Varying planetary heat sink led to global-warming slowdown and acceleration
1
2014
... 中—深层海洋是巨大的热储库,自20世纪中期以来,中—深层海洋吸收了全球变暖约1/3的热量[1 ] ,产生了全球性的气候效应.如1997—2015年出现的“全球气候变暖停滞”现象就被认为与海洋储热机制的变化有关[2 ,3 ] .由于缺少中—深层海洋观测数据以及气候系统之间复杂的遥相关作用,目前还无法弄清真正的调控机制.因为人类时间尺度上的中—深层海洋环境演化信息很难从沉积物中获取,原因是绝大多数地方海洋沉积物的沉积速率低、生物扰动严重.因此,利用冷水珊瑚重建中—深层海水温度的高分辨率变化,有望弥补中—深层海洋年际、年代际尺度上数据记录的空白. ...
Pacific origin of the abrupt increase in Indian Ocean heat content during the warming hiatus
1
2015
... 中—深层海洋是巨大的热储库,自20世纪中期以来,中—深层海洋吸收了全球变暖约1/3的热量[1 ] ,产生了全球性的气候效应.如1997—2015年出现的“全球气候变暖停滞”现象就被认为与海洋储热机制的变化有关[2 ,3 ] .由于缺少中—深层海洋观测数据以及气候系统之间复杂的遥相关作用,目前还无法弄清真正的调控机制.因为人类时间尺度上的中—深层海洋环境演化信息很难从沉积物中获取,原因是绝大多数地方海洋沉积物的沉积速率低、生物扰动严重.因此,利用冷水珊瑚重建中—深层海水温度的高分辨率变化,有望弥补中—深层海洋年际、年代际尺度上数据记录的空白. ...
Abrupt pre-Bolling-Allerod warming and circulation changes in the deep ocean
1
2014
... 中—深层海洋是巨大的热储库,自20世纪中期以来,中—深层海洋吸收了全球变暖约1/3的热量[1 ] ,产生了全球性的气候效应.如1997—2015年出现的“全球气候变暖停滞”现象就被认为与海洋储热机制的变化有关[2 ,3 ] .由于缺少中—深层海洋观测数据以及气候系统之间复杂的遥相关作用,目前还无法弄清真正的调控机制.因为人类时间尺度上的中—深层海洋环境演化信息很难从沉积物中获取,原因是绝大多数地方海洋沉积物的沉积速率低、生物扰动严重.因此,利用冷水珊瑚重建中—深层海水温度的高分辨率变化,有望弥补中—深层海洋年际、年代际尺度上数据记录的空白. ...
On the isotopic chemistry of carbonates and a paleotemperature scale
1
1950
... 碳酸盐氧同位素温度计是重建古水温的重要方法.早在20世纪50年代,就发现无机成因和生物成因的碳酸盐与水体之间存在温度控制的氧同位素分馏[4 ,5 ] .这种同位素温度计在有孔虫和浅水珊瑚中已经得到广泛应用,现在这种应用开始延伸到冷水珊瑚上. ...
Revised carbonate-water isotopic temperature scale
1
1953
... 碳酸盐氧同位素温度计是重建古水温的重要方法.早在20世纪50年代,就发现无机成因和生物成因的碳酸盐与水体之间存在温度控制的氧同位素分馏[4 ,5 ] .这种同位素温度计在有孔虫和浅水珊瑚中已经得到广泛应用,现在这种应用开始延伸到冷水珊瑚上. ...
Reefs of the deep: The biology and geology of cold-water coral ecosystems
1
2006
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
Deep-sea scleractinian coral age and depth distributions in the WN Atlantic for the last 225 thousand years
1
2007
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
Movement of deep-sea coral populations on climatic timescales
1
2013
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
Clumped isotope composition of cold-water corals: A role for vital effects?
6
2016
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
... 碳酸盐样品的二元同位素(Δ47 ,也称耦合同位素)也广泛应用于古水温重建,由于计算时不需要知晓周围水体的同位素成分,且二元同位素的变化与温度变化直接相关,因此是一种很有应用前景的古温度指标[9 ,33 ] .对于碳酸盐样品来说,碳酸根中的轻同位素会被重同位素替换,如12 C会被替换成13 C,16 O会被替换为17 O和18 O,使得CO3 2 - 表1 )[37 ] .在热力学平衡状态下,碳酸盐矿物晶格中的13 C和18 O倾向于进入同一基团中,形成13 C-18 O键,主要存在于13 C18 O16 O2 2 - 13 C-18 O键的丰度是碳酸盐形成时温度的函数,而与周围水体的同位素组成无关.实验研究中,碳酸钙与磷酸反应生成CO2 ,主要根据生成的13 C18 O16 O含量评估碳酸根中13 C-18 O键的丰度.碳酸钙中形成13 C18 O16 O2 2 - [9 ] : ...
... [9 ]: ...
... 之后不同的经验公式接踵出现[39 ,40 ,41 ] ,有孔虫、腕足类、双壳类等生物成因碳酸盐的Δ47 投点结果更接近Ghosh的校正方程[40 ,41 ] .但浅水珊瑚和某些属种冷水珊瑚的二元同位素都展现出显著的生命效应,如在冷水珊瑚Balanophyllia , Caryophyllia , Dasmosmilia ,Desmophyllum ,Enallopsammia 和Javania 等属种中[9 ] ,Δ47 比Ghosh校正方程计算的同等温度下的平衡值偏高,一些样品甚至高出约0.05‰(相当于温度被低估了约9 ℃).因此,使用二元同位素Δ47 重建古水温时也需要对冷水珊瑚属种进行生命效应校正. ...
