,如农药、抗生素和农膜等有机化合物污染,这些分子可以采用气相色谱法直接检测出来.早在20世纪80年代初,就有学者报道土壤中人为源化合物(污染化合物,xenobiotics)多达30多种,其在土壤有机质中可能高达10%~30%[136]. ...
Impact of agricultural management practices on the nutrient supply potential of soil organic matter under long-term farming systems
1
2018
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
Electrospray ionization mass spectrometry of terrestrial humic substances and their size fractions
2
2003
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
... [138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
Microbial community structure varies across soil organic matter aggregate pools during tropical land cover change
1
2014
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
Different chemical composition of free light, occluded light and extractable SOM fractions in soils of Cerrado and tilled and untilled fields, Minas Gerais, Brazil: A pyrolysis‐GC/MS study
1
2011
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
Molecular changes in Particulate Organic Matter (POM) in a typical Chinese paddy soil under different long‐term fertilizer treatments
2
2010
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Linking soil fungal community structure and function to soil organic carbon chemical composition in intensively managed subtropical bamboo forests
1
2017
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
Aliphatic lipids released from a soil humin after enzymatic degradation of cellulose
1
1998
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics
1
2004
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
68: A4-A9
1
2014
... 因生命活动输入土壤的有机物分子受不同微生物区系降解和团聚体更新集合保护的连续影响,因不同降解而残留于土壤的生命分子既与微生物生长及其活性的变化密切联系,又与自然条件与管理下土壤团聚体的形成和稳定息息相关[137,138].GC/MS方法的成熟与团聚体分离制备的完善,使得土壤有机质研究得以探测分子水平化合物信息,推动了土壤有机质研究进入分子结构与团聚体结构同步研究[138,139,140,141]的新时代.这一方面有助于理解土壤有机质的生命来源本质,另一方面可能重新审视土壤有机质分子与微生物分子生态的紧密联系[142,143],这可能构成土壤学中微域生物地理格局的前沿热点[144,145]. ...
The contentious nature of soil organic matter
1
2015
... Lehmann等[146]发表了一篇观点性论文,提出了土壤有机质的连续体模型(Soil Continuum Model, SCM),提示着土壤有机质组学框架的发展前景.该模型提出的观点涉及2个方面:土壤有机质分子的微生物分解与有机分子在团聚体中的分布和空间隔离有关;有机质分子在团聚体中的存在与其和土壤矿物结合保护而避免微生物挖掘利用有关.因此,进入土壤的有机质物质的分解序列与其在团聚体中分布和结合稳定的序列存在契合关系.于是,土壤中有机质是一系列既处于不同分解阶段又结合或保护于不同粒径团聚体生命来源的有机分子集合.考虑到有机物质的分解程度和微生物参与分解的区系序列,分解程度较低的生物大分子和主要参与初期分解的真菌及其残体多存在于粒径较大的团聚体,而充分降解释放的较小分子以及主要参与后期分解的细菌及其残留物趋向于向较小团聚体集中.团聚体结构中土壤微域生境多样性,可能赋予了土壤有机质的分子多样性与微生物区系及种群的多样性.在本体土壤中,团聚体是连续的,因而土壤中有机分子分布以及相应的微生物分布都是土壤中连续分布的整体(continuum). ...
Nano-pore distribution of biochar and soil aggregates revealed with the technology of nuclear magnetic resonance cryoporometry
1
2018
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
核磁冻融微孔度技术应用于测定生物质炭及土壤团聚体纳米孔隙
1
2018
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Stabilization mechanisms of soil organic matter: Implications for C-saturation of soils
1
2002
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
The turnover of soil organic matter in some of the Rothamsted classical experiments
1
1977
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
What happens to terrestrial organic matter in the ocean?
