



1 引 言
2018年春天,在南海深部有个重大发现:在超过千米的深水海底发现了珊瑚林。使用国产载人深潜器“深海勇士号”和加拿大ROPOS遥控深潜器,分别对西沙的深水区和北部的海山进行探索,均在1 000~3 000 m的深海底发现了成片分布的珊瑚林(图1)[1]。先前大家熟悉的是珊瑚礁,也知道深水里有单体珊瑚,但生长在深海硬基底上的“珊瑚林”,在南海还是首次发现。虽然个别的深水珊瑚标本早就采到过,但成林分布的深水珊瑚在南海、甚至整个热带西太平洋还是首次报道,因为珊瑚林只有通过深潜才能看到。在西沙最为突出的是鞭状的竹节珊瑚Lepidisis sp.(图1中a),主竿高逾2 m,加上蜷曲的活珊瑚部分总长可达4~5 m,宛如园林里的乔木;较矮的扇珊瑚 Calyptrophora sp.(图1中b),好比林下的乔木;再配上玻璃海绵(图1中c)等更矮的动物,构成了一片不可思议的深海园林。
图1

图1
南海的深水珊瑚林
(a),(b) 西沙深海的珊瑚林 (2018年“深海勇士号”拍摄); (c) 深海盆海山上的珊瑚林(2018年“ROPOS”拍摄,周怀阳
提供); a为竹节珊瑚Lepidisis sp.; b为扇珊瑚Calyptrophora sp.; c为玻璃海绵
Fig.1
Deep coral forests in the South China Sea
(a),(b) Deep-sea coral forests in the Xisha Islands (photo by HOV “Shenhai Yongshi” in 2018); (c) Coral forests on seamounts of the deep South China Sea basin (photo by ROV “ROPOS” in 2018, provided by Zhou Huaiyang). a: Bamboo coral Lepidisis sp.; b: Fan coral Calyptrophora sp.; c: Glass sponge
与热带的造礁珊瑚不同,深水珊瑚生活在冷水里,因此也叫冷水珊瑚。但是冷水珊瑚在浅海也可以出现,因此深水珊瑚一定在冷水里,冷水珊瑚却不一定都在深水。在国际学术界,深海珊瑚研究主要于21世纪开始,但是引起研究人员注意的并不是深水珊瑚林,而是大西洋的深水珊瑚礁。本文拟对深水珊瑚做一次比较系统地介绍,先从研究历史、即北大西洋深水珊瑚的研究谈起,然后介绍深水珊瑚林,接着从生态系统和海洋生物泵的角度讨论深水珊瑚、尤其是深水珊瑚林的价值,最后介绍深水珊瑚研究的地质意义,并探讨我国的研究方向。
2 深水珊瑚礁
2.1 深海造礁的石珊瑚
自然科学的众多领域是在欧洲创立的,研究深水珊瑚的源头也在西欧。早在18世纪已经发现深水珊瑚,最主要的2种深水“造礁”珊瑚——多孔冠珊瑚Lophelia pertusa和多眼筛珊瑚Madrepora oculata,都是生物分类法的创始人——瑞典的林奈(Carl Linné)于1758年命名的。至于深水珊瑚的海上研究,也可以上溯到19世纪,1872—1876年英国“挑战者号(HMS Challenger)”进行环球科考航行,从深水海底采回来大量珊瑚标本进行描述分类,可惜对其产状一无所知。海洋底栖生物的研究受采样手段限制,直到20世纪仍局限在软基底上,即有沉积覆盖的海底[2],因为无论箱式取样器或是海底拖网都只适用于软质海底。需等到出现深潜技术,载人或者不载人深潜器下至岩石基底的深海底面,方能开拓深海珊瑚的研究领域[3],到现在已经有越来越多的深海技术可以用于研究(例如着底器lander[4])。因此,深海珊瑚的研究是21世纪海洋新技术的产物。
