
王龙 , KhalidLatif
, KhalidLatif
Wang Long, Khalid Latif
中图分类号: P578.6
文献标识码: A
文章编号: 1001-8166(2018)10-1005-19
收稿日期: 2018-04-12
修回日期: 2018-09-4
网络出版日期: 2018-10-10
版权声明: 2018 地球科学进展 编辑部
基金资助:
作者简介:
First author:Wang Long(1990-),male,Huining County,Gansu Province, Ph.D student. Research areas include sedimentology and petroleum geology. E-mail:wanglong1127@163.com
作者简介:王龙(1990-),男,甘肃会宁人,博士研究生,主要从事沉积学和油气地质学的研究工作.E-mail:wanglong1127@163.com
展开
摘要
微生物碳酸盐岩以其漫长的地史分布、多样的产出环境、复杂的沉积构成和与微生物活动密切相关的特性而成为沉积学研究的焦点,同时也是沉积学研究的难点,被誉为“最难研究的沉积岩之一”。其研究的中心主要是钙化微生物席和微生物膜及其地质记录,即石化/成岩作用的产物。从成因着手,系统总结和梳理了2个类型的微生物群落,较薄的生物膜和较厚的微生物席,经过生物/有机矿化作用和石化/成岩作用形成微生物碳酸盐岩的过程。提出除微生物群落自身的复杂性和沉积过程的复杂性之外,多重状生物膜是导致微生物碳酸盐岩多样性的另外一个重要原因。以中国华北地台寒武系微生物碳酸盐岩的研究为基础,梳理了不同类型的微生物碳酸盐岩的沉积特征和分类依据。认为微生物碳酸盐岩应该置于Embry和Kloven描述的黏结岩类下,根据宏观组构可进一步分为叠层石、凝块石、核形石、层纹石和均一石5类,树形石不易独立成类而应归于凝块石。微生物碳酸盐岩为整个地质历史时期与微生物活动相关的生物/有机矿化作用过程研究提供了有用信息,不但是了解地球表层环境演变的一个重要窗口,而且还形成和储集了较大规模的油气,具有重要的科学意义和经济价值。
关键词:
Abstract
Currently, sedimentologists focus on the challenging issue of microbial carbonates, which are regarded as "one of the sedimentary rocks most difficult to study", having complicated sedimentary fabric. Their characteristic features closely related to microbial activity, distributed over a long period of geological time, and formed in diversified sedimentary environments. The main research concentrations are the calcified microbial mats and biofilms in geological records as the products of lithification and diagenesis. Starting from the origin, this paper systematically reviewed and explained the processes dwelling within two types of microbial communities, the thinner biofilm and the thicker microbial mat, which enabled them to convert into microbial carbonates through biomineralization and lithification. This study proposed that the existence of multiple microbial mats was another important cause for the diversification and complexity of microbial carbonates in addition to its complex depositional process. Moreover, the sedimentary characteristics and classification of different types of microbial carbonates were reviewed, exemplifying the Cambrian microbial carbonates in the North China Platform. These microbial carbonates are suggested to be placed under "bindstone" after Embry and Kloven, which can be further divided into 5 types, stromatolites, thrombolites, oncolites, laminites and leiolites. Dendrolite is not categorized as a separate class, instead attributed to thrombolites. The microbial carbonates may possess good source rock potential because of the enriched organic content, and may also serve as hydrocarbon reservoirs because of certain microbial textures and fabrics leading to significant porosity and permeability. Because of their biomineralization processes related to microbial activity, the microbial carbonates are not only an important window to understand the evolution of the earth's surface environment, but also capable of forming large-scale reservoirs, and their scientific and economic values are self-evident.
Keywords:
顾名思义,微生物岩(microbialite)是在与微生物相关的作用下形成的岩石。Burne等[1]在1987年首次将其定义为“底栖微生物群落通过捕获与黏结碎屑沉积物,或(并)形成矿物沉淀中心而产生增生及加积作用,在原地形成的生物成因的沉积”。鉴于微生物岩主要以碳酸盐岩的形式产出,Riding[2,3]提出了微生物碳酸盐岩(microbial carbonate)的概念:“与微生物生长、新陈代谢、细胞表面性质,以及细胞外聚合物(Extracellular Polymeric Substances,EPS)相互作用有关的沉积物(岩)”,并将其分为叠层石、凝块石、树形石和均一石四大类。梅冥相[4]进一步认为核形石和层纹石也有较独特的组构特征、发育背景和广泛的产出环境,应纳入微生物碳酸盐岩分类体系。
这些微生物碳酸盐岩(微生物岩)形成了从太古宙至今极为壮观的沉积:①在所有类型生物成因的灰岩中,它们有着最长的地质分布范围,出现于3.7 Ga[5];②它们形成于各种各样的环境中,包括海洋、湖泊、河流、溶洞和土壤等[2,6];③它们氧化了地球的大气圈,固定了大量C