1
1997
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
The soil mineralogy of lead at Horace's villa
2
2004
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
... [151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Soil organic matter quality interpreted thermodynamically
1
1999
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Latitudinal variation of leaf stomatal traits from species to community level in forests: Linkage with ecosystem productivity
1
2015
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Microbial activity promoted with organic carbon accumulation in macroaggregates of paddy soils under long-term rice cultivation
1
2016
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
Soil carbon sequestration with bioactivity: A new emerging frontier for sustainable soil management
2
2015
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
... 根据上述原理,土壤学基础理论研究的对象将主要是以团聚体为基础的有机质—微生物丰度、结构、组成及其耦合协同关系的解剖分析,这将是土壤学关于土壤可持续管理的核心理论基础.2015年在德国哥廷根举行的第五届国际土壤有机质大会附设了一个专门研讨会,讨论土壤团聚体及其有机质分组方法框架问题.欧盟地学联盟2018年年会(European Geosciences Union General Assembly 2018, Vienna, Austria),由德国科学基金资助项目(MAD soil, development and turnover of soil micro-aggregates, DFG RU2179)发起,德国科学家主持召开了土壤团聚体研究的专门研讨会.由此看来,团聚体的研究是深入和微观研究土壤有机质的基础工作.特别是与有机质微生物分室保持及隔离稳定有关[155]的土壤团聚体内外孔隙系统将是团聚体研究的新内容. ...
土壤碳固定与生物活性: 面向可持续土壤管理的新前沿
2
2015
... SCM观点承认团聚体可达性导致的有机质分子限制和微生物对有机质分子降解的团聚体空间限制.已知土壤团聚体中孔隙有微团聚体内部孔隙(其直径可能多在纳米尺寸)和微团聚体间孔隙(其大小可达至微米)[147],这样的团聚体孔隙分布可能造成矿物质—有机质—微生物组合分配的特征性差异.来自植物和土壤动物的有机质原始物料进入土壤的初期,首先被(食物网初级分解者)破碎分解成小块,而颗粒态碳被物理保护于大团聚体[66,148],这一过程可在有机质原始投入1年内观察到[149].而分解释放的单体有机质分子可能进入微团聚体内部小孔隙中,进而被矿物质结合而保护.在大团聚体内部,微生物胞外酶可将尺径相对较小(通常小于600道尔顿)的动植物残体或有机质初分解产物[150]跨送到微生物细胞壁供细菌等微生物分解利用[151].因此,遵循热力学定律,在具有正常生物活性的土壤中,应存在周转周期序列上处于不同降解程度的一系列有机质组分[152],从含能量高而分子量相对较大的化合物逐级降解为含能量低分子量相对较小的化合物[151],分布于大小序列递降的团聚体颗粒中.因此,短期管理或有机物投入下的有机质积累,将首先表现为颗粒态有机质的增加和物理保护稳定,并伴随着真菌丰度相对于细菌的增加.这种现象在关于植稻时间序列[153,154]和有机无机施肥的稻田[121,141]研究中观察到.这个土壤有机质连续体模型提供了一个深刻而系统理解土壤固碳与生物活性及生态系统功能[155]的逻辑框架. ...
... 根据上述原理,土壤学基础理论研究的对象将主要是以团聚体为基础的有机质—微生物丰度、结构、组成及其耦合协同关系的解剖分析,这将是土壤学关于土壤可持续管理的核心理论基础.2015年在德国哥廷根举行的第五届国际土壤有机质大会附设了一个专门研讨会,讨论土壤团聚体及其有机质分组方法框架问题.欧盟地学联盟2018年年会(European Geosciences Union General Assembly 2018, Vienna, Austria),由德国科学基金资助项目(MAD soil, development and turnover of soil micro-aggregates, DFG RU2179)发起,德国科学家主持召开了土壤团聚体研究的专门研讨会.由此看来,团聚体的研究是深入和微观研究土壤有机质的基础工作.特别是与有机质微生物分室保持及隔离稳定有关[155]的土壤团聚体内外孔隙系统将是团聚体研究的新内容. ...
The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization
1
2016
... 此外,土壤连续体模型肯定了微生物在土壤有机质更新和分子组成结构变化中的作用.在该模型中,微生物不仅是土壤有机质分解更新的推动者,同时微生物代谢产物及微生物残体加入土壤有机质,真菌孢子壁或几丁质作为化学稳定分子存在于大团聚体,或仍属于生物源未降解的有机分子.而在微团聚体中,则有更多细菌参与代谢过程,并且其产物残留于土壤中.因此,土壤中瞬时检出的有机物质,一方面可能是分解微生物区系的“食物舍弃”,而另一方面也表现为土壤矿物或者团聚体的“捕获”或者“纳贤”.最近,有较多的研究报道了细组分中微生物来源的有机分子对土壤有机质的贡献,在某些土壤中可能高达50%以上[156].但是,在连续团聚体组分中,作为微生物基质的有机分子的异质性将导致微生物区系和种群的多样性关系.作为连续体,有机质分子化学和微生物分子生态在丰度、组成、结构及其多样性上可能共同演化为一种协同耦合结构,我们曾称之为土壤生产力功能结构[157].这种有机质—微生物的连续体分布将可能构成土壤过程、功能及生态系统服务的新理论框架.当然,因为有机质和矿物基质的空间结合状况、氧化还原条件、微生物生态和与矿物表面水分和温度条件下的相互作用等方面的差异,这种连续体在不同土壤中的特征结构及功能性会有显著差异[158]. ...