深水珊瑚得到重视的直接原因,是20世纪晚期的深水捕鱼和海底采油,在北大西洋发现了深水珊瑚礁体。比如1990年代在挪威中部岸外水深220~310 m处,发现点状分布的深水珊瑚礁,礁体高30 m、长500 m、宽150 m,平均每平方公里有1.2个,放射性碳测年表明大约均是8 000年以来的产物。这类深水珊瑚礁沿着挪威与爱尔兰西岸大量发育,很可能与海底油气苗相关,于是推测泄出的甲烷会转化为无机碳,有利于冷水珊瑚礁的发育[5]。类似的深水珊瑚礁在大西洋对岸的加拿大和美国西岸也有分布,水深一般不超过1 000 m,是大西洋两侧的共同现象。深水珊瑚礁主要由多孔冠珊瑚L. pertusa组成,其次有多眼筛珊瑚Madrepora oculata等种。进入21世纪,涉及这两类珊瑚的文献数量剧增,反映出深水珊瑚礁研究迅速发展的趋势(图2)[6]。
图2
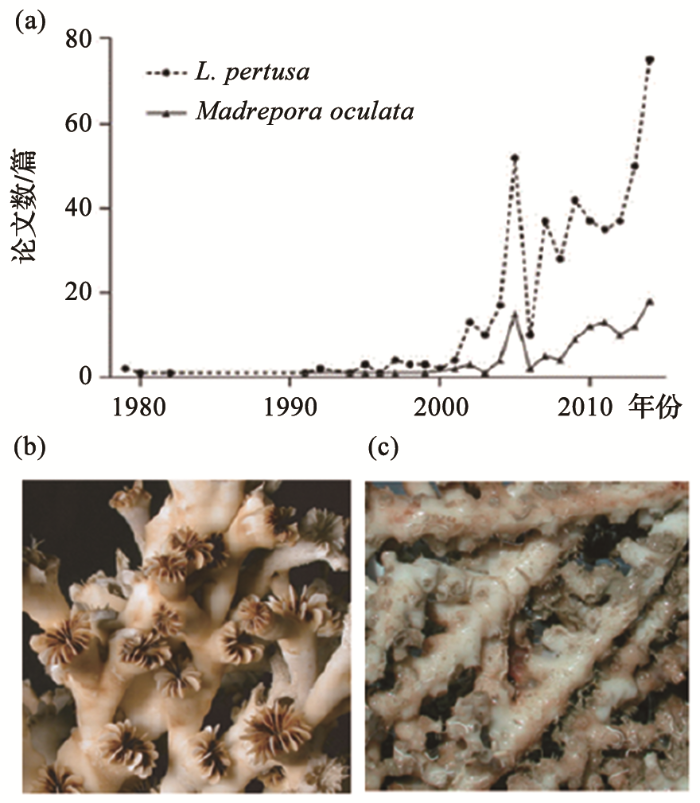
2.2 深水珊瑚礁和碳酸盐泥丘
图3
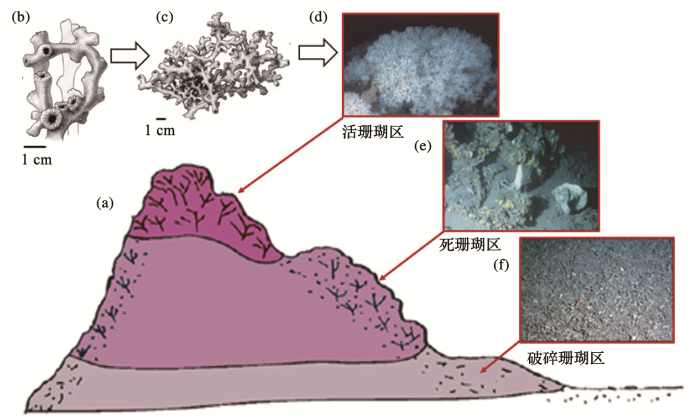
图3
一座典型的深水珊瑚礁
(a)礁体上下分三部分;(b)~(d)“活珊瑚”区(d)由多孔冠珊瑚和多眼筛珊瑚占压倒优势;b 和c分别表示多孔冠珊瑚L. pertusa的个体(b)和群体(c);(e)死珊瑚区种类最多,有活的柳珊瑚和海绵;(f)“破碎珊瑚”区多包壳型海绵[7]
Fig.3
A typical deep-sea coral reef
(a)The reef is vertically constituted by three parts;(b)~(d)The "living coral zone" (d) dominated by Lophelia pertusa and Madrepora oculata,and the individual living L. pertusa (b)interweaved into a community(c);(e)"Dead framework zone" with diverse species including living gorgonian and sponges;(f)"Coral rubble zone" occupied by encrusting sponges[7]
由此便出现了分歧。有的作者为了强调与热带珊瑚礁的区别,认为深水珊瑚形成的结构不能称为“礁”,只能叫珊瑚“丘”。不过近年来的研究表明,从生物学角度看,深水珊瑚与浅水珊瑚造成的骨架并没有根本区别,不妨统称为“珊瑚礁”;但是从沉积学角度看却大不相同,深水珊瑚形成的是一种比较松散的堆积体,虽然产生的碳酸盐可以堆到300 m高,但是与碳酸盐泥(carbonate mud)一起堆成的丘状隆起,与热带珊瑚礁致密的碳酸盐体明显不同。因此两者都没错:从生物学角度称为深水珊瑚礁,形成的地质体称为“碳酸盐泥丘(carbonate mud mound)”。
图4
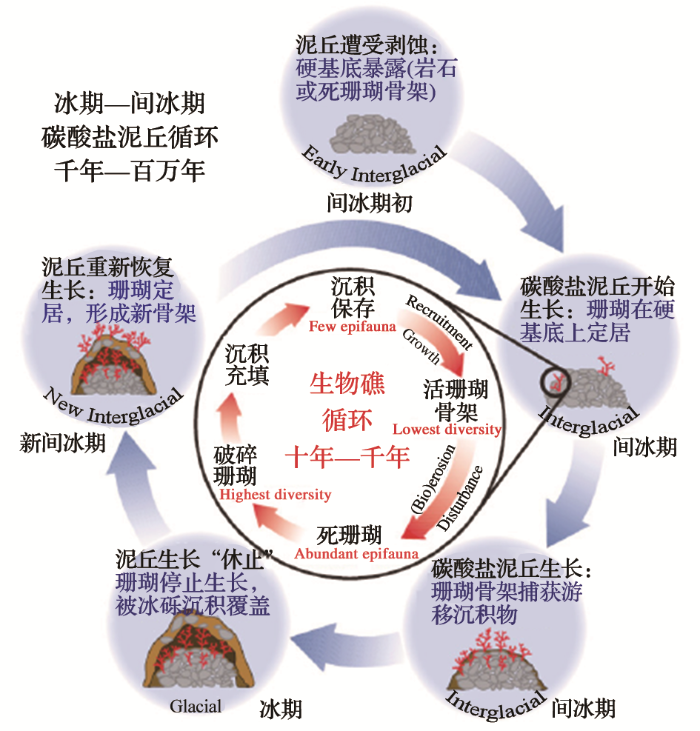
在现代的大洋里,深水碳酸盐泥丘主要分布在大西洋区(图5)[9],研究最多的是西欧岸外和美国东南一带[10]。这类沉积体规模相当大,在海底地形和地震剖面上都有表现,是第四纪地质研究的重要对象。如著名的挪威岸外200~300 m深处,沿着冰山带来的冰川砾石条带,形成了几百个2~30 m高的冷水珊瑚丘[7]。最近西非毛里塔尼亚岸外水深400~500 m处,也发现有高逾百米、延伸400 km的特大型冷水碳酸盐泥丘,这是12万年以来的碳酸盐堆积体[11]。一般来说,碳酸盐泥丘常见于200~300 m处,深度在几百米范围内,但也见于1 000~1 300 m的海山上,最大深度是2017年在爱尔兰岸外水深1 600 m处发现的。
图5
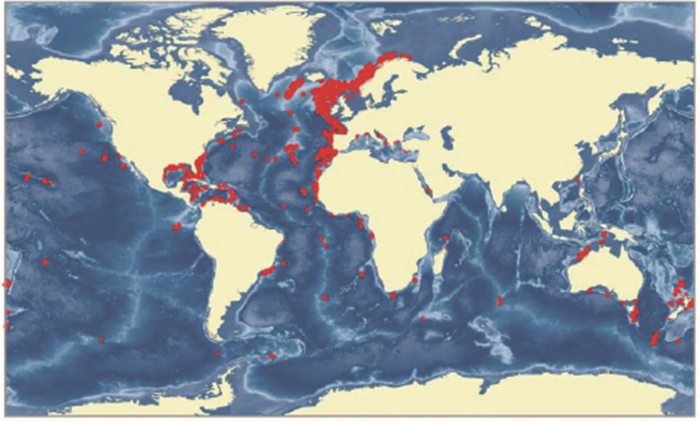