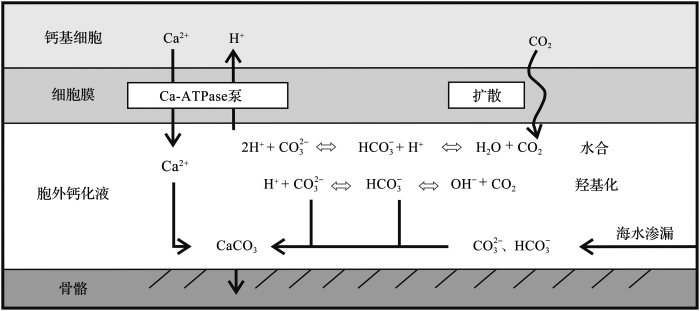
... 碳酸盐从溶解无机碳中结晶之前与水保持着同位素平衡.钙化过程发生在弱碱性的胞外钙化液中,CO
2 通过扩散作用进入其中,而HCO
3 - 和CO
3 2 - 无法透过细胞膜
[18 ] (
图4 ).胞外钙化液中的HCO
3 - 由CO
2 与H
2 O或OH
- 反应形成,CO
2 与H
2 O反应(水合反应)形成的HCO
3 - ,其δ
18 O
PDB 为-3.2‰;由CO
2 与OH
- 反应(羟基化反应)形成的HCO
3 - ,δ
18 O
PDB 为-15.8‰.2个反应生成HCO
3 - 的比例取决于溶液的pH,如pH=8.9时,由水合反应生成的HCO
3 - 占24%,羟基化反应生成的HCO
3 - 占76%;而pH=10时,2个反应生成HCO
3 - 的比例分别为2%和98%
[20 ] .两种情况下,HCO
3 - 综合表现出的δ
18 O
PDB 分别为-12.8‰和-15.5‰.ECF中升高的pH不仅导致溶液δ
18 O偏负,同时使CO
2 扩散效应增强,溶液中δ
13 C也出现负偏,于是生成的珊瑚骨骼δ
18 O和δ
13 C均负偏.
图4 冷水珊瑚动力学分馏模型示意图<sup>[<xref ref-type="bibr" rid="R9">9</xref>]</sup> Kinetic fractionation model of cold-water coral calcification<sup>[<xref ref-type="bibr" rid="R9">9</xref>]</sup> Fig.4 ![]()
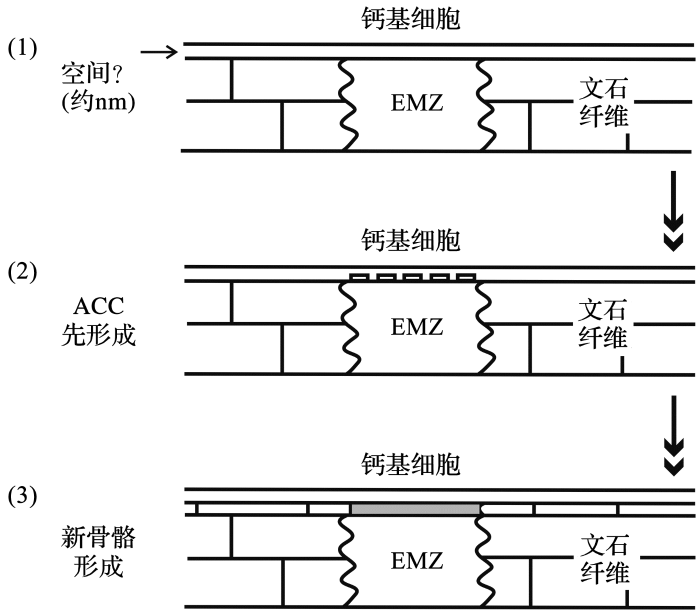
不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
... [
9 ]
Fig.4 ![]()
不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
Environmental and biological controls on Mg and Li in deep-sea scleractinian corals
1
300
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
Mg/Ca-temperature proxy in benthic foraminifera: New calibrations from the Florida Straits and a hypothesis regarding Mg/Li
1
2008
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
Sr/Ca and Mg/Ca vital effects correlated with skeletal architecture in a scleractinian Deep-Sea coral and the role of Rayleigh fractionation
1
261
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
Paleotemperatures from deep-sea corals: Overcoming ‘vital effects’
9
2000
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
... [13 ,14 ,15 ]. ...
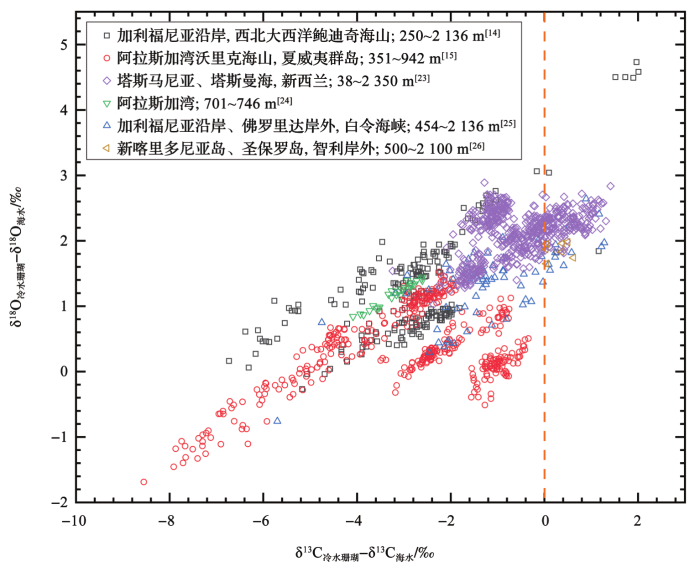
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... [13 ,14 ,15 ].这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... [13 ,15 ]: ...
... δ
18 O
cold-water coral -δ
18 O
seawater ,δ
13 C
cold-water cora -δ
13 C
seawater : δ
18 O and δ
13 C values are adjusted for local δ
18 O
seawater and δ
13 C
seawater values; The orange dashed line indicates δ
18 O axis at δ
13 C
cold-water coral -δ
13 C
seawater =0
Fig.1 ![]()
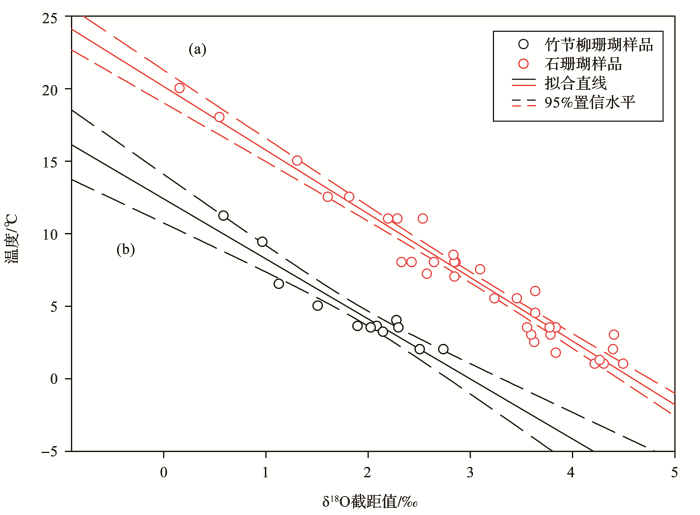
图2 <strong>δ<sup>18</sup>O</strong>截距值与温度之间的拟合关系<sup>[<xref ref-type="bibr" rid="R13">13</xref>,<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>]</sup> (a)石珊瑚δ18 O与温度间的拟合关系;(b)竹节柳珊瑚δ18 O与温度间的拟合关系 ...
... (a)石珊瑚δ
18 O与温度间的拟合关系;(b)竹节柳珊瑚δ
18 O与温度间的拟合关系
δ<sup>18</sup>O-temperature regressions from cold-water corals with 95% confidence intervals<sup>[<xref ref-type="bibr" rid="R13">13</xref>,<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>]</sup> (a) δ18 O-temperature regression for scleractinian corals;(b) δ18 O-temperature regression for bamboo corals ...
... 石珊瑚和竹节柳珊瑚的温度—氧同位素线性拟合线近乎平行,但截距值存在差异(图2 ),这与文石和高镁方解石两种矿物的结晶分馏过程有关[13 ,14 ,15 ] .在有分支(叉)结构的竹节柳珊瑚中,分支生长轴处δ18 O与δ13 C的亏损程度比主生长轴处更大,表现出更强的分馏效应.此外,靠近冷水珊瑚轴心的生长环带也会出现同位素显著亏损现象[14 ,15 ] . ...
... 利用微钻取冷水珊瑚粉末样时,通常有2种方式:一种沿切面径向自中心向边缘采样,另一种沿同心生长纹层采样,这2种取样方法获得的冷水珊瑚氧、碳稳定同位素都表现出线性关系[13 ,14 ,15 ] .利用截距法计算古水温时,前一种取样方式计算出的是冷水珊瑚生长周期内周围海水的平均水温,即几十年或几百年的平均温度;后者代表冷水珊瑚某一生长时间(段)内的古水温.因此,理论上沿着珊瑚不同生长纹层取样可以获得时间序列上的古水温变化.在实际操作中,由于冷水珊瑚单个生长环带仅有约10 μm宽[28 ] ,靠肉眼难以辨认,此外还受切面大小、微钻精度的影响,取到的样品多会横跨几个环带.一些较小的珊瑚样品取样数量也会受到限制,难以获得足够的样品数据建立起氧、碳同位素的可靠线性关系.因此,利用冷水珊瑚重建古水温演化序列的误差较大. ...
Temperature and vital effect controls on bamboo coral (Isididae ) isotope geochemistry: A test of the ‘lines method’
12
2011
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
... ,14 ,15 ]. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... ,14 ,15 ].这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... [
14 ,
15 ,
23 ,
24 ,
25 ,
26 ]
δ18 O冷水珊瑚 -δ18 O海水 、δ13 C冷水珊瑚 -δ13 C海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ13 C冷水珊瑚 -δ13 C海水 =0时的δ18 O轴 ...
... δ
18 O
冷水珊瑚 -δ
18 O
海水 、δ
13 C
冷水珊瑚 -δ
13 C
海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时的δ
18 O轴
δ<sup>13</sup>C versus δ<sup>18</sup>O in bamboo corals<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 Ocold-water coral -δ18 Oseawater ,δ13 Ccold-water cora -δ13 Cseawater : δ18 O and δ13 C values are adjusted for local δ18 Oseawater and δ13 Cseawater values; The orange dashed line indicates δ18 O axis at δ13 Ccold-water coral -δ13 Cseawater =0 ...
... δ
18 O
cold-water coral -δ
18 O
seawater ,δ
13 C
cold-water cora -δ
13 C
seawater : δ
18 O and δ
13 C values are adjusted for local δ
18 O
seawater and δ
13 C
seawater values; The orange dashed line indicates δ
18 O axis at δ
13 C
cold-water coral -δ
13 C
seawater =0
Fig.1 ![]()
图2 <strong>δ<sup>18</sup>O</strong>截距值与温度之间的拟合关系<sup>[<xref ref-type="bibr" rid="R13">13</xref>,<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>]</sup> (a)石珊瑚δ18 O与温度间的拟合关系;(b)竹节柳珊瑚δ18 O与温度间的拟合关系 ...
... (a)石珊瑚δ
18 O与温度间的拟合关系;(b)竹节柳珊瑚δ
18 O与温度间的拟合关系
δ<sup>18</sup>O-temperature regressions from cold-water corals with 95% confidence intervals<sup>[<xref ref-type="bibr" rid="R13">13</xref>,<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>]</sup> (a) δ18 O-temperature regression for scleractinian corals;(b) δ18 O-temperature regression for bamboo corals ...