Organic carbon sequestration and its co-benefits in China’s croplands
1
2013
... 此外,土壤连续体模型肯定了微生物在土壤有机质更新和分子组成结构变化中的作用.在该模型中,微生物不仅是土壤有机质分解更新的推动者,同时微生物代谢产物及微生物残体加入土壤有机质,真菌孢子壁或几丁质作为化学稳定分子存在于大团聚体,或仍属于生物源未降解的有机分子.而在微团聚体中,则有更多细菌参与代谢过程,并且其产物残留于土壤中.因此,土壤中瞬时检出的有机物质,一方面可能是分解微生物区系的“食物舍弃”,而另一方面也表现为土壤矿物或者团聚体的“捕获”或者“纳贤”.最近,有较多的研究报道了细组分中微生物来源的有机分子对土壤有机质的贡献,在某些土壤中可能高达50%以上[156].但是,在连续团聚体组分中,作为微生物基质的有机分子的异质性将导致微生物区系和种群的多样性关系.作为连续体,有机质分子化学和微生物分子生态在丰度、组成、结构及其多样性上可能共同演化为一种协同耦合结构,我们曾称之为土壤生产力功能结构[157].这种有机质—微生物的连续体分布将可能构成土壤过程、功能及生态系统服务的新理论框架.当然,因为有机质和矿物基质的空间结合状况、氧化还原条件、微生物生态和与矿物表面水分和温度条件下的相互作用等方面的差异,这种连续体在不同土壤中的特征结构及功能性会有显著差异[158]. ...
2
1997
... 此外,土壤连续体模型肯定了微生物在土壤有机质更新和分子组成结构变化中的作用.在该模型中,微生物不仅是土壤有机质分解更新的推动者,同时微生物代谢产物及微生物残体加入土壤有机质,真菌孢子壁或几丁质作为化学稳定分子存在于大团聚体,或仍属于生物源未降解的有机分子.而在微团聚体中,则有更多细菌参与代谢过程,并且其产物残留于土壤中.因此,土壤中瞬时检出的有机物质,一方面可能是分解微生物区系的“食物舍弃”,而另一方面也表现为土壤矿物或者团聚体的“捕获”或者“纳贤”.最近,有较多的研究报道了细组分中微生物来源的有机分子对土壤有机质的贡献,在某些土壤中可能高达50%以上[156].但是,在连续团聚体组分中,作为微生物基质的有机分子的异质性将导致微生物区系和种群的多样性关系.作为连续体,有机质分子化学和微生物分子生态在丰度、组成、结构及其多样性上可能共同演化为一种协同耦合结构,我们曾称之为土壤生产力功能结构[157].这种有机质—微生物的连续体分布将可能构成土壤过程、功能及生态系统服务的新理论框架.当然,因为有机质和矿物基质的空间结合状况、氧化还原条件、微生物生态和与矿物表面水分和温度条件下的相互作用等方面的差异,这种连续体在不同土壤中的特征结构及功能性会有显著差异[158]. ...
... 从本源上说,土壤物质就是大小、结构和功能(反应性和生物活性)不同的、矿物质—有机质—微生物有序集合的土壤团聚体颗粒集合.其形成、发育和稳定属于土壤的生物动力学过程(biodynamics)[158],是土壤形成及相关生命过程的基础.植物等生物活动剩余物输入的有机物质,一方面作为原始材料驱动土壤有机质形成和稳定;另一方面,它们被土壤中的食物网,特别是微生物分解利用,并提供其活动的能源.而土壤团聚体的形成、稳定和更新提供和维持了有机质—微生物—生物功能的偶联关系及其稳定,是土壤发挥生物地球化学作用和生物多样性保持[6]的根本.因此,土壤有机质连续体实际上是一种保持有机质—微生物—生物活性良好协同的微域机制框架. ...
Labile organic matter fractions as central components of the quality of agricultural soils: An overview
1
2005
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Soil organic matter content and quality: Effects of fertilizer applications, burning and trash retention on a long-term sugarcane experiment in South Africa
1
2002
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Soil organic matter fractions as early indicators for carbon stock changes under different land-use?