深水珊瑚在大洋碳酸盐沉积作用中起着很重要的作用,以美国东南佛罗里达海域为例,在陆坡上每3 km2就有1座深水珊瑚的丘。调查区12×104 km2就有40 000座深水珊瑚丘,以每座50 m×200 m面积计算,总面积可达400 km2,碳酸盐的总量十分可观。相比之下,热带浅水珊瑚礁只是在碳酸盐台地的边缘上发育生长,沿佛罗里达湾也不过100 km宽,这样看来“罕见”的冷水珊瑚,在美国东南比热带珊瑚礁的面积大[12]。但是这类深水珊瑚礁在太平洋海域很少见,南海发现的也不是深水珊瑚礁,而是深水珊瑚林。
3 深水珊瑚林
3.1 深海造林的软珊瑚
进入21世纪以来深水珊瑚礁的名声大噪,但南海发现的并不是深水珊瑚礁,而是珊瑚林。前文讨论的深水珊瑚礁分布在大西洋,太平洋尚未发现。深水珊瑚林与深水珊瑚礁无论是分类还是形态都不相同:大西洋造“礁”的是石珊瑚,南海造“林”的是软珊瑚,它们是两大类不同的珊瑚。
为什么大西洋有深水珊瑚礁,而太平洋只有珊瑚林?原因在于两者的碳酸盐不同。深水珊瑚林的软珊瑚骨骼属于高镁方解石,比较抗溶;深水珊瑚造“礁”的骨骼是文石,只有在文石的补偿深度以上才能出现,大西洋文石的补偿深度为2~3 km,太平洋只有0.5~1.5 km深[13],文石在太平洋比大西洋更容易溶解。
| 亚纲 | 目 | 实例 | 种数 | |
|---|---|---|---|---|
| 总数 | 水深>50 m | |||
| 六放珊瑚Hexocorallia | 石珊瑚Scleractinia | 造礁珊瑚 | 1 488 | 622 |
| 角珊瑚Antipatharia | 黑珊瑚等 | 241 | 182 | |
| 八放珊瑚Octocorellia | 软珊瑚Alcyonacea | 或称柳珊瑚,含竹节珊瑚、扇珊瑚等 | 3 159 | 2 325 |
| 珊瑚总计* | 5 160 | 3 356 | ||
分类上八放珊瑚的主体是软珊瑚目(Alcyonacea),也称为柳珊瑚目(Gorgonacea)。珊瑚的分类可能有点混乱,比如上面提到的黑珊瑚现在归为六放珊瑚,但是以前的许多文献里列为八放珊瑚;软珊瑚和柳珊瑚有时也被视为同义名,有时候又分成两个目。本文无意涉足分类学上的纠纷,在讨论深水珊瑚时不再区分柳珊瑚和软珊瑚。与石珊瑚形成的外骨骼不同,软珊瑚都有由骨片(sclerite)组成的内骨骼,骨片又由有机质的珊瑚硬蛋白(gorgonin)胶结,这是一种纤维状的蛋白质,质地有点像哺乳类的角,因此不像石珊瑚那样坚硬。
一部分种类具有钙质的中轴骨,还可以呈树状分枝,形成海底树木的模样。
图6

图6
深水软珊瑚实例
(a)Paragorgia arborea的群体与珊瑚虫;(b)Primnoa resedaeformis的群体与珊瑚虫;(c)红珊瑚Corallium rubrum的群体与珊瑚虫[10]
Fig.6
Examples of deep-sea Alcyonacea
(a) Community and polyp of Paragorgia arborea; (b) Community and polyp of Primnoa resedaeformis; (c) Community and polyp of the precious coral Corallium rubrum[10]
南海深水珊瑚林里最为突出的是竹节珊瑚(图1;图7a,b)。竹珊瑚是属于Isididae科的软珊瑚,常见的有Acanella,Isidella和Keratosis等属。竹珊瑚的杆像竹子,碳酸盐的杆和枝都被有机质(珊瑚蛋白等)的“节”分段(图7d),以保证有一定的弹性。活的珊瑚虫生长在群体的上端(图7c),迎着水流捕食为生。由于外形变化多端,按照外形分类就会引来许多矛盾。比如同一个属,不分枝的被定为Lepidisis,分枝的为Keratosis,经过分子生物学检验其实 是同一个属[17]。上面说到,红珊瑚之所以成为珍宝是因为碳酸钙的骨骼特别致密,竹节珊瑚杆、枝的碳酸钙也极为致密,因此会被不法商人染红后假冒红珊瑚出售。
图7
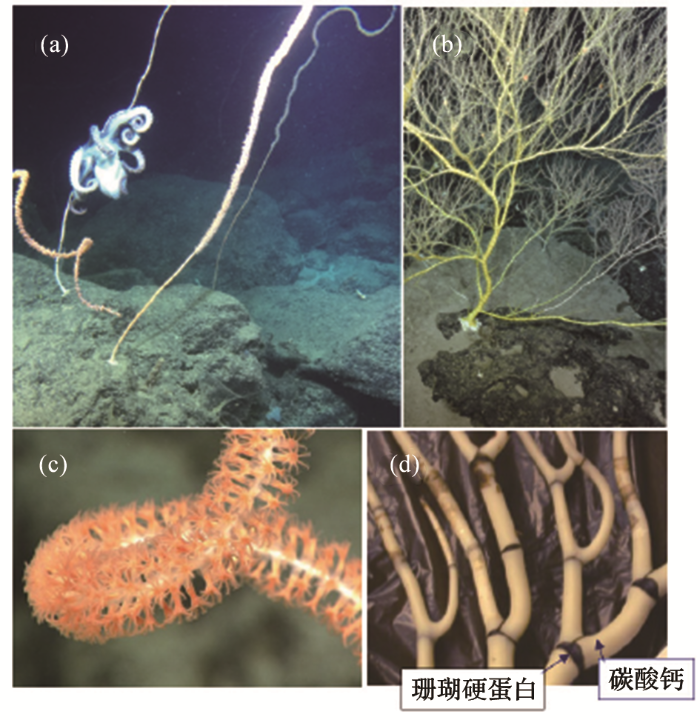