... 石珊瑚和竹节柳珊瑚的温度—氧同位素线性拟合线近乎平行,但截距值存在差异(图2 ),这与文石和高镁方解石两种矿物的结晶分馏过程有关[13 ,14 ,15 ] .在有分支(叉)结构的竹节柳珊瑚中,分支生长轴处δ18 O与δ13 C的亏损程度比主生长轴处更大,表现出更强的分馏效应.此外,靠近冷水珊瑚轴心的生长环带也会出现同位素显著亏损现象[14 ,15 ] . ...
... [14 ,15 ]. ...
... 利用微钻取冷水珊瑚粉末样时,通常有2种方式:一种沿切面径向自中心向边缘采样,另一种沿同心生长纹层采样,这2种取样方法获得的冷水珊瑚氧、碳稳定同位素都表现出线性关系[13 ,14 ,15 ] .利用截距法计算古水温时,前一种取样方式计算出的是冷水珊瑚生长周期内周围海水的平均水温,即几十年或几百年的平均温度;后者代表冷水珊瑚某一生长时间(段)内的古水温.因此,理论上沿着珊瑚不同生长纹层取样可以获得时间序列上的古水温变化.在实际操作中,由于冷水珊瑚单个生长环带仅有约10 μm宽[28 ] ,靠肉眼难以辨认,此外还受切面大小、微钻精度的影响,取到的样品多会横跨几个环带.一些较小的珊瑚样品取样数量也会受到限制,难以获得足够的样品数据建立起氧、碳同位素的可靠线性关系.因此,利用冷水珊瑚重建古水温演化序列的误差较大. ...
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
Oxygen and carbon isotope fractionation in calcitic deep-sea corals: Implications for paleotemperature reconstruction
11
2014
... 除了北冰洋外,全球各大洋的中—深层海水中均发现有冷水珊瑚分布[6 ] .其生命周期可达数百年,甚至上千年,并且部分珊瑚品种发育生长纹层.依据目前的定年方法,发现有距今至少22.5万年的冷水珊瑚化石样品[7 ,8 ] .因此,冷水珊瑚是重建高分辨率中—深层海水温度演变的重要载体.冷水珊瑚碳酸盐骨骼的元素比值、氧碳稳定同位素以及二元同位素(clumped isotope)都与水温变化存在关系[9 ,10 ,11 ,12 ,13 ,14 ] .目前对两种珊瑚开展了较系统的同位素温度计研究:一类是文石质的石珊瑚,另一类是高镁方解石质的竹节柳珊瑚[13 ,14 ,15 ] . ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... ,15 ].这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... ,15 ]: ...
... ,
15 ,
23 ,
24 ,
25 ,
26 ]
δ18 O冷水珊瑚 -δ18 O海水 、δ13 C冷水珊瑚 -δ13 C海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ13 C冷水珊瑚 -δ13 C海水 =0时的δ18 O轴 ...
... δ
18 O
冷水珊瑚 -δ
18 O
海水 、δ
13 C
冷水珊瑚 -δ
13 C
海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时的δ
18 O轴
δ<sup>13</sup>C versus δ<sup>18</sup>O in bamboo corals<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 Ocold-water coral -δ18 Oseawater ,δ13 Ccold-water cora -δ13 Cseawater : δ18 O and δ13 C values are adjusted for local δ18 Oseawater and δ13 Cseawater values; The orange dashed line indicates δ18 O axis at δ13 Ccold-water coral -δ13 Cseawater =0 ...
... δ
18 O
cold-water coral -δ
18 O
seawater ,δ
13 C
cold-water cora -δ
13 C
seawater : δ
18 O and δ
13 C values are adjusted for local δ
18 O
seawater and δ
13 C
seawater values; The orange dashed line indicates δ
18 O axis at δ
13 C
cold-water coral -δ
13 C
seawater =0
Fig.1 ![]()
图2 <strong>δ<sup>18</sup>O</strong>截距值与温度之间的拟合关系<sup>[<xref ref-type="bibr" rid="R13">13</xref>,<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>]</sup> (a)石珊瑚δ18 O与温度间的拟合关系;(b)竹节柳珊瑚δ18 O与温度间的拟合关系 ...
... (a)石珊瑚δ
18 O与温度间的拟合关系;(b)竹节柳珊瑚δ
18 O与温度间的拟合关系
δ<sup>18</sup>O-temperature regressions from cold-water corals with 95% confidence intervals<sup>[<xref ref-type="bibr" rid="R13">13</xref>,<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>]</sup> (a) δ18 O-temperature regression for scleractinian corals;(b) δ18 O-temperature regression for bamboo corals ...
... 石珊瑚和竹节柳珊瑚的温度—氧同位素线性拟合线近乎平行,但截距值存在差异(图2 ),这与文石和高镁方解石两种矿物的结晶分馏过程有关[13 ,14 ,15 ] .在有分支(叉)结构的竹节柳珊瑚中,分支生长轴处δ18 O与δ13 C的亏损程度比主生长轴处更大,表现出更强的分馏效应.此外,靠近冷水珊瑚轴心的生长环带也会出现同位素显著亏损现象[14 ,15 ] . ...
... ,15 ]. ...
... 利用微钻取冷水珊瑚粉末样时,通常有2种方式:一种沿切面径向自中心向边缘采样,另一种沿同心生长纹层采样,这2种取样方法获得的冷水珊瑚氧、碳稳定同位素都表现出线性关系[13 ,14 ,15 ] .利用截距法计算古水温时,前一种取样方式计算出的是冷水珊瑚生长周期内周围海水的平均水温,即几十年或几百年的平均温度;后者代表冷水珊瑚某一生长时间(段)内的古水温.因此,理论上沿着珊瑚不同生长纹层取样可以获得时间序列上的古水温变化.在实际操作中,由于冷水珊瑚单个生长环带仅有约10 μm宽[28 ] ,靠肉眼难以辨认,此外还受切面大小、微钻精度的影响,取到的样品多会横跨几个环带.一些较小的珊瑚样品取样数量也会受到限制,难以获得足够的样品数据建立起氧、碳同位素的可靠线性关系.因此,利用冷水珊瑚重建古水温演化序列的误差较大. ...
Oxygen and carbon isotopic growth records in a reef coral from Florida Keys and a deep-sea coral from Blake Plateau
4
1978
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... [16 ,17 ,18 ,19 ].相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ],经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
13 C and 18 O isotopic disequilibrium in biological carbonates: II. In vitro simulation of kinetic isotope effects
5
1989
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
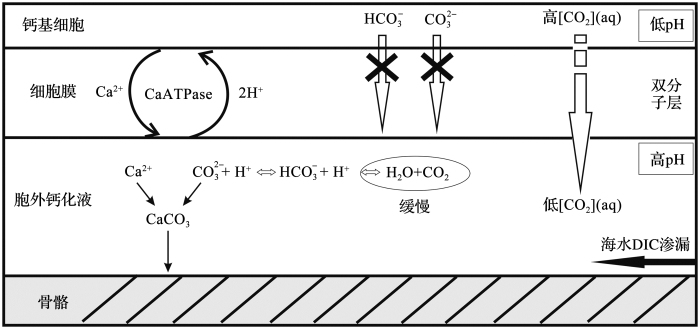
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
... [
17 ,
18 ]
骨骼与细胞膜之间存在钙化空间,碳酸钙在胞外钙化液中生成 ...
... 骨骼与细胞膜之间存在钙化空间,碳酸钙在胞外钙化液中生成
pH model schematic diagram of cold-water coral calcifying processes<sup>[<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R18">18</xref>]</sup> There is a calcification space between coral skeletons and the cell membrane, and calcium carbonate is generated ...
Stable isotopes in deep-sea corals and a new mechanism for ‘vital effects’
7
2003
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... ,18 ,19 ].相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
... ,
18 ]
骨骼与细胞膜之间存在钙化空间,碳酸钙在胞外钙化液中生成 ...
... 骨骼与细胞膜之间存在钙化空间,碳酸钙在胞外钙化液中生成
pH model schematic diagram of cold-water coral calcifying processes<sup>[<xref ref-type="bibr" rid="R17">17</xref>,<xref ref-type="bibr" rid="R18">18</xref>]</sup> There is a calcification space between coral skeletons and the cell membrane, and calcium carbonate is generated ...