因而,系统回顾和总结前人的研究成果,厘清微生物碳酸盐岩的形成机制、分类方案和沉积特征,以及当前所面临的挑战对微生物碳酸盐岩的研究很有必要。本文从微生物碳酸盐岩的成因入手,阐述了2个类型的微生物群落:较薄的生物膜和较厚的微生物席,经过生物/有机矿化作用和石化/成岩作用形成微生物碳酸盐岩的过程。并以中国华北地台寒武系微生物碳酸盐岩研究为基础,梳理了不同尺度下微生物碳酸盐岩的沉积特征和分类依据。微生物碳酸盐岩为整个地质历史时期与微生物活动相关的生物/有机矿化作用过程研究提供了有用信息[2,3,7],不但是了解地球表层环境演变的一个重要窗口,而且还形成和储集了较大规模的油气,具有重要的科学意义和经济价值。
我们对碳酸盐沉积及成岩作用的理解在过去10年中发生了很大变化,大部分灰岩本质上是生物成因,现代环境中超过90%的碳酸盐沉积被认为是生物起源的[11]。生物尤其是微生物的作用对于理解碳酸盐岩的形成机制至关重要。但微生物碳酸盐岩简单的矿物相(通常是单矿物相)往往掩盖了它们岩相学、化学性质和沉积过程的复杂性[8,11]。为弄清微生物碳酸盐岩复杂的成因机制,现代微生物席、钙化微生物席的地质记录,以及它们石化作用的产物成为了关键。
1.1.1 微生物膜和微生物席的概念及特征
地质学家很早就认识到一些显微级别的生物如藻类,可能与叠层石等的形成相关,在这些描述中藻席和生物结壳是2个常见的术语[11,12,13,14]。随着微生物岩概念的提出和成型[1,2,3],研究的焦点聚焦向形成微生物岩的微生物,发现其包括细菌、病毒、小型藻类和原生动物等在内的一大类生物群体,原来“藻席”的概念开始被“微生物席”所替代,而生物结壳本质上就是微生物膜[9,15]。
这些微生物群落形成的微生物膜(biofilms)和微生物席(microbial mats)出现于3.7 Ga的太古宙地层,构成了地球上最早的生态系统并持续至今[5,6,16,17],深刻影响着地球大气圈、水圈和岩石圈的演变[18,19]。但截止目前,对于“什么是微生物膜或微生物席”还没有统一的认识,所幸无论是较薄的微生物膜(纳米到微米级)还是较厚的微生物席(毫米到厘米级)[20,21,22,23],一些共有的属性和特征已经被很好地发现与总结:①生物学上的生理协同性,即构成微生物群落的所有微生物可以相互协同作用和工作[9,24];②EPS的黏附性和将细胞自身与其他有机物及/或无机物连接在一起的性能[25,26];③微生物群落可以在极薄的、不同的纹层内创造出较宽泛的化学梯度,使得纹层状结构和环境微梯度存在[9,27];④微生物的增生作用[3];⑤微生物席(膜)或多或少都可以看作是一个固着的底栖生态系统,因为它们常生长在不同物质的交界面(尤其是浸没的或者潮湿的物体表面),嵌入在黏液状的物质中并且固定在界面附近的底质上[24,28]。
简而言之,微生物席(膜)由微生物自身、微生物产生和分泌的EPS,以及捕获或者沉淀的外来物质3个部分构成(图1),但这3部分呈现为一个整体而不能分割。如同人类建造房子一般,微生物产生和分泌主要由多糖、糖蛋白、细胞外酶和核酸所组成的EPS(图1b),来增强细胞的适应性和弹性,抵抗不良环境的影响[32,33]。但EPS的功能和意义远非房子可比拟,它和捕获、黏结或沉淀的外来物质(主要是无机物)一起,在薄薄的几厘米甚至几毫米内就创造出较为宽泛的化学梯度,供各种各样的微生物种群定居和生活[27,29]。这种纹层状的微生物体系结构复杂、微生物种群多种多样,从表层到较深层微生物活动总体呈递减状态,在位于透光带时经常为蓝细菌所构建[34] (图1 a,c)。例如巴西东南部Lagoa地区发育的层状微生物席,自上而下依次发育:①黄色富EPS层;②绿色光合作用的蓝细菌层;③紫色硫细菌为主的红色层;④灰色硫酸盐还原细菌层,这些微生物席纹层在仅仅为4 cm的尺度范围内就完成了从富氧层到厌氧层的转换[35]。更有甚者,Bouton等[32]在研究古巴现代微生物席时,对顶部10 mm进行氧气和硫化物的微电极测量,发现氧气的峰值对应于绿色纹层,即由蓝细菌主导的纹层段;向下氧气的幅度递减明显,转变为少氧或缺氧环境,对应的微生物也变为硫酸盐还原细菌等(图1a);而微生物席的底部已开始石化成岩(早期成岩作用),并保存了部分钙化微生物残余物(图1a, c)。
图1 微生物席(膜)的构成及结构(据参考文献[
(a)古巴现代微生物席:纹层A为2014年发育的微生物席,纹层B为表面黏性微生物席,纹层C为较早(古代)形成的微生物席及沉积物;对微生物席的顶部10 mm进行氧气和硫化物的微电极测量(方框位置),发现氧气的峰值对应于绿色纹层(纹层A的顶部),表明了较高的光合作用活动(据参考文献[
Fig.1 The fabric and structure of microbial mats and biofilms (modified after references[29~31])
(a) Modern microbial mats of Cuba: Layer A corresponds to the microbial mat developed during the year 2014; Layer B is the surface cohesive mat;Layer C corresponds to ancient mats and sediments; Microelectrode measurements of oxygen and sulphide in the top 10 mm of the mat (the square indicates the location of the measurements). The O2 peak is associated with the green laminae (upper part of Layer A) and indicates high photosynthetic activity (modified after reference[29]). (b)The abundant Extracellular Polymeric Substance (EPS) secretions (green) surrounding cyanobacteria and heterotrophic bacteria (blue) within a stromatolite microbial mat. The large round objects (red) are calcium carbonate sediments,called ooids. Image was collected using a confocal scanning laser microscopy (modified after reference [30]) . (c) Cyanobacterial biofilms in stromatolites, which are dominated by filamentous cyanobacteria (C), and a small number of Coccoid Cyanobacteria (CC). The lower lithified portion shows the residues of cyanobacterial filaments (LS; modified after reference[31])
1.1.2 微生物膜和微生物席中的钙化微生物
在所有微生物席中,由蓝细菌所主导的微生物席最被关注。蓝细菌以前称为“蓝绿藻”,能够进行光合作用和固氮作用[15,36]。作为色素体的创始者,蓝细菌在藻类和植物演化中扮演着重要角色[37]。尽管在现代正常环境中一般很少作为主要的沉积物,但在地质历史时期是一种可以大规模形成微生物碳酸盐岩的极为重要的微生物[35,36,38]。蓝细菌光合作用吸收的无机碳能够促进碳酸钙的沉淀,这种钙化作用会在微生物席中产生丝状微化石(图1和图2),最终作为叠层石、凝块石等保存起来。蓝细菌的钙化作用也能引起湖底或海底碳酸盐泥的沉淀,形成“白垩”等事件[35,39,40]。蓝细菌是可以移动的,其生命活动具有向上穿越性,即在沉积物堆积后能够向上越过新的沉积物,在界面之上建立新的表面微生物席,之下则石化成岩(图1c)。此外,由蓝细菌所主导的微生物膜可以发育在沉积物和碎屑颗粒内部,并对早期成岩作用有所贡献,以生物结壳和泥晶套最为常见[9,41]。它们适应能力极强,分布广泛且种类多样,从前寒武纪至今,从冰川到热泉、从正常盐度到超盐度环境,都有一系列蓝细菌的钙化残余物及活动遗迹被发现[35,38]。