图7
竹节珊瑚
(a) 南海不分枝的竹节珊瑚; (b) 南海海山上分枝的竹节珊瑚(周怀阳提供);(c) 竹节珊瑚上端的活珊瑚虫;(d) 竹节珊瑚下部碳酸盐的干、枝,由暗色的珊瑚硬蛋白分成竹节状(c,d选自网页)
Fig.7
Bamboo coral
(a) Un-branched bamboo coral in the South China Sea; (b) Branched bamboo coral found on a deep-water sea-mount of South China Sea (provided by Zhou Huaiyang); (c) Living polyps at the upper part of a bamboo coral; (d) Carbonate trunks and branches of bamboo corals, divided by dark proteinic gorgonian into bamboo-like pattern (photos in c and d from the internet)
竹节珊瑚是全球分布的深水珊瑚,连两极都不例外,分布的深度可超过4 000 m,都在石质基底上生长。不过在加拿大巴芬湾的北极圈内,深逾900 m的泥质基底上也曾发现有密集的竹节珊瑚林,但不清楚是否是原来长在砾石上后来被沉积物覆盖所致[18]。
3.2 深水珊瑚林的分布
无论造礁或者造林深水珊瑚的属种,更确切地应该叫冷水珊瑚,因为其在浅水里也可以分布,关键在于水温。冷水珊瑚生活在4~12 oC的低温海水里,并不在乎水深,因此从赤道到极地、从浅水到深海,都有这类冷水珊瑚出现。在高纬度海域这种水温出现在50~1 000 m较浅的海域,而在低纬海域则局限于深海,可以达到4 000 m[9]。比如竹珊瑚Tenuisis microspiculata与其他冷水珊瑚一起,在南极洲出现在水深10~45 m的潮下带[19];在阿拉斯加岸外,柳珊瑚在100~200 m最多,每平方米平均1.23个群体,最高有3.85个/m2的珊瑚林[20];更高的密度则见于阿留申群岛的珊瑚林,每平米上的软珊瑚超过5株群体[10]。
与深水珊瑚礁局限于大西洋不同,软珊瑚的分布遍及全球,因此个别的深水珊瑚在各个海洋都可以出现,但是只有密集分布才能够成“珊瑚林”。深水珊瑚林的分布主要受3个条件限制:硬质的基底,适宜的海水和足够的食物。硬基底的种类不限,从深海玄武岩到冰砾都可以,因此深水海山和深沟断崖都是合适的条件,尤其因为强烈起伏的地形有利于形成活跃的海流,从而带来更多的食物。深水珊瑚靠羽状触须捕食,它们的触须顺着海里水流的方向生长,这样才能捉到海水流动带来的食物,主要是小型浮游动物、粪粒和有机质碎屑[9],海面降下的海雪是珊瑚食物的主要源头,但是对于深水珊瑚来说,悬浮的细颗粒也是重要的食物来源。
软珊瑚的外形与取食的需求相对应。竹节珊瑚主干依靠分节提供的弹性,可以迎着水流弯曲,而生长活珊瑚虫的“竹竿”末端可以旋卷摇曳,在水流中灵活捕食;扇珊瑚总是以“扇面”朝向水流,因此可以凭扇珊瑚的方向判断海水的流向[7]。所以柳珊瑚有3种形状:双向水流处呈扇状,湍流处为鞭状或者疏枝状,单向流处为内凹状。
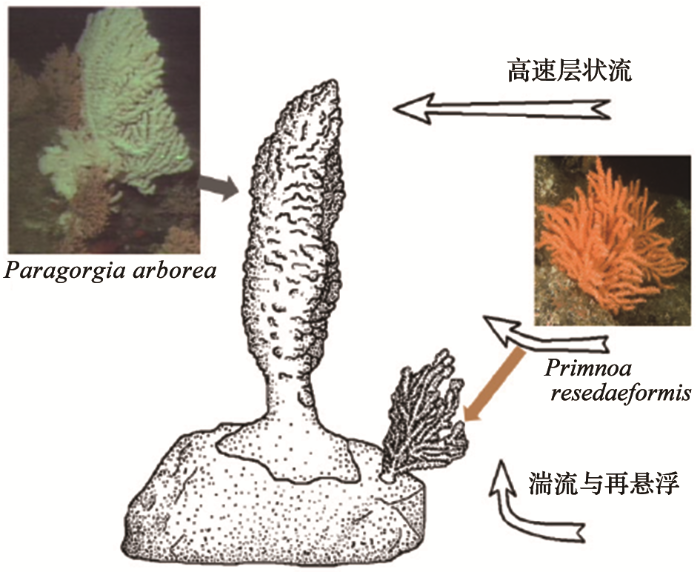
以加拿大岸外大西洋200~600 m的深水珊瑚为例,比较大的有2种常见的软珊瑚:Paragorgia arborea(图6a),高达180 cm,枝比较粗;Primnoa resedaeformis(图6b)比较小,最高86 cm,枝较细。P. resedaeformis属于疏枝状,而P. arborea随着生长从疏枝状变为内凹状。在海底,P. resedaeformis常长在卵石的侧面,P. arborea长在卵石顶面,其形状与水流对应:P. resedaeformis体积小,对应于海底的乱流;P. arborea体积大,已经超越了海底乱石造成乱流的高度,因而适应于固定的水流方向(图8)[21]。