... 碳酸盐从溶解无机碳中结晶之前与水保持着同位素平衡.钙化过程发生在弱碱性的胞外钙化液中,CO2 通过扩散作用进入其中,而HCO3 - 3 2 - [18 ] (图4 ).胞外钙化液中的HCO3 - 2 与H2 O或OH- 2 与H2 O反应(水合反应)形成的HCO3 - 18 OPDB 为-3.2‰;由CO2 与OH- 反应(羟基化反应)形成的HCO3 - 18 OPDB 为-15.8‰.2个反应生成HCO3 - 3 - 3 - 3 - [20 ] .两种情况下,HCO3 - 18 OPDB 分别为-12.8‰和-15.5‰.ECF中升高的pH不仅导致溶液δ18 O偏负,同时使CO2 扩散效应增强,溶液中δ13 C也出现负偏,于是生成的珊瑚骨骼δ18 O和δ13 C均负偏. ...
The geochemistry of deep-sea coral skeletons: A review of vital effects and applications for paleoceanography
3
2014
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... [19 ],增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
In situ measurements of oxygen isotopic composition in deep-sea coral, Lophelia pertusa : Re-examination of the current geochemical models of biomineralization
6
2010
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... 碳酸盐从溶解无机碳中结晶之前与水保持着同位素平衡.钙化过程发生在弱碱性的胞外钙化液中,CO2 通过扩散作用进入其中,而HCO3 - 3 2 - [18 ] (图4 ).胞外钙化液中的HCO3 - 2 与H2 O或OH- 2 与H2 O反应(水合反应)形成的HCO3 - 18 OPDB 为-3.2‰;由CO2 与OH- 反应(羟基化反应)形成的HCO3 - 18 OPDB 为-15.8‰.2个反应生成HCO3 - 3 - 3 - 3 - [20 ] .两种情况下,HCO3 - 18 OPDB 分别为-12.8‰和-15.5‰.ECF中升高的pH不仅导致溶液δ18 O偏负,同时使CO2 扩散效应增强,溶液中δ13 C也出现负偏,于是生成的珊瑚骨骼δ18 O和δ13 C均负偏. ...
... 不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
... [
20 ]
EMZ表示“早期钙化区”; ACC表示“无定形碳酸钙” ...
... EMZ表示“早期钙化区”; ACC表示“无定形碳酸钙”
ACC model of cold-water coral calcification<sup>[<xref ref-type="bibr" rid="R20">20</xref>]</sup> EMZ: Early Mineralization Zone; ACC: Amorphous Calcium Carbonates ...
Skeletal microstructure and stable isotope signature of three bathyal solitary cold-water corals from the Azores
2
2013
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
Potential climate signals from the deep-sea gorgonian coral Primnoa resedaeformis
3
2002
... 生物自身的钙化过程会造成地球化学信号偏离热力学平衡值,这种偏差被称为“生命效应”[16 ,18 ] ,不同于有孔虫和浅水珊瑚,冷水珊瑚表现出显著的生命效应[16 ,17 ,18 ,19 ] .相对碳酸盐同位素分馏平衡的理论值,冷水珊瑚的δ18 O和δ13 C呈现显著亏损,一般情况下,δ13 C亏损约10‰,δ18 O亏损4‰~6‰[20 ,21 ,22 ] .与上层海水环境变化大相比,中—深层海水环境相对稳定,季节和年际波动幅度微弱,因此生命效应在冷水珊瑚地球化学信号变化中更加凸显[19 ] ,增加了冷水珊瑚在古环境重建应用中的难度. ...
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... ,22 ,23 ],经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
Scale dependence of environmental and physiological correlates of δ18 O and δ13 C in the magnesium calcite skeletons of bamboo corals (Gorgonacea; Isididae )
3
2016
... 冷水珊瑚表现出强烈的生命效应,δ18 O与水温之间没有直接的相关关系[16 ,22 ] .尽管如此,由于冷水珊瑚δ18 O亏损的同时常常伴随着δ13 C的亏损[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ18 O与δ13 C之间存在着线性关系[13 ,14 ,15 ] .这种线性关系会在δ18 O轴上形成截距(图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ13 C冷水珊瑚 -δ13 C海水 =0时δ18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(T ,单位:℃)—氧同位素截距(δ18 O截距 )线性关系分别如下[13 ,15 ] : ...
... ,
23 ,
24 ,
25 ,
26 ]
δ18 O冷水珊瑚 -δ18 O海水 、δ13 C冷水珊瑚 -δ13 C海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ13 C冷水珊瑚 -δ13 C海水 =0时的δ18 O轴 ...
... δ
18 O
冷水珊瑚 -δ
18 O
海水 、δ
13 C
冷水珊瑚 -δ
13 C
海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时的δ
18 O轴
δ<sup>13</sup>C versus δ<sup>18</sup>O in bamboo corals<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 Ocold-water coral -δ18 Oseawater ,δ13 Ccold-water cora -δ13 Cseawater : δ18 O and δ13 C values are adjusted for local δ18 Oseawater and δ13 Cseawater values; The orange dashed line indicates δ18 O axis at δ13 Ccold-water coral -δ13 Cseawater =0 ...
Microanalysis of C and O isotopes of azooxanthellate and zooxanthellate corals by ion microprobe
3
2003
... 冷水珊瑚表现出强烈的生命效应,δ
18 O与水温之间没有直接的相关关系
[16 ,22 ] .尽管如此,由于冷水珊瑚δ
18 O亏损的同时常常伴随着δ
13 C的亏损
[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ
18 O与δ
13 C之间存在着线性关系
[13 ,14 ,15 ] .这种线性关系会在δ
18 O轴上形成截距(
图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时δ
18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(
图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(
T ,单位:℃)—氧同位素截距(δ
18 O
截距 )线性关系分别如下
[13 ,15 ] :
图1 竹节柳珊瑚氧、碳同位素关系图<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 O冷水珊瑚 -δ18 O海水 、δ13 C冷水珊瑚 -δ13 C海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ13 C冷水珊瑚 -δ13 C海水 =0时的δ18 O轴 ...
... δ
18 O
冷水珊瑚 -δ
18 O
海水 、δ
13 C
冷水珊瑚 -δ
13 C
海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时的δ
18 O轴
δ<sup>13</sup>C versus δ<sup>18</sup>O in bamboo corals<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 Ocold-water coral -δ18 Oseawater ,δ13 Ccold-water cora -δ13 Cseawater : δ18 O and δ13 C values are adjusted for local δ18 Oseawater and δ13 Cseawater values; The orange dashed line indicates δ18 O axis at δ13 Ccold-water coral -δ13 Cseawater =0 ...
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
Preboreal onset of cold-water coral growth beyond the Arctic Circle revealed by coupled radiocarbon and U-series dating and neodymium isotopes
2
2012
... 冷水珊瑚表现出强烈的生命效应,δ
18 O与水温之间没有直接的相关关系
[16 ,22 ] .尽管如此,由于冷水珊瑚δ
18 O亏损的同时常常伴随着δ
13 C的亏损
[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ
18 O与δ
13 C之间存在着线性关系
[13 ,14 ,15 ] .这种线性关系会在δ
18 O轴上形成截距(
图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时δ
18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(
图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(
T ,单位:℃)—氧同位素截距(δ
18 O
截距 )线性关系分别如下
[13 ,15 ] :
图1 竹节柳珊瑚氧、碳同位素关系图<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 O冷水珊瑚 -δ18 O海水 、δ13 C冷水珊瑚 -δ13 C海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ13 C冷水珊瑚 -δ13 C海水 =0时的δ18 O轴 ...
... δ
18 O
冷水珊瑚 -δ
18 O
海水 、δ
13 C
冷水珊瑚 -δ
13 C
海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时的δ
18 O轴
δ<sup>13</sup>C versus δ<sup>18</sup>O in bamboo corals<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 Ocold-water coral -δ18 Oseawater ,δ13 Ccold-water cora -δ13 Cseawater : δ18 O and δ13 C values are adjusted for local δ18 Oseawater and δ13 Cseawater values; The orange dashed line indicates δ18 O axis at δ13 Ccold-water coral -δ13 Cseawater =0 ...