图2 钙化蓝细菌的形态学分类(据参考文献[
(a)~(c)为管状蓝细菌,其中葛万菌的菌丝未分支,管状构造发育良好,互相不规则的缠绕状,单一直径的管状丝状体小于50 μm,并且具有一厚度均匀的薄壁;取样层为寒武系徐庄组,取样地点吉林白山剖面;(d)~(f)团块状、棒状或球粒状致密泥晶构成的蓝细菌,其中附枝菌为粗枝杈状,枝晶状生长;取样层为寒武系凤山组,取样地点辽宁金州湾剖面;(g)~(i)中空状(具房室状)蓝细菌,其中肾形菌或饼状菌呈不规则肾形或小球状,泥晶壁的厚度约为10 μm,中腔为微亮晶方解石填充;取样层为寒武系碱厂组,取样地点辽宁复州湾剖面
Fig.2 Morphological classification of cyanobacteria (modified after reference[35]),and some typical Calcified cyanobacteria of Cambrian in the North China Platform
(a)~(c) Tubular cyanobacteria, in which the Girvanella have unbranched, well developed tubular, and irregularly tanged filaments with a diameter less than 50 μm,and uniform thickness; Cambrian Xuzhuang Formation, Jilin Baishan section. (d)~(f) The cyanobacteria are composed by lumpy,rod-shaped or dense micritc thalli, with branching and dendritic grown; Cambrian Fengshan Formation, Liaoning Jinzhou Bay section. (g)~(i) Hollow chambers of cyanobacteria, in which the Renalcis or Tarthina are preserved in irregular kidney or spherical shapes, with micritic walls of 10 μm thickness, and their cavities are filled with micro-sparite; Cambrian Jianchang Formation, Liaoning Fuzhou Bay section
钙化蓝细菌的分类主要依据形态学特征,3种主要的形态群为:①管状;②团块状、棒状或球粒状(致密泥晶);③中空状(具房室状),在微生物碳酸盐岩中呈不规则凝块或者分枝状形态出现[38](图2)。华北地台寒武系微生物碳酸盐岩中,管状蓝细菌主要为葛万菌(Girvanella),放大后可以观察到发育良好的管状构造,菌丝体的个体相互缠绕和穿插,稠密的泥晶方解石围绕菌丝体鞘选择性沉淀,形成明显的泥晶外壁(图2a~c)。同样是管状菌丝,但是常常呈缆线状分布的类别包括束线菌(Subtifloria)、Batinivia、Botominella、Cladogirvanella和Razumovskia等;螺线菌(Obruchevella)的管状菌丝被盘绕起来或被卷成圈,直管菌(Ortonella)的菌丝类似葛万菌,但有直立分支(图1a)。附枝菌(Epithyton)是华北地台寒武系中另外一种常见的蓝细菌,以明显密而坚固的分枝为特征,这些分枝由棒状或球粒状的致密泥晶构成(图2d~f)。此组形态群还包括由亚毫米级别的垂向极不规则似灌木丛状的致密泥晶构成的角胞菌(Angulocellularia),以及Gordonophyton和中管状菌(Tubomorphophyton)等类型。中空状(具房室状)蓝细菌以肾形菌(Renalcis)最为典型,化石呈肾形或小球状,内部为空腔,外壁由暗色泥晶构成,较厚但厚薄不均,粗糙(图2g~i)。此组形态群还包括芽孢菌(Gemma)、饼状菌(Tarthina)和Izhella 等相似化石,主要依靠壁的厚度和房室数量来分辨,很难区分[35]。
但多数情况下,微生物碳酸盐岩中并未发现真正的钙化微生物化石,我们只是通过整体的沉积构造(例如叠层石普遍发育的叠片状构造和窗格状构造),外部钙化丝状体、球体(例如多数微生物可以形成树枝状生长的泥晶灰岩球粒及细枝状结核)或管状构造,微晶灰岩结壳和不规则的外膜,以及被捕获的沉积物来识别微生物碳酸盐岩,在扫描电镜下有时也可识别出一些哑铃状、棒状、丝状和球状等形态的细菌构造[42,43]。这些钙化微生物沉积物在镜下没有一致的骨骼形态,因为蓝细菌本身多数并未钙化,而是伴随着蓝细菌的生长和向上超越,不含活细胞的、由各种高分子所组成的特殊的有机质覆盖层EPS,在对矿物表面的吸附以及自我组织的催化过程起着关键作用,发生“非骨骼”或者“骨骼外”的钙化[2,44]。这种钙化本质上是生物矿化作用和/或有机矿化作用的结果,是一个复杂的生物化学过程[8]。
数十亿年来,蓝细菌等微生物一直强烈影响着地球上碳、氮和氧等元素的地球化学循环,特别是在碳酸盐的沉淀(钙化作用)中起到了举足轻重的作用[45]。碳酸钙在水中的沉淀与其饱和状态密切相关。蓝细菌等微生物的光合作用会吸收大量的CO2,虽然这个过程对外界环境的pH值影响有限,但活跃的HC
图3 推测的CCMs诱导下蓝细菌鞘的钙化作用机制(据参考文献[
(a)葛万菌丝状体及暗色泥晶鞘(白色箭头);华北地台寒武系徐庄组,辽宁金州湾;(b)CCMs机制:细胞主动吸HC
Fig.3 Presumed mechanism of calcified cyanobacteria sheath induced by CCMs (modified after reference[47])
(a)Girvanella filaments and sheath (white arrow); Cambrian Xuzhuang Formation of North China Platform, Liaoning Jinzhou Bay section;(b) CCMs. The cells actively absorb HC
CCMs导致的蓝细菌钙化作用与细胞关系紧密但又在细胞外发生,最终这些被碳酸钙晶体浸染和保存下来的鞘,保留了蓝细菌管状或灌丛状等形态,但鞘内的细胞丝状体并没有发生钙化而是降解消失了(图2和图3a),因此在这个过程中产生的丝状体是鞘钙化作用的产物而不是细胞本身的钙化产物[44,47]。而且在光合作用所驱动的钙化作用过程中,如果只是在细胞附近沉淀了一些孤立的晶体,那么就无法形成一个可保留的结构,而是以外来颗粒(如“白垩”)的方式释放,在湖底或者海底堆积形成大量微米级别的碳酸盐泥的沉积物[35,44]。CCMs能够被局部的碳限度(比如微生物席或浮游植物导致的水华内),或者大气圈中正在下降的CO2浓度所激发,形成较为普遍的钙化蓝细菌化石[35,44]。显生宙大气的CO2含量波动较大,当下降到大约相当于10 PAL(Present Atmospheric Levels,现今大气含CO2水平),即约0.36%的临界点之下时,钙化化石就能普遍出现,形成蓝细菌钙化作用事件[2,44]。
蓝细菌等微生物绝大部分都不具有钙化骨骼而只是鞘(胶鞘)的钙化,自然未能留下实体化石。所以没法像识别珊瑚、有孔虫等那样去识别出具体的“造岩微生物”,只能宽泛地叫做钙化微生物化石[2,8,49](图2)。实际上,微生物本身一般并不直接产生碳酸钙的沉淀,而是微生物席(膜)有机矿化作用的结果。有机矿化就是受生物诱导和影响的矿化作用,与生物控制的矿化作用不同[8]。在20世纪90年代以前,这2个术语曾被笼统地称为生物矿化作用[50,51]。但为了更好地说明生物的成矿过程及成矿机理,生物矿化作用被限定为由基因控制的、受生物调节的矿化作用过程;只是在生物的影响下,有机聚合物和无机化合物相互作用而发生的矿化称为有机矿化作用[51,52,53]。举例来说,生物骨骼、生物介壳和生物碎屑等就是典型的生物矿化作用产物,而细菌结壳作用、叠层石、硅质热泉沉积等则为有机矿化作用的产物[41,53]。