1
2005
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Qualitative and quantitative soil organic matter estimation for sustainable soil management
1
2018
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Biochar for sustainable soil management: Biomass technology and industry from soil perspectives
1
2018
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
生物质炭与土壤可持续管理: 从土壤问题到生物质产业
1
2018
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Biochar compound fertilizer increases nitrogen productivity and economic benefits but decreases carbon emission of maize production
1
2017
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Yield, quality and nutrient uptake of chinese cabbage under organ/mineral water soluble fertilizer based on biochar extracts derived from pyrolyzed biomass
1
2018
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
基于生物质裂解活性有机物的有机—无机水溶肥对空心菜产量、品质及养分的影响
1
2018
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Identification of black carbon derived structures in a volcanic ash soil humic acid by Fourier transform ion cyclotron resonance mass spectrometry
1
2004
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Reducing capacity of water extracts of biochars and their solubilization of soil Mn and Fe
2
2014
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
... [167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Functions of organic matter in polluted soils: The effect of organic amendments on phytoavailability of heavy metals
1
2018
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Induced systemic resistance in tomato (Solanum lycopersicum) against Botrytis cinerea by biochar amendment involves jasmonic acid signaling
2
2015
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
... [169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Soil organic matter mobilization by root exudates
1
2000
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Induction of systemic resistance in plants by biochar, a soil-applied carbon sequestering agent
1
2010
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Soil Micromorphology: Development and Application
1
1991
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
土壤微形态学——发展及应用
1
1991
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Compost, vermicompost, and compost tea: Feeding the soil on the organic farm
1
2011
... 土壤有机质一直是土壤学研究中的一个物质黑箱,从定性到定量认识土壤有机质的本质及品质的研究经久不衰[159,160,161,162].从生物质炭土壤改良[163]和秸秆炭基肥的生态农业试验[164,165]得到启示,土壤有机质发挥着以下功能:结构建成功能,作为可塑有机构件(building blocks)参与形成矿质—有机复合体和土壤团聚体[166].反应性功能,即通过包被矿物质表面,修饰了土壤的颗粒表面,呈现出氧化还原活性、可变电荷的吸附活性以及对无机元素的螯合和对有机化合物的捕获固定活性[167,168].生物激活功能,既刺激土壤生物(包括根系)代谢活动的功能[169],也包括可矿化有机质对土壤微生物的激发效应[48];例如,部分可溶性有机分子(可能是土壤微生物代谢的来源,例如吲哚乙酸)对植物根系是刺激效应,称之为植物激素类有机质[170].最近关于生物质炭可提取有机质对植物生长的效应研究[122,171],进一步明确至少部分土壤可溶性有机物可以刺激植物生长,促进植物生长与健康,进而对植物病害具有抵御作用(system resistance)[169],尽管这种效应还可能是由于这些活性有机质所具有的氧化还原(电子迁移)活性[167].当然,从结构性有机质、反应性有机质到促生性有机质,从有机质的可溶解性、可分解性和可稳定性,从年轻的活泼有机碳到年老的惰性有机碳,这些可能不是组分或者分子的差异,或者不是由分子性质决定的,而是由其存在状态、存在位置或者与矿物质的结合及其团聚体的保护性决定的,即由各种营力的可达性(accessibility)决定的.从土壤微形态学[172]的视角,土壤有机质既可以通过与矿物质的结合而表现出骨骼(solid skeleton)作用,通过团聚体的包裹和黏附于表面;又起到土壤的肌腱(muscle tendon)作用,而游离于溶液的有机质具有生物活性的营养汁液(nutrition juice)作用[173]. ...
Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems
1
1995
... 土壤有机质的质量是长期困扰土壤学家的科学问题,众说纷纭.土壤腐殖质学说曾用E4/E6值代表其假设的腐殖化程度,土壤肥料学中常用的C/N值则考虑了有机质的生物化学活泼性,而有机质不稳定程度(lability,或者分解性)及其衍生出的有机质可持续管理指数可衡量有机质被氧化分解的潜力[174].土壤学中更常见的类似提法是有机物源质量(如litter quality)[175],其仅关注微生物分解利用性.作为一种分子探索,有研究提出用中性洗涤纤维(neutral detergent fibre)与粗蛋白(crude protein)分子比值[176]来衡量微生物分解性的土壤有机质基质质量(substrate quality).尽管早有土壤可溶性有机质的蛋白质组学指纹方法尝试[177],但在表征和权衡土壤有机质主要功能的基础上,如何从分子有机质角度表征土壤有机质的质量仍有待深入研究.看来,审视土壤有机质的物理、化学和生物学的不同状态及其意义,才能更好地联系和揭示土壤有机质的组成、结构和功能. ...