Optical observations and geochemical data in deep-sea Hexa- and Octo-Coralla Specimens
2
2017
... 冷水珊瑚表现出强烈的生命效应,δ
18 O与水温之间没有直接的相关关系
[16 ,22 ] .尽管如此,由于冷水珊瑚δ
18 O亏损的同时常常伴随着δ
13 C的亏损
[13 ,14 ,15 ,16 ,17 ,18 ,19 ,20 ,21 ,22 ,23 ] ,经海水同位素校正后,δ
18 O与δ
13 C之间存在着线性关系
[13 ,14 ,15 ] .这种线性关系会在δ
18 O轴上形成截距(
图1 ),取自不同水温中的冷水珊瑚样品会形成一系列不同截距值,这些截距值(一般取δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时δ
18 O轴上形成的截距值)与现代海水温度之间具有线性拟合关系(
图2 ),因此利用这种拟合关系可以恢复古水温变化,该方法被称为“截距法”.石珊瑚[公式(1)]和竹节柳珊瑚[公式(2)]的温度(
T ,单位:℃)—氧同位素截距(δ
18 O
截距 )线性关系分别如下
[13 ,15 ] :
图1 竹节柳珊瑚氧、碳同位素关系图<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 O冷水珊瑚 -δ18 O海水 、δ13 C冷水珊瑚 -δ13 C海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ13 C冷水珊瑚 -δ13 C海水 =0时的δ18 O轴 ...
... δ
18 O
冷水珊瑚 -δ
18 O
海水 、δ
13 C
冷水珊瑚 -δ
13 C
海水 代表珊瑚氧、碳同位素与海水同位素的差值,橘黄色虚线表示δ
13 C
冷水珊瑚 -δ
13 C
海水 =0时的δ
18 O轴
δ<sup>13</sup>C versus δ<sup>18</sup>O in bamboo corals<sup>[<xref ref-type="bibr" rid="R14">14</xref>,<xref ref-type="bibr" rid="R15">15</xref>,<xref ref-type="bibr" rid="R23">23</xref>,<xref ref-type="bibr" rid="R24">24</xref>,<xref ref-type="bibr" rid="R25">25</xref>,<xref ref-type="bibr" rid="R26">26</xref>]</sup> δ18 Ocold-water coral -δ18 Oseawater ,δ13 Ccold-water cora -δ13 Cseawater : δ18 O and δ13 C values are adjusted for local δ18 Oseawater and δ13 Cseawater values; The orange dashed line indicates δ18 O axis at δ13 Ccold-water coral -δ13 Cseawater =0 ...
Carbon and oxygen isotopic fractionation in biogenic aragonite-temp effects
1
1986
... 其中石珊瑚温度校正方程与文石质有孔虫Hoeglundina elegans 、腹足类及掘足类生物的相关方程非常接近,它们的校正方程如下[27 ] : ...
Linear correlations in bamboo coral δ13 C and δ18 O sampled by SIMS and micromill: Evaluating paleoceanographic potential and biomineralization mechanisms using δ11 B and Δ47 composition
4
2017
... 利用微钻取冷水珊瑚粉末样时,通常有2种方式:一种沿切面径向自中心向边缘采样,另一种沿同心生长纹层采样,这2种取样方法获得的冷水珊瑚氧、碳稳定同位素都表现出线性关系[13 ,14 ,15 ] .利用截距法计算古水温时,前一种取样方式计算出的是冷水珊瑚生长周期内周围海水的平均水温,即几十年或几百年的平均温度;后者代表冷水珊瑚某一生长时间(段)内的古水温.因此,理论上沿着珊瑚不同生长纹层取样可以获得时间序列上的古水温变化.在实际操作中,由于冷水珊瑚单个生长环带仅有约10 μm宽[28 ] ,靠肉眼难以辨认,此外还受切面大小、微钻精度的影响,取到的样品多会横跨几个环带.一些较小的珊瑚样品取样数量也会受到限制,难以获得足够的样品数据建立起氧、碳同位素的可靠线性关系.因此,利用冷水珊瑚重建古水温演化序列的误差较大. ...
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
... [28 ].因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
... 同位素温度计方程提供了另外一种重建古水温的方法[33 ] .在非生物成因的碳酸盐结晶实验中,方解石与水之间的氧、碳同位素差值是温度、pH和结晶速率的函数.给定不同的pH值和碳酸盐结晶速率,可以得到Δ18 O和Δ13 C与温度之间的方程[34 ,35 ,36 ] ,Δ18 O和Δ13 C分别表示δ18 O方解石 -δ18 O水 和δ13 C方解石 -δ13 C水 (ΔA-B =δA -δB ,ΔA-B 称同位素富集系数,A和B可表示2种物质或者同一种物质的2种相态)[28 ,33 ] ,如前所述,Δ18 O和Δ13 C之间也满足线性关系. ...
1 021
1
2005
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
Radiocarbon‐based ages and growth rates of bamboo corals from the Gulf of Alaska
1
2005
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
Varying growth rates in bamboo corals: Sclerochronology and radiocarbon dating of amid‐Holocene deep‐water gorgonian skeleton (Keratoisis sp.: Octocorallia ) from Chatham Rise (New Zealand)
1
2008
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
1 005
1
2005
... 微钻取样的分辨率可达约150 μm,通常冷水珊瑚的生长速率在每年几微米到每年几百微米之间[14 ,28 ,29 ,30 ,31 ] ,因此利用生长纹层采样法,恢复出的古水温序列具有年际—年代际的时间分辨率.若要获取更高分辨率的数据,需要借助二次离子质谱(Secondary Ion Mass Spectroscopy, SIMS),其测试空间分辨率可达几十微米[24 ,32 ] ,对应的时间分辨率可达年际尺度.尽管如此,现有数据表明,SIMS分析方法的古温度重建结果误差偏大,一般微钻取样的古水温重建误差小于2 ℃,在线性关系显著的情况下,误差甚至小于1 ℃,但SIMS测量误差一般大于2 ℃[28 ] .因此,利用截距法重建古海水温度,微钻取样通常是最有效可行的手段. ...
A refined method for calculating paleotemperatures from linear correlations in bamboo coral carbon and oxygen isotopes
5
2016
... 同位素温度计方程提供了另外一种重建古水温的方法[33 ] .在非生物成因的碳酸盐结晶实验中,方解石与水之间的氧、碳同位素差值是温度、pH和结晶速率的函数.给定不同的pH值和碳酸盐结晶速率,可以得到Δ18 O和Δ13 C与温度之间的方程[34 ,35 ,36 ] ,Δ18 O和Δ13 C分别表示δ18 O方解石 -δ18 O水 和δ13 C方解石 -δ13 C水 (ΔA-B =δA -δB ,ΔA-B 称同位素富集系数,A和B可表示2种物质或者同一种物质的2种相态)[28 ,33 ] ,如前所述,Δ18 O和Δ13 C之间也满足线性关系. ...
... ,33 ],如前所述,Δ18 O和Δ13 C之间也满足线性关系. ...
... 如在珊瑚生长速率(R )为12~160 μm/a [lgR =-7.3 mol/(m2 · s)]、水体pH=8时,Δ13 C-温度[公式(4),温度单位:K]与Δ18 O-温度[公式(5)]的方程为[33 ] : ...
... 虽然Δ13 Caeq 和Δ18 Oaeq 本身都是温度的函数,但Δ18 Oaeq 对温度的敏感性比Δ13 Caeq 大1个数量级[33 ] .因此可以将Δ13 Caeq 近似看成定值,用Δ18 Oaeq 变化反映温度的变化.实际应用中,将现代观测的海水温度代入公式(4)中计算Δ13 Caeq ,通过Δ18 O和Δ13 C间的线性关系得出Δ18 Oaeq ,进而反推冷水珊瑚生长时周围海水的古温度.但这种方法计算误差较大,通常情况下为±(2~3)℃.总体来说,该方法本身存在逻辑假设上的缺陷,但可以依据水体环境参数和珊瑚生长速率,定量地计算出古温度,摆脱了经验校正关系的局限. ...
... 碳酸盐样品的二元同位素(Δ47 ,也称耦合同位素)也广泛应用于古水温重建,由于计算时不需要知晓周围水体的同位素成分,且二元同位素的变化与温度变化直接相关,因此是一种很有应用前景的古温度指标[9 ,33 ] .对于碳酸盐样品来说,碳酸根中的轻同位素会被重同位素替换,如12 C会被替换成13 C,16 O会被替换为17 O和18 O,使得CO3 2 - 表1 )[37 ] .在热力学平衡状态下,碳酸盐矿物晶格中的13 C和18 O倾向于进入同一基团中,形成13 C-18 O键,主要存在于13 C18 O16 O2 2 - 13 C-18 O键的丰度是碳酸盐形成时温度的函数,而与周围水体的同位素组成无关.实验研究中,碳酸钙与磷酸反应生成CO2 ,主要根据生成的13 C18 O16 O含量评估碳酸根中13 C-18 O键的丰度.碳酸钙中形成13 C18 O16 O2 2 - [9 ] : ...