在某些时候,要准确界定一些微生物的矿化作用过程是生物矿化还是有机矿物还存在困难。例如Drews[54]和Teske等[55]发现形成于某些大型硫细菌细胞内的聚合磷酸盐颗粒和硫化物球粒,为关键的新陈代谢产物提供了存储场所,应该是生物矿化作用的产物;但Schulz等[42]的研究又表明这些硫细菌的活动提高了细胞外孔隙水中的磷酸盐的聚集度,间接导致了磷灰石的沉淀,此时这些磷灰石的形成又是有机矿化作用的结果。但无论如何,前寒武纪地层中那些被认为与微生物作用相关的碳酸盐岩,如叠层石中一直都未能找到钙化的微生物化石[2,8,49]。即便从寒武纪起,用附枝菌、葛万菌等微生物名称来命名的化石开始出现,也大多是一些有机矿化作用的钙化残余物或遗迹化石而非真正的骨骼,真正经过生物矿化作用形成大量碳酸盐岩的微生物——钙质超微生物(主要是颗石藻)直到侏罗纪才开始迅速繁衍[11,43]。微生物的矿化作用(主要是钙化作用)通常以有机矿化作用为主[2,8,45]。
而且,鞘只是EPS的一种结构形式,更多的是大量呈弥散状分布的EPS[55](图1b)。这些弥散状EPS的降解主要依靠硫细菌构成的微生物群落,化能有机异养微生物、发酵微生物、化能有机自养微生物铁还原和硫酸盐还原细菌等(图4):蓝细菌等光合作用微生物率先将无机碳转化为有机碳,在固定CO2环节起关键作用,速率大约为5 g/(d ·m2)[58];有机质的生成滋养了喜氧异养微生物,它们在微生物席中迅速繁殖并消耗掉大量氧气,使得紧邻蓝细菌的厌氧微生物能够存活并分解有机质;喜氧异养微生物可以将大分子有机质降解为小分子有机质,最终被硫酸盐还原细菌、产甲烷细菌及其他“终极”细菌的呼吸作用所分解[8]。这些过程都和碳酸钙的沉淀及溶解密切相关,大致可以分成6种主要的化学代谢反应,光合作用和硫酸盐还原反应通过增加微环境碱度(pH值)促进沉淀,而呼吸作用、硫化物的氧化反应和发酵作用产生的碳氢酸和有机酸则很可能会导致碳酸钙溶解作用的发生(图4)。
图4 微生物席垂向微环境梯度中细菌群落的活动与碳酸钙的沉淀—溶解作用(据参考文献[8,56,57]修改)
(a)微生物席中氧气、硫化物和pH值在24小时内的变化,剖面I和II分别为下午2点和早上3点的“地球化学快照”,可以最显著地反映出微生物席在白天和黑夜之间的不同。当黑夜降临时,由于光合作用的停止和异常细菌的消耗,微生物席很快就转变为缺氧环境(据参考文献[8,56]修改);(b)发生在一个典型的微生物席中的6种主要化学代谢反应,这些反应按对碳酸钙沉淀作用的影响分类;光合作用和硫酸盐还原反应通过增加微环境碱度而促进沉淀,而呼吸作用、硫化物的氧化反应和发酵作用则很可能会导致溶解作用的发生,当黑夜降临时,需氧细菌的活动逐渐停止,厌氧的异养微生物的活动开始占据主导地位。碳酸钙的净沉淀量取决于不同类型的微生物的新陈代谢活动及其时空分布及变化(据参考文献[8,57]修改)
Fig.4 Activities of microbial communities in a microbial mat under vertical microenvironment gradients leading to carbonate precipitation and dissolution (modified after references[8,56,57])
(a)The variation in oxygen, sulfide and pH within a microbial mat over a 24 h period. Profiles I and II represent two geochemical "snapshots" taken at 2pm and 3am that show key differences in depth profiles between day and night. As soon as the dark period starts, the photosynthesis ceases and the mat turns completely anoxic because of rapid O2 consumption by aerobic heterotrophs (modified after references[8,56]). (b) The six major guilds of microorganisms that compose a typical microbial mat are arranged by their respective effects on the precipitation process. Photosynthesis and sulfate reduction are known to increase alkalinity (promoting carbonate precipitation), whereas aerobic respiration, sulfide oxidation and fermentation are more likely to induce dissolution. When oxygen-depending metabolisms stop during the night, anaerobic heterotrophy such as sulfate reduction prevails. The net carbonate precipitation depends on the balance between the different metabolic activities as well as their temporal and spatial variations (modified after references[8,57])
总结起来,促使酸钙沉淀更重要的原因通常是有机矿化作用,包含生物诱导矿化和生物影响矿化2个过程[8]。生物诱导的有机矿化作用中,最重要的机制是蓝细菌等的光合作用和硫酸盐还原细菌(Sulfate-Reducing Bacteria,SRB)的硫酸盐还原反应导致的“碱度发动机”:在微生物席顶部含氧区域,蓝细菌等通过光合作用吸收CO2和/或HC
在对微生物碳酸盐岩详细分类之前,首先应该考虑其在整个灰岩分类体系中的位置(鉴于微生物白云岩的争议性,本文暂不讨论)。由于灰岩的结构是沉积、成岩和生物等不同作用过程相互作用的产物,因而存在一个宽松的谱系,Wright[61]依据成因—结构将其划分为沉积作用类、生物作用类和成岩作用类(图5)。其中沉积作用类可以根据Folk[62]或Dunham[63]的碳酸盐岩分类方案进行描述,生物作用类可以根据Embry等[64]的分类方案进行描述,成岩作用类则主要指重结晶的灰岩。作为微生物席钙化作用的产物,我们赞成梅冥相[4]的观点,微生物碳酸盐岩总体应该归类到生物作用类中的黏结岩中(结壳状构筑生物为主;图5)。
图5 灰岩的成因—结构分类(据参考文献[
Fig.5 The genetic-textural classification of limestones (modified after reference[61])
进一步,常采用4个不同的尺度来对微生物碳酸盐岩分类和描述[65]:①巨型构造,如生物丘,生物层等(图6a,b);②大型构造,几十厘米到几米,如柱状、穹窿状等(图6c);③中型构造,用于描述宏观上观察到的组构,如纹层状、凝块状和树枝状等(图6c~f);④微型构造,显微镜下才能观察到的微观组构,如胶结物、晶体形式和微生物组分等(图6g)。将这些不同尺度的构造特征相叠加,是微生物碳酸盐岩岩体单元的一个重要命名方法,如柱状叠层石生物丘,穹窿状均一石生物丘等。
图6 不同尺度下微生物碳酸盐岩的沉积构造及组构特征(华北地台寒武系)