图8
4 深水珊瑚生态系统
深水珊瑚群的发现改变了人类对海洋生态系统的理解。如果说深水珊瑚礁的分布范围主要局限于大西洋,那么深水珊瑚林却在全大洋普遍发现,从而为深海在热液、冷泉之外又增添了一种更为普遍的海底生态系统。它们主要依靠源自上层的有机物为生,加上能够固着的基底和活跃的水流这两个必要条件,因此在营养丰富、地形复杂的海山、峡谷等区广泛发育。固着生活的冷水珊瑚群落与陆地的园林颇为相似,也像陆地树林那样为活动的动物提供了栖居地。不同的是形态,冷水珊瑚群体可以像竹珊瑚那样成杆状或者鞭状,以一维几何形态在流水中游转,或者像扇珊瑚那样呈二维几何形态迎流招展,因此很少有像陆地树木那样呈三维形态,这样有利于在流水中立足。
作为21世纪深海生物学的重要突破,深水珊瑚群的发现归功于新技术的应用。地球上的生境90%以上在海里,而除了大陆架,88%的海域水深超过1 000 m,因此人类关于海洋生物的知识主要来自海岸和浅海生态系,至于深海海底,研究过的只占约0.0001%[22]。200 m以下的深海一片漆黑,在软基底上的深海底栖生物还可以用沉积取样的方法调查,而对硬基底的深海只能依靠拖网或者推想。但即便是软基底,靠沉积采样的方法也只是“瞎子摸象”,只有深潜观测才会发现真面目。比如新西兰的Kaikoura海沟采用新技术Van Veen grab抓样调查,发现900~1 000 m海域有丰富的海参、多毛类蠕虫和海胆等动物,居然平均每平方米就有516个,比超过500 m的深海丰度高出10倍之多[23]。深水珊瑚群也是一样,正是载人或不载人深潜器的运用,方才揭示了海洋生态系统研究中的这个重要的遗漏环节。
深潜技术发现的“深水珊瑚林”,与陆上的树林有许多相似之处。竹节珊瑚之类高达数米的珊瑚群体,有时还分支分叉,相当于树林里的乔木;高度不过几十公分的扇珊瑚之类,相当于陆上的灌木;而更矮的珊瑚和苔藓虫、玻璃海绵等,类似于陆上的草本植物。此外还有一类不属于软珊瑚的八放珊瑚叫做海鳃(Penattula),在羽状的群体下端还有个短柄,可以插在泥沙中固定,因此往往在软基底上密集分布,宛如陆地上的秧田。
图9

其实深海造林的动物不仅有珊瑚,海绵和苔藓虫等也都有贡献[25],只是不如珊瑚那样突出。现在,学术界正式提出了“海洋动物林”的新概念:陆地上有植物林,海底下有动物林,而且是全球分布的动物林,甚至于南极海底也会有“海绵场(sponge ground)”[26]。当然,“海洋动物林”无论分布和种类都极不均匀,而且随着生命演化的变更,如今海洋中的珊瑚扮演着主角。按照美国的提法,“深水珊瑚”是指无虫黄藻共生的石珊瑚和软珊瑚,分布在50~6 000 m深处,已知的有3 300种以上[15]。海山由于其硬质基底、水流加速、生产力高,是深水珊瑚发育的理想地点[27]。而海底油气苗有利于冷水珊瑚礁的发育,有助于在大西洋东北油区附近的发育[5]。
5 深水珊瑚的地质意义
深水珊瑚礁、珊瑚林的发现是地质界的一大福音,既为深海古环境再造提供了无可替代的高分辨率记录,又为理解地质时期生命与地球过程耦合的重要环节提供了现代样本。底栖动物造礁、造“林”,应当是地质历史的“常情”,只是地质界识悟太晚。追溯起来,“海洋动物林”在寒武纪大爆发前已经出现,叶子模样的渗养生物(osmotrophs)可以在前寒武纪的海底上竖起2 m之高,以便在刚开始有氧的海底直接以溶解有机碳为生,可惜这类“埃迪卡拉动物群”只有印模化石,难以进行深入分析[30,31]。后来的海洋里造礁生物变多,从寒武纪早期的古杯海绵到白垩纪的固着蛤、厚壳蛤,以及古生代各种各样的珊瑚。海底造林的动物也极其多样,也许最为出彩的是中晚古生代的海百合。
图10
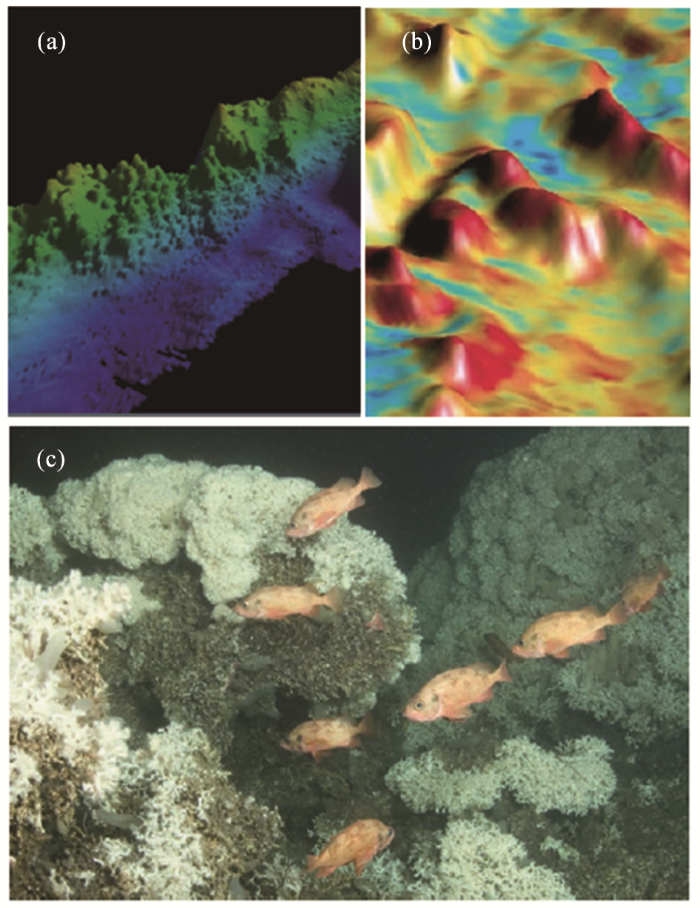
图10