The influence of kinetics on the oxygen isotope composition of calcium carbonate
1
2013
... 同位素温度计方程提供了另外一种重建古水温的方法[33 ] .在非生物成因的碳酸盐结晶实验中,方解石与水之间的氧、碳同位素差值是温度、pH和结晶速率的函数.给定不同的pH值和碳酸盐结晶速率,可以得到Δ18 O和Δ13 C与温度之间的方程[34 ,35 ,36 ] ,Δ18 O和Δ13 C分别表示δ18 O方解石 -δ18 O水 和δ13 C方解石 -δ13 C水 (ΔA-B =δA -δB ,ΔA-B 称同位素富集系数,A和B可表示2种物质或者同一种物质的2种相态)[28 ,33 ] ,如前所述,Δ18 O和Δ13 C之间也满足线性关系. ...
The influence of temperature, pH, and growth rate on the δ18 O composition of inorganically precipitated calcite
1
2014
... 同位素温度计方程提供了另外一种重建古水温的方法[33 ] .在非生物成因的碳酸盐结晶实验中,方解石与水之间的氧、碳同位素差值是温度、pH和结晶速率的函数.给定不同的pH值和碳酸盐结晶速率,可以得到Δ18 O和Δ13 C与温度之间的方程[34 ,35 ,36 ] ,Δ18 O和Δ13 C分别表示δ18 O方解石 -δ18 O水 和δ13 C方解石 -δ13 C水 (ΔA-B =δA -δB ,ΔA-B 称同位素富集系数,A和B可表示2种物质或者同一种物质的2种相态)[28 ,33 ] ,如前所述,Δ18 O和Δ13 C之间也满足线性关系. ...
A process-based model for nonequilibrium clumped isotope effects in carbonates
1
2015
... 同位素温度计方程提供了另外一种重建古水温的方法[33 ] .在非生物成因的碳酸盐结晶实验中,方解石与水之间的氧、碳同位素差值是温度、pH和结晶速率的函数.给定不同的pH值和碳酸盐结晶速率,可以得到Δ18 O和Δ13 C与温度之间的方程[34 ,35 ,36 ] ,Δ18 O和Δ13 C分别表示δ18 O方解石 -δ18 O水 和δ13 C方解石 -δ13 C水 (ΔA-B =δA -δB ,ΔA-B 称同位素富集系数,A和B可表示2种物质或者同一种物质的2种相态)[28 ,33 ] ,如前所述,Δ18 O和Δ13 C之间也满足线性关系. ...
13 C-18 O bonds in carbonate minerals: A new kind of paleothermometer
4
2006
... 碳酸盐样品的二元同位素(Δ47 ,也称耦合同位素)也广泛应用于古水温重建,由于计算时不需要知晓周围水体的同位素成分,且二元同位素的变化与温度变化直接相关,因此是一种很有应用前景的古温度指标[9 ,33 ] .对于碳酸盐样品来说,碳酸根中的轻同位素会被重同位素替换,如12 C会被替换成13 C,16 O会被替换为17 O和18 O,使得CO3 2 - 表1 )[37 ] .在热力学平衡状态下,碳酸盐矿物晶格中的13 C和18 O倾向于进入同一基团中,形成13 C-18 O键,主要存在于13 C18 O16 O2 2 - 13 C-18 O键的丰度是碳酸盐形成时温度的函数,而与周围水体的同位素组成无关.实验研究中,碳酸钙与磷酸反应生成CO2 ,主要根据生成的13 C18 O16 O含量评估碳酸根中13 C-18 O键的丰度.碳酸钙中形成13 C18 O16 O2 2 - [9 ] : ...
... 同位素随机分布时CO3 和CO2 各同位素体的丰度[37 ] ...
... Abundances of isotopologues of CO3 and2 ,assuming a stochastic distribution of isotopes [37 ] ...
... Δ47 的取值直接受到温度的影响,但目前还没有公认的利用Δ47 计算温度的公式,主要是建立两者间的经验关系.Ghosh等[37 ] 率先通过合成方解石实验建立了Δ47 与温度(单位:K)间的拟合方程[公式(8)]: ...
“Clumped-isotope” geochemistry—The study of naturally-occurring, multiply-substituted isotopologues
1
262
... Δ47 的定义为碳酸钙酸解生成的质量数为47的CO2 (主要是13 C18 O16 O)分子丰度相对于随机分布时该分子丰度的差异程度,用以表示13 C-18 O键的聚合程度,单位‰,计算公式如下[公式(7)][38 ] : ...
Schrag D P. Clumped isotope thermometry of carbonatites as an indicator of diagenetic alteration
1
2010
... 之后不同的经验公式接踵出现[39 ,40 ,41 ] ,有孔虫、腕足类、双壳类等生物成因碳酸盐的Δ47 投点结果更接近Ghosh的校正方程[40 ,41 ] .但浅水珊瑚和某些属种冷水珊瑚的二元同位素都展现出显著的生命效应,如在冷水珊瑚Balanophyllia , Caryophyllia , Dasmosmilia ,Desmophyllum ,Enallopsammia 和Javania 等属种中[9 ] ,Δ47 比Ghosh校正方程计算的同等温度下的平衡值偏高,一些样品甚至高出约0.05‰(相当于温度被低估了约9 ℃).因此,使用二元同位素Δ47 重建古水温时也需要对冷水珊瑚属种进行生命效应校正. ...
Carbonate clumped isotope thermometry of deep-sea corals and implications for vital effects
2
2011
... 之后不同的经验公式接踵出现[39 ,40 ,41 ] ,有孔虫、腕足类、双壳类等生物成因碳酸盐的Δ47 投点结果更接近Ghosh的校正方程[40 ,41 ] .但浅水珊瑚和某些属种冷水珊瑚的二元同位素都展现出显著的生命效应,如在冷水珊瑚Balanophyllia , Caryophyllia , Dasmosmilia ,Desmophyllum ,Enallopsammia 和Javania 等属种中[9 ] ,Δ47 比Ghosh校正方程计算的同等温度下的平衡值偏高,一些样品甚至高出约0.05‰(相当于温度被低估了约9 ℃).因此,使用二元同位素Δ47 重建古水温时也需要对冷水珊瑚属种进行生命效应校正. ...
... [40 ,41 ].但浅水珊瑚和某些属种冷水珊瑚的二元同位素都展现出显著的生命效应,如在冷水珊瑚Balanophyllia , Caryophyllia , Dasmosmilia ,Desmophyllum ,Enallopsammia 和Javania 等属种中[9 ] ,Δ47 比Ghosh校正方程计算的同等温度下的平衡值偏高,一些样品甚至高出约0.05‰(相当于温度被低估了约9 ℃).因此,使用二元同位素Δ47 重建古水温时也需要对冷水珊瑚属种进行生命效应校正. ...
Paleo-environmental implication of clumped isotopes in land snail shells
2
2011
... 之后不同的经验公式接踵出现[39 ,40 ,41 ] ,有孔虫、腕足类、双壳类等生物成因碳酸盐的Δ47 投点结果更接近Ghosh的校正方程[40 ,41 ] .但浅水珊瑚和某些属种冷水珊瑚的二元同位素都展现出显著的生命效应,如在冷水珊瑚Balanophyllia , Caryophyllia , Dasmosmilia ,Desmophyllum ,Enallopsammia 和Javania 等属种中[9 ] ,Δ47 比Ghosh校正方程计算的同等温度下的平衡值偏高,一些样品甚至高出约0.05‰(相当于温度被低估了约9 ℃).因此,使用二元同位素Δ47 重建古水温时也需要对冷水珊瑚属种进行生命效应校正. ...
... ,41 ].但浅水珊瑚和某些属种冷水珊瑚的二元同位素都展现出显著的生命效应,如在冷水珊瑚Balanophyllia , Caryophyllia , Dasmosmilia ,Desmophyllum ,Enallopsammia 和Javania 等属种中[9 ] ,Δ47 比Ghosh校正方程计算的同等温度下的平衡值偏高,一些样品甚至高出约0.05‰(相当于温度被低估了约9 ℃).因此,使用二元同位素Δ47 重建古水温时也需要对冷水珊瑚属种进行生命效应校正. ...