(a)微生物生物层(白色箭头),寒武系徐庄组,河北省邢台市北会村;(b)微生物丘(白色箭头),寒武系徐庄组,河南省鲁山县;(c)(b)的局部放大,不显任何构造,由泥晶组成的均一石;(d)叠层石,箭头指向叠层石主体,寒武系凤山组,河北省井陉县;(e)凝块石,寒武系凤山组,河北省井陉县;(f)核形石(白色箭头),寒武系毛庄组,河北省邢台市北会村;(g)叠层石的显微照片,箭头指向微叠层状构造,寒武系凤山组,路通沟
Fig.6 Sedimentary structures and fabric of microbial carbonates in different scales (Cambrian of North China Platform)
(a) Microbialbiostrome (white arrows); Cambrian Xuzhuang Formation; Xingtai Beihui section, Hebei Province; (b) Microbial mound (white arrows);Cambrian Xuzhuang Formation; Lushan section, Henan Province; (c) A close up of (b), note leiolite without any macro-structure; (d) Stromatolite,and the arrows point to its column; Cambrian Fengshan Formation; Jingxing section, Hebei Province; (e) Thrombolite; Cambrian Fengshan Formation; Jingxing section, Hebei Province; (f) Oncoids (white arrows); Cambrian Maozhuang Formation; Xingtai Beihui section, Hebei Province; (g) Microstromatdite and the arrows pointing to micro-lamella;Cambrian Fengshan Formation; Lutonggou section
按照这一分类方案,微生物碳酸盐岩形成的大范围的岩石单元(巨型构造)可以分为微生物礁、微生物层和微生物丘(图5)。微生物礁和微生物丘都是具有穹形特征的微生物碳酸盐岩建隆,而微生物层不具有典型的地貌隆起,是横向延伸很远的层状微生物碳酸盐岩构成的岩体(图6a,b)。对于微生物礁的研究还不太成熟,比较肯定的是,微生物碳酸盐岩可以作为由小型藻类或后生动物构建的生物礁的组分或外来沉积物,但是否可以独立成礁还存在争议[2]。到寒武纪的时候,一些微生物碳酸盐组构的转变产生了一种似骨骼的骨架结构,这种骨架以含有树枝状组构(可能由直立的钙化蓝细菌丝状体产生)为特征,但现在就归为生物礁似乎还为时尚早,有待进一步深入研究[66,67]。
微生物丘多以丝状体的碳酸盐包壳或者细管状结构为主,有大量共性的特征,包括凝块的球粒状灰泥、层状晶洞构造、凝块石、叠层石、窗格孔和大量的纤状胶结物等(图6b~g)。在对微生物碳酸盐岩的描述和识别过程中,丘状、柱状和结壳等野外特征往往最为直观也极为重要。尤其是一些由均一石构成的微生物灰泥丘,或者以灰泥为主具有穹形特征的碳酸盐岩建隆,岩性总体为致密泥晶,在镜下无法判断这些灰泥究竟是生物成因还是非生物成因[68](图6b,c)。
相比较为简单的巨型和大型构造,微生物碳酸盐的中型和微型组构复杂且难以解释,尤其是微型组构。成分和结构是理解微生物碳酸盐岩所涉及的生物和形成过程的关键,但多数微生物碳酸盐岩镜下都以微晶结构为主导,明显的均一性掩盖了一系列可能的起源[11,43],只有一些捕获的颗粒、钙化的蓝细菌和包壳结构是最可能识别的成分[2]。所以,纹层的(叠层石)、凝块的(凝块石)、树枝状的(树形石)和隐晶质的(均一石)等宏观组构特征(中型构造)成为微生物碳酸盐岩识别和分类的主要依据,应用起来也较为便利。目前应用最广的是Riding[2]按照宏观组构(中型构造)划分的叠层石、凝块石、树形石和均一石四大类,梅冥相[4]增加了层纹石和核心石将其变为六大类。但依据组构特征进行的分类和命名会导致一系列概念上的混乱,引起术语究竟是描述定义还是成因定义的争论。
叠层石被Kalkowsky[12]最初提出时,强调了2个范畴:纹层结构和生物成因。Walter[69]对这个概念进行了强化,认为这种纹层结构都是微生物成因,是微生物在生长和新陈代谢过程中产生的捕获、黏结或沉淀作用所致。Semikhatov等[70]则仅强调其纹层结构,对微生物是否参与了叠层石的形成并未回答。直至今天,对叠层石和叠层结构的起源依然争论不断,尤其是前寒武纪叠层石中极少发现钙化微生物化石[40,71]。但正如Awramik[71]所强调的那样,除非是更加可靠和直接的证据,否则不应该因为叠层结构(在理论上)可以形成于非生物的沉积过程,而减弱将叠层石作为一种微生物组构的信心,叠层石仍然被广泛当作37亿年前生命存在的可能证据之一[5,71]。
凝块结构是微生物碳酸盐岩中另外一种常见的中型组构,Aitken[72]依据这一特征建立了凝块石这个术语,认为它是与叠层石相关的隐藻构造。与此同时,前苏联地质学家将与叠层石相区别的黏结灰岩统称为微植石(microphytolite),包括凝块石与核心石2种[4]。杜汝霖[73]则因凝块石中类似于花纹的组构而称其为“花纹石”,还曾被命名为藻团块、葡萄石和巴哈马石等。直至今天,尽管凝块石的使用几乎已和叠层石一样广泛,但凝块组构比纹层组构更加广泛也更难定义。而且不同于叠层结构的原生成因,凝块结构既可能是原生的也可能是次生的:除微生物钙化作用或凝聚作用外,微生物的降解或者后生动物的扰动似乎也可以形成凝块石[2]。这种组构特征和起源的多样性,让凝块石作为一个术语在概念上存在不一致和不确定性,并常常引发困惑。
树形石以树枝状的凝块为特征[2],在鲁西地区寒武系第三统张夏组大量产出[49]。树枝石在风化面上可以看到隐约的树枝形状或迷宫状,树枝斑块通常向上变宽,且排列规则,抛光面呈模糊的不规则凝块状[67,74]。实际上,树枝石和凝块石之间的界限模糊,难以辨认[74]。Riding[2]也说树形石的中型组构以凝块状组构出现,但又把树形石作为微生物碳酸盐岩中独立的一类和凝块石并列,这难免让人困惑。就目前的研究来看[2,66,67,74],树形石本质上与凝块石难以区别,应当归于凝块石。
核形石常常与凝块石共生,但形状为较规则的球形或椭球型而不同于凝块石(图6)。核形石的基本结构包括核心和外层2个部分,钙质外层为非同心的和部分重叠的纹层,常常被解释成生物起源,因而也一度被归类到叠层石,认为是球状叠层结构[14]。Riding[2]将其纳入次要的微生物碳酸盐岩,而梅冥相[4]则认为核形石有较为广泛的发育和特殊的组构,是一种典型的微生物碳酸盐岩类型。但究其术语本身,同叠层石等一样,核形石是纯粹描述性的名称术语,并不包含形成机制和环境意义[11]。
层纹石和均一石的研究就更为困难。层纹石以发育纹理化构造为特征,这种水平状纹理延伸极远,层理清晰,较稳定(跟叠层石区别)。纹理化构造单个纹层的厚度多在0. 5~1. 5 mm,被初步解释为未受到改造的席底构造,类似于“有机纹理”,形成于较深的水体环境[4]。均一石是指那些不显构造,宏观组构呈现隐晶状态的微生物碳酸盐岩[2,4]。
微生物碳酸盐岩微型构造的矿物相非常简单,往往以泥晶为主,但其微型组构却极其多样化,一个典型代表就是“石松藻状”(Lithocodium)钙化蓝细菌残余物,无论在叠层石还是凝块石中都有发现(图7)。石松藻(Lithocodium)最初被Elliott发现并解释为一种绿藻[75],后来被Schmid等[76]解释为一种结壳多孔的有孔虫,Cherchi等[75]解释为钙化蓝细菌的聚集物,以及近来又被解释为海绵骨针的网状物[66,67]和钙化蓝细菌菌落[77],泥晶为主的组分和不规则的形态让Lithocodium的解释和归类成了难题。
图7 微观尺度下的钙化(蓝细菌)微生物席(Lithocodium)