Plant litter quality affects the accumulation rate, composition, and stability of mineral-associated soil organic matter
1
2018
... 土壤有机质的质量是长期困扰土壤学家的科学问题,众说纷纭.土壤腐殖质学说曾用E4/E6值代表其假设的腐殖化程度,土壤肥料学中常用的C/N值则考虑了有机质的生物化学活泼性,而有机质不稳定程度(lability,或者分解性)及其衍生出的有机质可持续管理指数可衡量有机质被氧化分解的潜力[174].土壤学中更常见的类似提法是有机物源质量(如litter quality)[175],其仅关注微生物分解利用性.作为一种分子探索,有研究提出用中性洗涤纤维(neutral detergent fibre)与粗蛋白(crude protein)分子比值[176]来衡量微生物分解性的土壤有机质基质质量(substrate quality).尽管早有土壤可溶性有机质的蛋白质组学指纹方法尝试[177],但在表征和权衡土壤有机质主要功能的基础上,如何从分子有机质角度表征土壤有机质的质量仍有待深入研究.看来,审视土壤有机质的物理、化学和生物学的不同状态及其意义,才能更好地联系和揭示土壤有机质的组成、结构和功能. ...
Substrate quality influences organic matter accumulation in the soil silt and clay fraction
1
2016
... 土壤有机质的质量是长期困扰土壤学家的科学问题,众说纷纭.土壤腐殖质学说曾用E4/E6值代表其假设的腐殖化程度,土壤肥料学中常用的C/N值则考虑了有机质的生物化学活泼性,而有机质不稳定程度(lability,或者分解性)及其衍生出的有机质可持续管理指数可衡量有机质被氧化分解的潜力[174].土壤学中更常见的类似提法是有机物源质量(如litter quality)[175],其仅关注微生物分解利用性.作为一种分子探索,有研究提出用中性洗涤纤维(neutral detergent fibre)与粗蛋白(crude protein)分子比值[176]来衡量微生物分解性的土壤有机质基质质量(substrate quality).尽管早有土壤可溶性有机质的蛋白质组学指纹方法尝试[177],但在表征和权衡土壤有机质主要功能的基础上,如何从分子有机质角度表征土壤有机质的质量仍有待深入研究.看来,审视土壤有机质的物理、化学和生物学的不同状态及其意义,才能更好地联系和揭示土壤有机质的组成、结构和功能. ...
A proteomic fingerprint of dissolved organic carbon and of soil particles
1
2005
... 土壤有机质的质量是长期困扰土壤学家的科学问题,众说纷纭.土壤腐殖质学说曾用E4/E6值代表其假设的腐殖化程度,土壤肥料学中常用的C/N值则考虑了有机质的生物化学活泼性,而有机质不稳定程度(lability,或者分解性)及其衍生出的有机质可持续管理指数可衡量有机质被氧化分解的潜力[174].土壤学中更常见的类似提法是有机物源质量(如litter quality)[175],其仅关注微生物分解利用性.作为一种分子探索,有研究提出用中性洗涤纤维(neutral detergent fibre)与粗蛋白(crude protein)分子比值[176]来衡量微生物分解性的土壤有机质基质质量(substrate quality).尽管早有土壤可溶性有机质的蛋白质组学指纹方法尝试[177],但在表征和权衡土壤有机质主要功能的基础上,如何从分子有机质角度表征土壤有机质的质量仍有待深入研究.看来,审视土壤有机质的物理、化学和生物学的不同状态及其意义,才能更好地联系和揭示土壤有机质的组成、结构和功能. ...