71
1
2007
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
Sub-equilibrium oxygen-18 and carbon-13 levels in biological carbonates: Carbonate and kinetic models
1
2003
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
An explanation of the effect of seawater carbonate concentration on foraminiferal oxygen isotopes
1
63
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
An expression for the overall oxygen isotope fractionation between the sum of dissolved inorganic carbon and water
1
2007
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
Effect of seawater carbonate ion concentration on foraminiferal carbon and oxygen isotopes
1
1997
... 第一种可称为“pH驱动模型”,该模型认为冷水珊瑚δ18 O的偏负与胞外钙化液中pH值的变化有关.假想珊瑚骨骼是在钙化液体库中形成的(图3 ),该液体库被称为胞外钙化液(Extracytoplasmic Calcifying Fluid, ECF),海水能够通过一些通道补充ECF[17 ,18 ,42 ] .Ca2+ 和H+ 通过细胞膜上的Ca-ATPase(Ca2+ -ATP酶通道蛋白)进出ECF中[43 ] ,Ca-ATPase每次从ECF中运出2个质子,同时从钙基细胞内泵入1个Ca2+ ,使得ECF中pH值升高.不同的pH造成ECF中H2 CO3 ,HCO3 - 3 2 - 18 O值[44 ,45 ] .因此,新形成的珊瑚骨骼δ18 O也会随之变化.在有孔虫中,pH值的增加导致壳体δ18 O降低[46 ] ,冷水珊瑚中也应存在类似效应. ...
Carbon isotopic fractionation in synthetic aragonite and calcite: Effects of temperature and precipitation rate
1
1992
... 冷水珊瑚δ13 C的偏负与钙基细胞中的HCO3 - 3 2 - 2 是3种粒子中唯一可以被动透过细胞膜的分子.Ca-ATPase产生的Ca泵效应造成细胞膜两侧存在较大的p CO2 和pH梯度,从而驱动着CO2 进入到ECF中.由于海水可以补充到ECF中,因此形成冷水珊瑚骨骼的碳有2个来源:钙基细胞和海水.当Ca泵效率较高时,钙化速率也升高,随着CaCO3 的生成,钙化母液不能及时供应生成珊瑚骨骼所利用的碳,此时形成的骨骼δ13 C会非常接近补充到ECF中碳的同位素值,该值取决于ECF中钙基细胞和海水的比例.在5 ℃时,热力学平衡状态下,CO2 相对于文石的δ13 C偏负约13‰,海水的碳同位素通常在-1‰~1‰[47 ,48 ] .海水渗漏程度取决于钙化时的地球化学条件.因此,当来自钙基细胞的CO2 通量达最大值时,冷水珊瑚骨骼δ13 C最负;海水补充到ECF中,使δ13 C负偏程度减小. ...
Carbon isotope fractionation during gas-water exchange and dissolution of CO2
1
1995
... 冷水珊瑚δ13 C的偏负与钙基细胞中的HCO3 - 3 2 - 2 是3种粒子中唯一可以被动透过细胞膜的分子.Ca-ATPase产生的Ca泵效应造成细胞膜两侧存在较大的p CO2 和pH梯度,从而驱动着CO2 进入到ECF中.由于海水可以补充到ECF中,因此形成冷水珊瑚骨骼的碳有2个来源:钙基细胞和海水.当Ca泵效率较高时,钙化速率也升高,随着CaCO3 的生成,钙化母液不能及时供应生成珊瑚骨骼所利用的碳,此时形成的骨骼δ13 C会非常接近补充到ECF中碳的同位素值,该值取决于ECF中钙基细胞和海水的比例.在5 ℃时,热力学平衡状态下,CO2 相对于文石的δ13 C偏负约13‰,海水的碳同位素通常在-1‰~1‰[47 ,48 ] .海水渗漏程度取决于钙化时的地球化学条件.因此,当来自钙基细胞的CO2 通量达最大值时,冷水珊瑚骨骼δ13 C最负;海水补充到ECF中,使δ13 C负偏程度减小. ...
Microscopic and systematic study of madreporarian type of corals
1
895
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
Microstructural and physicochemical characterization of “centers of calcification” in septa of some recent scleractinian corals
1
1998
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
Vital effects in coral skeletal composition display strict three-dimensional control
1
2006
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
Compositional variations at ultra-structure length scales in coral skeleton
1
2008
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
A numerical model of trace element coprecpitation in a physicochemical calcification system: Application to coral biomineralization and trace element ‘vital effects’
1
2006
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
Effect of environmental conditions on the Li isotopic composition of scleractinian corals
1
286
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
The Sr/Ca temperature relationship incoralline aragonite: Influence of variability in (Sr/Ca) seawater and skeletal growth parameters
1
1994
... 第二种,“动力学分馏模型”认为冷水珊瑚δ18 O和δ13 C的异常偏负现象与pH驱动的同位素动力学分馏效应有关.石珊瑚的微结构中存在着一些球粒状的“早期钙化区”(Early Mineralization Zone, EMZ),也被称为钙化中心,EMZ之外的区域是束状堆积的纤维层[49 ,50 ,51 ,52 ,53 ] .两种结构间存在成分差异,具体表现为EMZ内δ18 O、δ13 C、δ11 B及Sr/Ca偏负,而Mg/Ca偏高[54 ,55 ] .相比于周围的纤维层,EMZ内偏负的δ18 O和δ13 C反映了较强的动力学分馏效应. ...
Observations of the tissue-skeleton interface in the scleractinian coral Stylophora pistillata
1
2007
... 不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
Calcium associated with a fibrillar organic matrix in the scleractinian coral Galaxea fascicularis
2
220
... 不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
... [57 ].事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
Taking advantage of disorder: Amorphous calcium carbonate and its roles in biomineralization
1
2003
... 不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
Stable prenucleation calcium carbonate clusters
1
2008
... 不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
Nanostructure, composition and mechanisms of bivalve shell growth
1
2008
... 不同于前两种钙化模型,Rollion-Bard 等[20 ] 提出了新的简易模型框架——“无定形碳酸钙模型”.“pH驱动模型”和“动力学分馏模型”都假设冷水珊瑚骨骼与组织之间存在着钙化空间,但石珊瑚的显微结构显示两者直接接触[56 ,57 ] ,并不存在钙化空间,如果存在也仅限于几纳米的区域.“无定形碳酸钙模型”认为EMZ形成之前,无定形碳酸钙(Amorphous Calcium Carbonates, ACC)会作为先存相态出现,并被特定的大分子固定[58 ] (图5 ).软体动物贝类、棘皮类等生物,它们在建造方解石或文石质壳体时,ACC通常作为初期形态出现[59 ] .此外,ACC的结晶过程也不一定发生在类似海水成分的钙化液体中,而是发生在特定成分构成的有机水凝胶层上[57 ] .事实上在软体动物贝类中,只发现ACC富集Mg、S和P[60 ] .对冷水珊瑚还没有开展类似的分析工作,至于能否解释EMZ异常偏低的δ18 O和δ13 C值,仍需要更多地微区分析实验来验证. ...
World ocean heat content and thermosteric sea level change (0-2000m), 1955-2010
1
2012
... 冷水珊瑚早在一个多世纪之前就被发现,但直到最近20多年才被广泛地用于高分辨率的古海洋环境变化研究[61 ,62 ] .利用冷水珊瑚δ18 O-δ13 C截距恢复古水温的方法可重复性强,操作简单,目前也应用最广.在一些线性关系较好的珊瑚样品中,温度重建误差小于1 ℃,但校正数据点仍然较少,仍需要更多来自全球各大洋的活体珊瑚样品来提供更加可靠的约束条件.冷水珊瑚氧、碳同位素温度计方程可以定量化计算古水温,但结果误差较大且存在逻辑假设缺陷.相比之下,二元同位素方法前景较好,但目前仅限应用于一些未表现出生命效应的属种.同时,未来的研究需要开发配套的定年方法.比如可以借助石珊瑚样品U-Th和14 C联合定年计算出特定海区某深度范围内的碳储库年龄[63 ,64 ,65 ,66 ] ,为竹节柳珊瑚14 C测年提供储库校正,配合氧、碳同位素取样,从而重建中—深层海水温度序列变化. ...
The southern ocean's role in carbon exchange during the last deglaciation
1
2012
... 冷水珊瑚早在一个多世纪之前就被发现,但直到最近20多年才被广泛地用于高分辨率的古海洋环境变化研究[61 ,62 ] .利用冷水珊瑚δ18 O-δ13 C截距恢复古水温的方法可重复性强,操作简单,目前也应用最广.在一些线性关系较好的珊瑚样品中,温度重建误差小于1 ℃,但校正数据点仍然较少,仍需要更多来自全球各大洋的活体珊瑚样品来提供更加可靠的约束条件.冷水珊瑚氧、碳同位素温度计方程可以定量化计算古水温,但结果误差较大且存在逻辑假设缺陷.相比之下,二元同位素方法前景较好,但目前仅限应用于一些未表现出生命效应的属种.同时,未来的研究需要开发配套的定年方法.比如可以借助石珊瑚样品U-Th和14 C联合定年计算出特定海区某深度范围内的碳储库年龄[63 ,64 ,65 ,66 ] ,为竹节柳珊瑚14 C测年提供储库校正,配合氧、碳同位素取样,从而重建中—深层海水温度序列变化. ...