(a)左下角为Lithocodium,可见饼状菌或肾形菌化石(白色箭头),右上角主要为致密球粒状泥晶构成的附枝菌,枝晶结构已经很模糊(可能是硫酸盐还原细菌降解所致),两者之间没有明显的界限;(b)棒状或球粒状致密泥晶构成的附枝菌,趋近(a)取样点,进而推断了(a)右上角的蓝细菌类型;(c)不规则分布的丝状蓝细菌的残余物(亮晶方解石),呈网状散布在泥晶基质中,较多的黄铁矿(白色箭头)表明了可能的硫酸盐还原细菌的活动;(d)现象同(c),隐约可见管丝状蓝细菌化石(白色箭头);(a)~(d)都为单偏光镜显微照片;(a)和(b)为寒武系凤山组,辽宁金州湾剖面,(c)为寒武系张夏组,河北秦皇岛驻操营剖面,(d)为寒武系长山组,吉林郭家店剖面
Fig.7 Calcified (Cyanobacteria) microbial mats in microcosmic scale (Lithocodium)
(a) The lower-left corner is Lithocodium, note Tarthina (white arrows); the upper-right part is Epithyton with blurred branches (probably due to the degradation by sulfate-reducing bacteria), and there is no obvious boundary between the two; (b) Epithyton with rod or spherical shapes, the sample near (a), impies the type of cyanobacterial in the upper-right part of (a); (c) Irregularly distributed calcified cyanobacteria (sparite) are dispersed in the micrite, and pervasive pyrite (white arrows) indicates the possible activities of sulfate-reducing bacteria; (d) has similar phenomena with (c), note faintly visible tubular filamentous of cyanobacteria (white arrows); (a)and (b) is Cambrian Fengshan Formation, Liaoning Jinzhou Bay section,(c)is Cambrian Zhangxia Formation, Hebei Qinhuangdao Zhucaoying section,and (d) is Cambrian Changshan Formation, Jilin Guojiadian section
但正如Cherchi等[75]指出的那样,Lithocodium中缺乏连接相邻孔洞的开口和这些开口在大量泥晶基质中非常不规则的排列形态大大降低了其为石松藻和有孔虫的可能性(图7)。很多Lithocodium中海绵骨针的缺乏,管状菌(Tubomorphophyton)、胞网菌(Bacinella)和葛万菌(Girvanella)等蓝细菌丝状体的出现和短柱状亮晶沉积物(很可能是钙化蓝细菌残余物)在不规则泥晶团块和基质中的广泛出现也更加支持其是蓝细菌钙化微生物席沉积物[75,77](图7)。Lithocodium与纹层状、叠层状和凝块状一样,显示了它作为钙化微生物席沉积物的特征。弥散状EPS在降解和钙化作用过程中不会复制蓝细菌等微生物的丝状体或者鞘的外形和尺寸,这就是为什么微生物碳酸盐岩大多只可见纹层状、叠层状和凝块状以及“石松藻状”等构造,而很少出现蓝细菌等钙化微生物化石的原因。
在微生物席钙化作用过程中,大量呈弥散状分布的降解和钙化的EPS,以及一些形成在细胞附近的孤立碳酸钙晶体,会作为微米级别的碳酸盐泥沉淀在湖底或海底,构成Lithocodium的泥晶基质;少数厚鞘的蓝细菌或大气中的CO2值在临界点之下时,鞘被碳酸钙晶体浸染而得以保存,形成钙化蓝细菌化石;而大量鞘较薄或者不发育鞘的蓝细菌,在降解和钙化过程中只是作为核心沉淀形成了一些短柱状亮晶沉积物,不规则地散落在泥晶基质中(图7),形成了这种谜一样的钙化蓝细菌化石Lithocodium[77]。
总结起来,微生物碳酸盐岩在巨型—大型构造尺度下的特征较为简单和明显,常呈丘状、穹窿状、厚层状和柱状等形态;在小型构造尺度下,大多情况下以泥晶为主要组分,互相难以区分,所以目前的命名主要依据其宏观组构为依据,纹层、凝块(球粒)、均质是微生物碳酸盐岩进一步命名和分类的3个结构端元,叠层石、凝块石和均一石是最重要的也是最基本的类型。随着研究的深入,介于这些单元之间的一些分布较为广泛和具有独特宏观组构的其他微生物碳酸盐岩逐渐开始纳入这个命名体系。但在这个过程中,有关微生物碳酸盐岩的术语普遍具有不一致的问题(不同人的定义和理解都有所差别),甚至是令人迷惑不解的。也许就像Tucker等[11]所说,这些术语的混乱还将持续下去,甚至没有被理顺的一天。但有术语总比没有好,一个不变的和被普遍接受的科学名称可推进该化石更深层次的研究[78]。术语体系的复杂性正是微生物碳酸盐岩成因复杂性的客观反映,这些术语概念的演变,也是地质学家对微生物碳酸盐岩认识不断深化的过程。目前看来,将微生物碳酸盐岩置于Wright[61]分类方案的生物作用类之下,归类到Embry等[64]所描绘的黏结灰岩是可行的。进一步可以分为巨型构造、大型构造、中型构造(宏观组构)和微型构造(微观组构)4个尺度进行描述。其中按照宏观组构(中型构造)微生物碳酸盐岩可以分为叠层石、凝块石、核形石、层纹石和均一石5类,树形石的命名可以继续使用,但不应与其他5类并列而独立成类。
微生物碳酸盐岩具有显著的组构多样性,而且这种现象难以解释,很多时候只是笼统地解释为微生物席及其降解过程的复杂性所导致。与其复杂的沉积组构相反,微生物碳酸盐岩的矿物成分却非常简单,大部分主要由泥晶、微亮晶和亮晶构成(如叠层石、凝块石和均一石等),部分也以颗粒为主(如核形石等)。而且同一分类的微生物碳酸盐岩(如叠层石),可以形成于完全不同的环境,使得用环境来解释它们的成因变得困难。更甚的是,被认为是微生物成因的微生物碳酸盐岩中,很多时候其实并未发现微生物化石。这些矛盾的情况正是研究微生物碳酸盐岩艰难性的真实写照,也是其魅力所在。
形成微生物碳酸盐的微生物活动范围几乎包括了所有的环境,并以巨大的生物量和飞快的新陈代谢为特征,而我们对这种广泛参与沉积物生产、沉积及成岩过程的微生物知之甚少,使其成了名副其实的“生物学中沉睡的巨人”[28,79]。对微生物碳酸盐岩和微生物席沉积特性的研究是建立在对微生物,尤其是微生物席的生物学和生态学特性之上。因而微生物及微生物席沉积学等交叉学科应该受到重视。
目前微生物的分类比较复杂,而且不太确定,事实上多数微生物更适合作为“可疑微生物”来对待[43],因而对微生物碳酸盐岩的研究还有赖于微生物学的进展。例如在镜下观察到的纳米球究竟是什么东西,细菌?生物体腐烂产物?还是病毒或膜泡囊?目前比较广为接受的观点是,蓝细菌是现代微生物席的主要建造者,但绿色和紫色硫细菌构成的微生物群落,化能有机异养微生物、发酵微生物、化能有机自养微生物铁还原和硫酸盐还原细菌作为微生物席重要的功能性和结构性构成[20,80],所起的作用究竟多大?Castanier等[81]甚至发现在有机矿化作用导致的碳酸盐沉淀过程中,异养细菌的生产效率更高。一些现代微生物席中,硫酸盐还原反应也矿化了微生物席中全部有机碳的49%~63%[82]。此外,钙化微生物化石相对而言比较稀少,且不易保存。一般而言,管状、丝状和灌丛状构造是最具特征的细菌构造,但更多的时候只是一些斑斑点点的微晶凝块、平卧的纹层状沉积、被解体的球粒状泥晶和结壳状化微生物席沉积物,这极大地增加了鉴别和定义微生物碳酸盐岩的难度。