运用多波束回声测深仪探索北大西洋的L. pertusa深水珊瑚礁
(a)多波束地形图揭示挪威岸外的珊瑚礁泥丘;(b) 苏格兰岸外的珊瑚礁泥丘;(c) 挪威岸外的L. pertusa深水珊瑚礁照片[10]
Fig.10
Applying multi-beam echo sounder to explore the L. pertusa deep-sea coral reefs in the North Atlantic
(a) Coral-reef mud-mounds along the Norwegian shore revealed by multi-beam typography; (b) Coral-reef mud-mounds offshore Scotland; (c) A photo of L. pertusa deep-sea coral reef offshore Norway
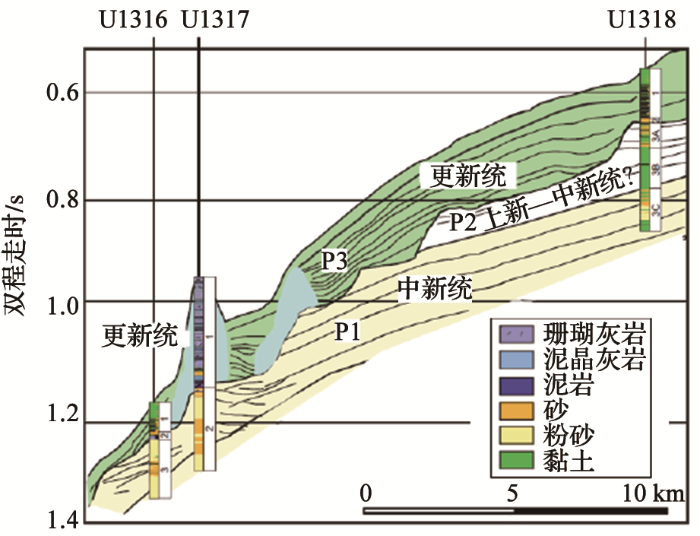
地质界格外注意大西洋东北的深水珊瑚礁密集分布区,这里爱尔兰西南单独一个Porcupine Seabight海盆就有2 000个珊瑚礁泥丘,是什么机制导致深水珊瑚礁的大量发育?目前有2种假说——内因说和外因说。内因说强调珊瑚礁和深部油气的关系:这里的珊瑚礁不但发育在含油气地层之上,附近还有油苗活动。于是产生了假说:来自深层的烃类流体运移促使珊瑚礁开始发育,加上甲烷菌氧化作用能提高碱度,有利于碳酸岩丘的形成[5]。外因说强调海流:这里的珊瑚礁发育在2个分层海水的界线附近,上有北流的东北大西洋暖水,下有高盐的地中海外流水,底流强劲、营养丰富,这种海流格局的形成才是深水珊瑚发育的原因。2005年,大洋钻探IODP 307航次在爱尔兰西岸外钻探冷水珊瑚礁,对2种假说进行了检验(图11)[35]。结果发现冷水珊瑚礁开始形成于195万年前,以后的发育又与冰期旋回相对应,但是未能检测出珊瑚礁有加强的微生物活动。因此,大洋钻探的结果支持冰期旋回控制的外因假说,否定了与油气运移相关的内因假说[36]。关于IODP 307航次十多年前就有报道[37],此处不再重复;需要介绍的倒是20多年前大洋钻探的ODP 182航次。
图11
本文主题是深水珊瑚礁和珊瑚林,但是在地质历史上多种门类都可以在海底形成碳酸盐结构进入地层,并非珊瑚独有的专利。澳大利亚南岸外的大澳大利亚湾(Great Australian Bight),在陆坡顶部地层里就有由苔藓虫组成的碳酸盐泥丘,1998年大洋钻探ODP 182航次在这里钻探了9个站位,取了3 500 m岩芯[38],发现150 m地层里记录了将近30万年来的苔藓虫碳酸盐泥丘的发育史。这里有枝状、扇状和块状等多种多样的苔藓虫(图12c),像珊瑚一样形成碳酸盐泥丘(图12b)。每层苔藓虫碳酸盐的厚度为5~10 m,反映了碳酸盐泥丘的形成受生长窗口的限制:窗口的上限是波浪破碎的底界,下限是贫养南极中层水的顶界(图12a)。冰期时亚热带辐合带北移,亚南极模态水带来的营养物使这里出现高生产力;间冰期来临时水团发生变化,苔藓虫礁的生长难以为继,碳酸盐泥丘堆积便告中断。大洋钻探的岩芯取得了更新世多个冰期的记录,但是这类苔藓虫礁在今天已经不再出现[39]。可见,苔藓虫提供的古海洋学信息,并不亚于珊瑚礁。
图12
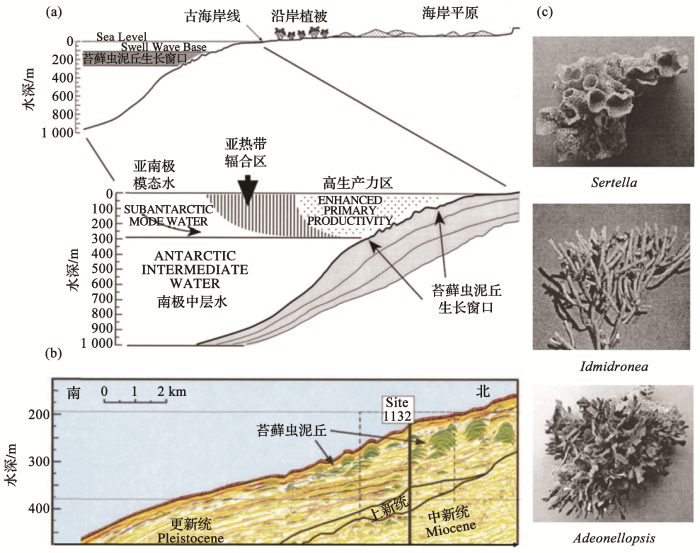
图12
大洋钻探ODP182航次在大澳大利亚湾揭示的更新世苔藓虫碳酸盐泥丘
(a)苔藓虫碳酸盐泥丘的生长窗口;(b) ODP 1132井地震剖面显示的苔藓虫碳酸盐泥丘的地层分布;(c)形成碳酸盐泥丘的几种主要苔藓虫[39]
Fig.12
Ocean Drilling Program Leg ODP 182 revealed the Pleistocene bryozoan carbonate mud-mount in the Great Australian Bight