Synchronous centennial abrupt events in the ocean and atmosphere during the last deglaciation
1
2015
... 冷水珊瑚早在一个多世纪之前就被发现,但直到最近20多年才被广泛地用于高分辨率的古海洋环境变化研究[61 ,62 ] .利用冷水珊瑚δ18 O-δ13 C截距恢复古水温的方法可重复性强,操作简单,目前也应用最广.在一些线性关系较好的珊瑚样品中,温度重建误差小于1 ℃,但校正数据点仍然较少,仍需要更多来自全球各大洋的活体珊瑚样品来提供更加可靠的约束条件.冷水珊瑚氧、碳同位素温度计方程可以定量化计算古水温,但结果误差较大且存在逻辑假设缺陷.相比之下,二元同位素方法前景较好,但目前仅限应用于一些未表现出生命效应的属种.同时,未来的研究需要开发配套的定年方法.比如可以借助石珊瑚样品U-Th和14 C联合定年计算出特定海区某深度范围内的碳储库年龄[63 ,64 ,65 ,66 ] ,为竹节柳珊瑚14 C测年提供储库校正,配合氧、碳同位素取样,从而重建中—深层海水温度序列变化. ...
Coral provides way to age deep water
1
1998
... 冷水珊瑚早在一个多世纪之前就被发现,但直到最近20多年才被广泛地用于高分辨率的古海洋环境变化研究[61 ,62 ] .利用冷水珊瑚δ18 O-δ13 C截距恢复古水温的方法可重复性强,操作简单,目前也应用最广.在一些线性关系较好的珊瑚样品中,温度重建误差小于1 ℃,但校正数据点仍然较少,仍需要更多来自全球各大洋的活体珊瑚样品来提供更加可靠的约束条件.冷水珊瑚氧、碳同位素温度计方程可以定量化计算古水温,但结果误差较大且存在逻辑假设缺陷.相比之下,二元同位素方法前景较好,但目前仅限应用于一些未表现出生命效应的属种.同时,未来的研究需要开发配套的定年方法.比如可以借助石珊瑚样品U-Th和14 C联合定年计算出特定海区某深度范围内的碳储库年龄[63 ,64 ,65 ,66 ] ,为竹节柳珊瑚14 C测年提供储库校正,配合氧、碳同位素取样,从而重建中—深层海水温度序列变化. ...
CO2 storage and release in the deep Southern Ocean on millennial to centennial timescales
1
2018
... 冷水珊瑚早在一个多世纪之前就被发现,但直到最近20多年才被广泛地用于高分辨率的古海洋环境变化研究[61 ,62 ] .利用冷水珊瑚δ18 O-δ13 C截距恢复古水温的方法可重复性强,操作简单,目前也应用最广.在一些线性关系较好的珊瑚样品中,温度重建误差小于1 ℃,但校正数据点仍然较少,仍需要更多来自全球各大洋的活体珊瑚样品来提供更加可靠的约束条件.冷水珊瑚氧、碳同位素温度计方程可以定量化计算古水温,但结果误差较大且存在逻辑假设缺陷.相比之下,二元同位素方法前景较好,但目前仅限应用于一些未表现出生命效应的属种.同时,未来的研究需要开发配套的定年方法.比如可以借助石珊瑚样品U-Th和14 C联合定年计算出特定海区某深度范围内的碳储库年龄[63 ,64 ,65 ,66 ] ,为竹节柳珊瑚14 C测年提供储库校正,配合氧、碳同位素取样,从而重建中—深层海水温度序列变化. ...
Deep-sea coral evidence for rapid change in ventilation of the deep North Atlantic 15,400 years ago
1
1998
... 冷水珊瑚早在一个多世纪之前就被发现,但直到最近20多年才被广泛地用于高分辨率的古海洋环境变化研究[61 ,62 ] .利用冷水珊瑚δ18 O-δ13 C截距恢复古水温的方法可重复性强,操作简单,目前也应用最广.在一些线性关系较好的珊瑚样品中,温度重建误差小于1 ℃,但校正数据点仍然较少,仍需要更多来自全球各大洋的活体珊瑚样品来提供更加可靠的约束条件.冷水珊瑚氧、碳同位素温度计方程可以定量化计算古水温,但结果误差较大且存在逻辑假设缺陷.相比之下,二元同位素方法前景较好,但目前仅限应用于一些未表现出生命效应的属种.同时,未来的研究需要开发配套的定年方法.比如可以借助石珊瑚样品U-Th和14 C联合定年计算出特定海区某深度范围内的碳储库年龄[63 ,64 ,65 ,66 ] ,为竹节柳珊瑚14 C测年提供储库校正,配合氧、碳同位素取样,从而重建中—深层海水温度序列变化. ...
Correlation of boron isotopic composition with ultrastructure in the deep-sea coral Lophelia pertusa : Implications for biomineralization and paleo-pH
1
2007
... 前人提出的3种钙化模型虽然都可以解释冷水珊瑚δ18 O和δ13 C的异常偏负现象,但同一种模型用于不同的指标会出现矛盾.如“早期钙化区”中同时出现δ18 O和δ11 B偏轻,根据“pH驱动模型”的解释,δ18 O偏轻表明钙化液中pH升高,此时δ11 B应该偏重[67 ] .“动力学分馏模型”表明异常的同位素信号是由钙化液中pH的变化驱动的,若要使“早期钙化区”中δ18 O和δ13 C偏负,要求冷水珊瑚中所有形成“早期钙化区”的地方pH值最高,这也与δ11 B恢复的pH数据相矛盾[68 ,69 ] .“无定形碳酸钙模型”能否解释δ18 O和δ13 C同位素和一些元素比值的异常仍未可知.因此,今后仍需要更多的微区观察与分析工作,以及针对性的实验来揭示影响冷水珊瑚的钙化过程的因素,如ACC形成时方解石或文石中同位素的分馏系数,有机水凝胶层的作用等. ...
Effects of seawater-pH and biomineralization on the boron isotopic composition of deep-sea bamboo corals
1
2015
... 前人提出的3种钙化模型虽然都可以解释冷水珊瑚δ18 O和δ13 C的异常偏负现象,但同一种模型用于不同的指标会出现矛盾.如“早期钙化区”中同时出现δ18 O和δ11 B偏轻,根据“pH驱动模型”的解释,δ18 O偏轻表明钙化液中pH升高,此时δ11 B应该偏重[67 ] .“动力学分馏模型”表明异常的同位素信号是由钙化液中pH的变化驱动的,若要使“早期钙化区”中δ18 O和δ13 C偏负,要求冷水珊瑚中所有形成“早期钙化区”的地方pH值最高,这也与δ11 B恢复的pH数据相矛盾[68 ,69 ] .“无定形碳酸钙模型”能否解释δ18 O和δ13 C同位素和一些元素比值的异常仍未可知.因此,今后仍需要更多的微区观察与分析工作,以及针对性的实验来揭示影响冷水珊瑚的钙化过程的因素,如ACC形成时方解石或文石中同位素的分馏系数,有机水凝胶层的作用等. ...
Boron isotopes as pH proxy: A new look at boron speciation in deep-sea corals using B-11 MAS NMR and EELS
1
2011
... 前人提出的3种钙化模型虽然都可以解释冷水珊瑚δ18 O和δ13 C的异常偏负现象,但同一种模型用于不同的指标会出现矛盾.如“早期钙化区”中同时出现δ18 O和δ11 B偏轻,根据“pH驱动模型”的解释,δ18 O偏轻表明钙化液中pH升高,此时δ11 B应该偏重[67 ] .“动力学分馏模型”表明异常的同位素信号是由钙化液中pH的变化驱动的,若要使“早期钙化区”中δ18 O和δ13 C偏负,要求冷水珊瑚中所有形成“早期钙化区”的地方pH值最高,这也与δ11 B恢复的pH数据相矛盾[68 ,69 ] .“无定形碳酸钙模型”能否解释δ18 O和δ13 C同位素和一些元素比值的异常仍未可知.因此,今后仍需要更多的微区观察与分析工作,以及针对性的实验来揭示影响冷水珊瑚的钙化过程的因素,如ACC形成时方解石或文石中同位素的分馏系数,有机水凝胶层的作用等. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}