因而,对现代微生物席更加深入的研究是进行微生物碳酸盐岩研究的前提和条件。单个的微生物细胞或许是脆弱的,但它们相互协同构成的微生物群落——微生物席和微生物膜却有着广泛的适应能力。同一分类的微生物碳酸盐岩(如叠层石),可以形成于完全不同的环境,正是微生物席适应性的良好体现。而微生物席的一系列活动也会引起微生物碳酸盐岩沉积构造和组构的不同。有关叠层石纹层的成因和所代表的生物信号一直是研究的关键,归结起来大致有3种原因:①微生物席的自然属性所致,因为微生物席天然就是分层的,纹层就是本身生物群落的分层[34];②生态成因,指微生物席随着昼夜、四季的变更而形成的昼夜纹层或者季节性纹层[83];③聚合作用强调多种微生物的作用过程,诸如蓝细菌及EPS降解和钙化,以及微生物席下部硫化物的氧化作用和硫酸盐还原反应等共同导致了纹层的形成,并且随着石化作用微生物席原来的很多纹层将融合成一个单一的纹层[84]。这些研究表明,微生物席(膜)转化成叠层石不是一个直接的作用过程,而是多种因素综合作用的结果[39]。同样,叠层石石纹层的形态与微生物的新陈代谢活动以及组成微生物席/膜的EPS的降解作用也存在复杂的成因联系:①不连续凝块纹层可能是与微生物群落生长和腐烂相关的、不同时期的石化作用的产物;②网格状纹层可能为弱水流对生物席颗粒的簸选;③纹层中暗色丝状泥晶可能代表钙化了的EPS,或微生物产生的黏液或生物膜[85,86]。
各种各样的微生物群落相互协作构成了微生物席的纹层结构和复杂体系,甚至多重状微生物席。如前文所述,核形石常常与凝块石共生,但原因一直不明。对华北地台寒武系的微生物碳酸盐岩的研究发现,不仅核形石和凝块石,多种微生物碳酸盐岩共生的现象是较为普遍的,如叠层石与均一石、凝块石与均一石、叠层石与鲕粒、不同大小的鲕粒、鲕粒与核形石等(图8)。梅冥相[87]将产出于叠层石生物丘中的鲕粒称为“底栖鲕粒”,认为这些不均匀地散布在泥晶相叠层石中的放射状—纤维状鲕粒以其较小的粒径、多样的类型和平滑但不连续的泥晶化鲕粒圈层,以及外部边缘的泥晶套等特征,表现出较为明显的与微生物相关的微组构。这种关系仅仅用颗粒捕获作用是难以解释的,更何况两者都是微生物席钙化作用的产物,更有可能的情况是,这是多重状生物席(膜)的产物,凝块石常常与核形石共生就是一个鲜明的实例,而不同类型的微生物丘之间的交互生长更是多重生物席(膜)复杂性的体现(图8e)。
图8 华北地台寒武系中多种微生物碳酸盐岩的共生现象
(a)整体为均一石质微生物丘,球形,直径约2.2 m,寒武系崮山组,北京下苇甸;(b)为(a)的局部放大,可见叠层石和硬地(底)构造;(c)为(a)的局部放大,见凝块构造,在丘上可见清晰的缝合线;(d)为(a)的显微照片,在泥晶和微亮晶基质中可见溶解—充填结构的鲕粒;(e)均一石微生物丘和含鲕粒含生物碎屑微生物丘交互生长,寒武系徐庄组,河北省唐县;(f)核心石、粗鲕粒和细鲕粒共生现象,寒武系徐庄组,河南省济源市
Fig.8 Symbiosis of various microbial carbonates in Cambrian of the North China Platform
(a) A leiolite microbial mound, spherical, about 2.2 m in diameter, Cambrian Gushan Formation, Beijing Xiaweidian section; (b) A close up of (a), note stromatolite and hard ground; (c) A close up of (a), note mud clots and clear stylolite; (d) A micrograph of (a), note the ooid with a dissolved-filled structure distributed in the micrite and micro-sparite; (e) Symbiosis of leiolite microbial mound and oolitic-bioclastic microbial mound, Cambrian Xuzhuang Formation, Hebei Tangxian section; (f) Symbiosis of oncoids, coarse ooide and fine ooides, Cambrian Xuzhuang Formation, He'nan Jiyuan section
此外,微生物和微生物席的生态学研究为我们思考地质现象和认识地质规律提供了新线索和新思路。例如不论叠层石的纹层与昼夜循环有关、与季节变更有关还是与太阳黑子周期有关[14,88,89],这都将微生物碳酸盐岩的形成和天文节律相联系。因此,从数量级上查明替代指标变化周期或频率与这些天文周期或频率的一致性,就能用替代指标变化周期或频率确定地层的近似年限。而且,只需要选定特定的坐标单位谱(如岁差、斜度、偏心率周期等),建立与人类社会接轨的地质学时间坐标也成为可能,从而满足我们对年代地层精确性、严谨性的不懈追求。天文节律对地球的影响在整个地史时期应该均有记录,而且具有全球统一性,因而运用微生物碳酸盐岩分析缺乏宏观化石的显生宙之前的地层具有广泛前景。
微生物席的钙化是一个生物化学作用过程,是生物矿化和有机矿化作用的结果。通常来讲,生物矿化作用(如骨骼的形成)往往有利于生物的生存,但生物为什么要发生有机矿化,却是一个格外复杂的课题[41]。问题就在于,与微生物相关的钙化又主要是“非骨骼”或“骨骼外”的钙化,是有机矿化作用的结果。而且,尽管前文已经讨论了很多生物矿化作用和有机矿化作用的过程,但更多的细节和机理依然模糊不清,只是触及到冰山的一角[8]。
在所有问题中最引人入胜的莫过于“前寒武纪之谜”:叠层石是前寒武纪地层中最常见的化石,通常被认为是钙化微生物席的石化产物[34],但是直到今天,我们还未能在前寒武纪叠层石中找到钙化蓝细菌等微生物化石[6,40]。科学家们认为叠层石应该是生物成因主要依据于它奇特的形状和纹层,从简单的丘状到复杂的分支柱状、锥状,以及纹层状组构,仅仅靠机械沉积过程是无法解释的;叠层石在不同的环境和盐度下表现出活性,发育不同形状,也很难用非生物沉积过程解释[69,70]。但前寒武纪叠层石缺乏直接的微生物证据无疑给其微生物成因假说蒙上了阴影,显生宙之后的叠层石才见到钙化的微生物化石,是否与当时大气、海洋环境不利于蓝细菌鞘的钙化有关[44]?解决这个问题有待于微生物(席)钙化作用机制的进一步研究,而且意义重大。因为最早的叠层石是否为微生物起源牵扯到2个最基本的科学问题——光合作用和钙化作用[16]。
近期的研究表明,微生物碳酸盐岩的形成除蓝细菌等的光合作用和硫酸盐还原细菌的硫酸盐还原反应,以及碱度发动机和细胞外聚合物质的降解过程所驱动外,非晶质碳酸钙(Amorphous Calcium Carbonate,ACC)的形成可能是另外一个重要原因[87,90]。ACC是生物矿化所涉及的一种特殊的碳酸盐矿物形式,是一种较为典型的生物矿物的过渡型前体相[53,90,91]。在鲕粒外表皮ACC微薄层的存在,暗示了细菌活动所导致的局部微环境碱度的增加,以及EPS基质中胞外有机物质等成核中心的存在促使了ACC的沉淀。而纯净的ACC是高度不稳定的,只是作为更稳定的晶质文石和方解石的过渡型前体[90]。因而ACC的成因成为了关键,是促进碳酸钙沉淀作用的第一个阶段,如果ACC真是生物成因,无疑会增加人们对鲕粒等生物成因模型的信心[87]。
目前看来,有关微生物(席)钙化作用机制的研究还有很长的路要走。尽管地质学家们描述、表征和解释了各种各样的微生物碳酸盐岩,但只获得了一些推测性答案,一切仍然是个谜[68]。面对这个复杂且困难的钙化机制,微生物碳酸盐工厂将会是一个综合有效的分析方法。碳酸盐工厂就是像工厂那样生产碳酸盐的地方,即不管其中细节如何,可以将海洋等生产碳酸盐的地方看作一个整体[92,93,94]。同样,不管微生物碳酸盐的形成是多么复杂的生物矿化和有机矿化作用的过程,我们都可以将其视作一个整体来研究其对环境的响应。例如热带碳酸盐工厂[95]主要用来描述南北纬30°之间,热带浅水透光带的海洋这一沉积场所,在那儿碳酸盐沉积物的生产作用与海洋植物和微生物的光合作用活动紧密联系,虽然不知道这些海洋生物生成碳酸盐的具体细节和机制,我们依然可以分析其动态:这儿碳酸盐的产率随着水体深度的增加而呈指数式的降低,当水体深度超过100 m左右时碳酸盐产率几乎降为零[94,95]。更为重要的是,尽管碳酸盐工厂可以呈“黑匣子”的形式来预测碳酸盐岩的生产、堆积作用过程,但并不妨碍对碳酸盐沉积作用机制的研究。相反,对沉积机制了解的越多,碳酸盐工厂就更能以清晰的面貌展现出来。