(a) Living window for bryozoan carbonate mud-mount; (b) Stratigraphic distribution of bryozoan carbonate mud-mount at Site ODP 1132 revealed by seismic profile; (c) Major bryozoan taxa forming the carbonate mud-mount[39]
6 结语:深水珊瑚的研究
南海深水珊瑚林的发现,把我们的注意力引向了深海海底的底栖生物。人们对深海珊瑚并不陌生,因为许多深水珊瑚的群体在百米上下的浅水里也有零星出现,只是不能像在深海那样成礁、成林。这些类型实际上是冷水珊瑚,并不限于深海,因而作为海洋药物或者珍宝,早已为社会知晓。21世纪深潜技术的应用,发现深海珊瑚构成了全球分布的底栖生物群,提供了深海气候环境变迁的珍贵档案,引起了学术界极大的重视; 只是受技术手段的限制,研究主要限于欧美岸外和澳大利亚、新西兰一带,大部分发展中国家岸外属于研究的空白。
与此同时,深水珊瑚林也正在成为海洋环境保护的新命题。深水珊瑚生长极其缓慢,却是很容易受到破坏的脆弱生态系统。现在的渔业技术不断更新,并且向深海远洋发展,所采用的破坏型技术直接损害着深海珊瑚群;再说全球变暖引起海水“酸化”,尤其不利于文石骨骼珊瑚礁的发育[40,41]。为此,欧美各国近年来分别立项开展调查研究。比如美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration,NOAA)设立了“深海珊瑚研究与技术计划(Deep Sea Coral Research and Technology Program)”,每年用大约250万美金对其各个海区的深水珊瑚进行调查监察[42]。深海珊瑚也引起了联合国环境规划署(United Nations Environment Programme,UNEP)的注意,由其麾下的世界保护监测中心(World Conservation Monitoring Centre,UNEP-WCMC)发布报告,呼吁各国主管部门加以注意[7]。
深水珊瑚也是国际学术界的新兴热点。进入21世纪以来,欧美学者已经组织了7届“深海珊瑚国际研讨会(International Symposium Deep-Sea Corals)”,先后在加拿大Halifax(2000年)、德国Erlangen(2003年)、美国Miami(2005年)、新西兰Wellington(2009年)、荷兰Amsterdam(2012年)、美国Boston(2016年)和哥伦比亚Cartagena(2019年)举办,一般200人的规模,范围从地质、生物到管理,也在Deep-Sea Research等学报发表专辑[43],但参加者主要来自西欧、北美和新西兰,很少有亚洲人。
南海深水珊瑚林的发现,应当唤起我国学术界的注意。一方面要抓紧利用深海珊瑚的骨骼开展分析,为南海提供中层水和深层水的高分辨率古海洋学信息,补上以往研究中的缺口;另一方面要对深水珊瑚林的生态系统进行调查研究,设立地质和生物相结合的跨学科项目,尤其是要采用高新技术调查深海珊瑚林的分布格局。载人深潜是深水珊瑚研究的必要手段,同时为了扩大调查面积、提高调查效率,尚待使用ROV和AUV开展大面积的观察。国外开始采用模拟分析的技术,根据生长深海珊瑚林的地质和水文等自然条件进行分析,预测深水珊瑚林的分布与种类[44],也值得我们借鉴。可以预期,一旦我国有关部门做出决定,通过跨部门、跨学科的合作,定能带动国际学术界,使南海成为西太平洋深水珊瑚林研究的制高点。
参考文献
Discovery of deep-water bamboo coral forest in the South China Sea
[J].