Soil zymography as a powerful tool for exploring hotspots and substrate limitation in undisturbed subsoil
1
2018
... 伴随着土壤生物物理结构的发育和土壤生物生命区系的更新周转,土壤中的有机物质循环兼具了物质流动和信息流动.随着土壤微域鉴定技术和Nano-SIMS的应用,可视化和原位化的土壤团聚体生物物理结构的重现成为可能.结合土壤团聚体的非干扰分离制备技术,土壤有机质分子组成与土壤团聚体物理结构的耦合发育可能构建土壤有机质的分子序列,并逐渐成熟为团聚体—有机质—微生物—酶活性等要素的分子组学.配合业已成熟的酶活性组学技术(Zymography)[178],这种有机质组学技术将可能回答土壤学的关键问题:如何解析作为连续体的土壤有机质分子大家庭的基本构成与存在状态,如何表征其与土壤的团聚体结构、微生物区系结构及其生态系统功能活性的联系,如何刻画和鉴别不同生态系统、不同利用和管理下土壤有机质的差异,如何建立土壤可持续管理的土壤分子有机质本质及品质.纵观当今的土壤学研究,土壤腐殖质学说已不能适应土壤有机质过程和功能的认识[179],特别是土壤与生命耦合协同关系的探索;腐殖质组学虽然理论框架和方法基本成型,但其对土壤有机质的存在状况及其与生命过程的联系尚存疑隙;土壤有机质分子研究已经走出萌芽阶段,借助于现代高分辨率分子分析技术,已经凸显为新时期全球土壤基础研究的新兴领域. ...
Humic matter in soil and the environment, principles and controversies
1
2015
... 伴随着土壤生物物理结构的发育和土壤生物生命区系的更新周转,土壤中的有机物质循环兼具了物质流动和信息流动.随着土壤微域鉴定技术和Nano-SIMS的应用,可视化和原位化的土壤团聚体生物物理结构的重现成为可能.结合土壤团聚体的非干扰分离制备技术,土壤有机质分子组成与土壤团聚体物理结构的耦合发育可能构建土壤有机质的分子序列,并逐渐成熟为团聚体—有机质—微生物—酶活性等要素的分子组学.配合业已成熟的酶活性组学技术(Zymography)[178],这种有机质组学技术将可能回答土壤学的关键问题:如何解析作为连续体的土壤有机质分子大家庭的基本构成与存在状态,如何表征其与土壤的团聚体结构、微生物区系结构及其生态系统功能活性的联系,如何刻画和鉴别不同生态系统、不同利用和管理下土壤有机质的差异,如何建立土壤可持续管理的土壤分子有机质本质及品质.纵观当今的土壤学研究,土壤腐殖质学说已不能适应土壤有机质过程和功能的认识[179],特别是土壤与生命耦合协同关系的探索;腐殖质组学虽然理论框架和方法基本成型,但其对土壤有机质的存在状况及其与生命过程的联系尚存疑隙;土壤有机质分子研究已经走出萌芽阶段,借助于现代高分辨率分子分析技术,已经凸显为新时期全球土壤基础研究的新兴领域. ...




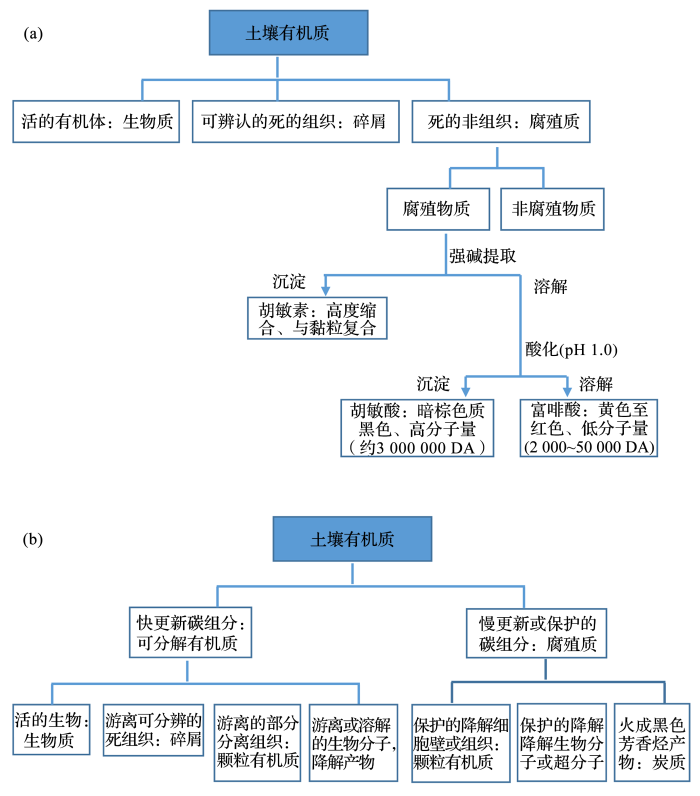

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}