Benthic biodiversity across and along the continental margin: Patterns, ecological and historical determinants, and Anthropogenic threats
[C]// Wefer G, Billett G, Hebbeln D,
The “Sardinian cold-water coral province” in the context of the Mediterranean coral ecosystems
[J].
Long-term baited lander experiments at a cold-water coral community on Galway Mound (Belgica Mound Province, NE Atlantic)
[J].
Ahermatypic coral banks off mid-Norway: Evidence for a link with seepage of light hydrocarbons
[J].
Two pillars of cold-water coral reefs along Atlantic European margins: Prevalent association of Madrepora oculata with Lophelia pertusa, from reef to colony scale
[J].
Cold-Water Coral Reefs: Out of Sight—No Longer Out of Mind
[R].
Structure and composition of organic reefs and carbonate mud mounds: Concepts and categories
[J].
Reefs of the deep: The biology and geology of cold-water coral ecosystem
[J].
Cold-Water Corals:The Biology and Geology of Deep-Sea Coral Habitats
[M].
The giant Mauritanian cold-water coral mound province: Oxygen control on coral mound formation
[J].
Lithoherms on the Florida-Hatteras slope
[J].
Deep-sea carbonate: Pteropod distribution and the aragonite compensation depth
[J].
A brief history of taxonomic research on azooxanthellate Scleractinia (Cnidaria: Anthozoa)
[J].
Deep-water corals: An overview with special reference to diversity and distribution of deep-water scleractinian corals
[J].
Cold-water corals in a changing ocean
[J].
Genetic analysis of bamboo corals (Cnidaria: Octocorallia: Isididae): Does lack of colony branching distinguishing Lepidisis from Keratoisis?
[J].
Deep-water bamboo coral forests in a muddy Arctic environment
[J].
Taxonomic study of suborder calcaxonia (Alcyonacea: Octocorallia: Anthozoa) from King Sejong Station, Antarctic
[J].
Coral habitat in the Aleutian Islands of Alaska: Depth distribution, fine-scale species associations, and fisheries interactions
[J].
Morphology and growth of the Deep-water gorgonians Primnoa resedaeformis and Paragorgia arborea
[J].
Predicting global habitatsuitability for stony corals on seamounts
[J].
Submarine canyons: Hotspots of benthic biomass and productivity in the deep sea
[J].
Deep-sea suprabenthic communities: The forgotten biodiversity
[C] // Rossi S, Bramanti L, Gori A,
Sponge grounds as key marine habitats: A synthetic review of types, structure, functional roles, and conservation concerns
[C]// Rossi S, Bramanti L, Gori A,
Animal forests of the world: An overview
[C] // Rossi S, Bramanti L, Gori A,
Box 7: Deep-sea corals on seamounts
[J].
Marine animal forests: Foreword
[C] // Rossi S, Bramanti L, Gori A,vii-x
.
Living in the Canopy of the animal forest: Physical and biogeochemical aspects
[C]//Rossi S, Bramanti L, Gori A,
Reconstructing a lost world: Ediacaran rangeomorphs from Spaniard’s Bay, Newfoundland
[J].
Canopy flow analysis reveals the advantage of size in the oldest communities of multicellular eukaryotes
[J].
Skeletal Mg/Ca in Primnoa resedaeformis: Relationship to temperature?
[C]// Freiwald A, Roberts J M.1 061
-
Radiocarbon-based ages and growth rates of bamboo corals from the Gulf of Alaska
[J].
Acoustic mapping using a multibeam echosounder reveals cold-water coral reefs and surrounding habitats
[J].
IODP Expedition 307 Drills cold-water coral mound along the irish continental margin
[J].
The pleistocene cooling built challenger mound, a deep-water coral mound in the NE Atlantic: Synthesis from IODP Expedition 307
[J].
A review of cenozoic deep sea cold-water carbonate mounds and preliminary results of carbon and oxygen isotopes from IODP 307
[J].
新生代深海冷水碳酸盐泥丘成因及IODP 307航次初步研究结果
[J].
Great Australian Bight: Cenozoic Coolwater Carbonates
[R].
Origin of Late Pleistocene Bryozoan Reef Mounds: Great Australian Bight
[J].
Will human‐induced changes in seawater chemistry alter the distribution of deep‐sea scleractinian corals?
[J].
Northeastern Atlantic cold-water coral reefs and climate
[J].
The State of Deep‐Sea Coral and Sponge Ecosystems of the United States
[R].
Introduction: Proceedings ISDSC5
[J].
Geological features supporting deep-sea coral habitat in Atlantic Canada
[J].

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}