同一分类的微生物碳酸盐岩,可以形成于完全不同的环境。在南极冰川湖泊中、新西兰热水体系中、巴哈马台地和澳大利亚鲨鱼湾等热带、亚热带海域,都有现代叠层石的形成[39,85]。核形石也出现在湖泊、海洋等各种各样的环境,甚至河流和土壤中[74]。这使得用环境来解释微生物碳酸盐岩的成因或者用微生物碳酸盐岩来作为某种沉积环境的指示器都变得困难。尽管如此,微生物碳酸盐岩依然可以反映和解释古地理环境,只是依靠它的构造形态、宏微观组构和(尤其是)钙化作用特征:由于微生物席的生命活性,它们在不同环境下会调整自己的形态和结构以适应环境的要求和变化;随着时间的推移和海水化学性质的波动,蓝细菌钙化作用的形式和微生物丘的形态也发生着相应的变化[2,44]。
值得注意的是,用微生物碳酸盐岩的沉积构造和组构进行环境解释时,不得不面对它的形态和组构是受环境控制还是受微生物群落控制的问题。长期以来,都把叠层石的形态特征与水动力条件相联系,例如在潮汐环境下,一般认为近水平缓波状叠层石往往在水动力弱的潮上带形成,缓波状叠层石主要分布于水动力弱的潮间带,柱状叠层石集中在水动力强的潮间带,而锥状和半球状叠层石则形成于水动力很强的潮间带[89]。但实际情况要复杂得多,最近在南极常年被冰覆盖的Untersee湖中就发现了小尖塔状叠层石和大型锥状叠层石,并认识到生物学行为是造成这种泥质叠层石形态学特征的主要起因[86]。Playford[96]早在1990年就提出过叠层石的不同形态主要是不同微生物群落所致,并将研究区微生物席分成胶粒状、凝胶状、光滑形态、针垫状、毛簇状、脓疱状、薄膜状、网状和小气泡状9种。
就像在一种极端淡水中看到的泉华叠层石与另一种极端寒冷海洋水域中浅海丘状叠层石的难以对比一样,着眼于更长尺度的地质历史可能有助于我们获得更可靠的结论,因而海洋—大气圈系统的长周期震荡和气候变化对微生物碳酸盐岩矿物学组分和沉积组构的效应是一个重大而有趣的课题。Riding[2,44]从微生物碳酸盐岩及过去海洋中微生物碳酸盐岩的丰度随着时间变化推断环境的控制高于一切。从叠层石长期演变来看,太古代以亮晶叠层石为主,新元古代以后则以泥质叠层石为主(图9)。在寒武纪—早奥陶纪、晚泥盆纪和中晚三叠纪发生了3次明显的蓝细菌钙化作用事件(Cyanobacterial Calcification Events,CCEs),蓝细菌钙化幕似乎增强了海相碳酸盐的沉淀,也大致与鲕粒和海洋胶结物不寻常的含量周期一致[2,44]。这可能与大气圈CO2的浓度(图9)、全球气候(高温可以提高碳酸盐沉淀率)以及来自分层盆地碱泵的产生等有着密切的联系[2,44,47]。
图9 大气中CO2水平与微生物碳酸盐岩的组构演化及蓝细菌鞘的出现(据参考文献[
Fig.9 The CO2 level in the atmosphere, the fabric evolution of microbial carbonates and first appearance of calcified cyanobacteria sheath (modified after reference[35])
微生物碳酸盐岩有着最长的地质历史和最广泛的分布(跟所有的生物成因的碳酸盐岩相比较),也是地球上最早的生命证据,对整个地质历史时期与微生物活动相关的生物/有机矿化作用过程都提供了有用信息。但这个过程中最重要的钙化作用——同沉积钙化作用并不完全由微生物自身控制,而是微生物的进化、与后生动物的竞争,以及环境因素共同作用的结果,而且微生物活动往往和环境是协同的,是互相影响的。所以对微生物碳酸盐岩沉积环境的解释过程中,单一的强调某种因素难免有失偏颇。更为合理的解释需要建立在2个基础之上:一是进一步调查钙化微生物席的地质记录,结合现代微生物席沉积特征(如钙化蓝细菌CCMs研究),形成概括性的概念;二是对微生物碳酸盐岩的研究应该置于更详细的具体的环境,尽可能建立特定环境下不同因素对其沉积组构和钙化作用的贡献参数。在此之上,根据微生物碳酸盐岩的构造形态和宏微观组构解释沉积环境,通过微生物的钙化作用反映海水化学性质和大气成分的变化,以及这些环境中微生物(席)结构和生理的变化一定能够成为可能。
微生物自身、微生物产生和分泌的EPS以及捕获或者沉淀的外来物质共同构成了微生物席(膜)。微生物席(膜)是一个复杂的有机整体,构成了地球上最早的生态系统并持续至今,深刻影响着地球的大气圈、水圈和岩石圈的演变。微生物席可以在不同的纹层段创造出较宽泛的环境微梯度,捕获、黏结和沉淀碳酸盐。微生物碳酸盐岩就是钙化微生物席(膜)石化作用的产物。
微生物碳酸盐岩的形成主要取决于微生物席的2个主要作用过程,一是颗粒捕获及黏结,二是矿化作用及矿物沉淀。颗粒的捕获和黏结在局部可能是重要的,但最重要的还是矿化作用和矿物的沉淀,主要形成于以下3个机制:①碱度发动机和细胞外聚合物质的降解过程所驱动;②蓝细菌等光合作用及硫酸盐还原细菌的硫酸盐还原作用;③非晶质钙碳酸盐ACC的产生和转变。
微生物碳酸盐岩可以归类到Embry等所描绘的黏结岩(基于Wright在1992年的成因—结构分类),进一步可以按照巨型构造、大型构造、中型构造(宏观组构)和微型构造(微观组构)4个尺度进行描述。其中按照宏观组构(中型构造)微生物碳酸盐岩可以分为叠层石、凝块石、核形石、层纹石和均一石5类,树形石的命名可以继续使用,但应归于凝块石而不应与其他5类并列。
微生物碳酸盐岩被誉为“最难研究的沉积岩之一”,有几个鲜明的特点:尽管微生物碳酸盐岩的矿物成分非常简单,但却有显著的组构多样性;被认为是微生物成因的微生物碳酸盐岩中很多时候其实并未发现微生物化石;同一分类的微生物碳酸盐岩(如叠层石),可以形成于完全不同的环境,使得用环境来解释它们的成因变得困难。这些现象尽管都可以笼统地归为微生物席及其降解过程的复杂性所导致,但更多的细节有待进一步研究。微生物,尤其是微生物席的沉积学特性和钙化作用机制对于理解微生物碳酸盐岩的形成至关重要。
The authors have declared that no competing interests exist.
| [1] |
Microbialites, organosedimentary deposits of benthic microbial communities [J].
Microbialites are organosedimentary deposits formed from interaction between benthic microbial communities (BMCs) and detrital or chemical sediments. Processes involved in the formation of calcareous microbialites include trapping and binding of detrital sediment (forming microbial boundstones), inorganic calcification (forming microbial tufa), and biologically influenced calcification (forming microbial framestones). The latter process is probably the result either of chemical changes associated with photosynthesis, or the nucleation of crystals on the polysaccharide-bearing sheaths of the microorganisms. Elevated 未13C values in these crystals may reflect isotopic fractionation associated with the biological setting of the mineralization. Microbialites contrast with other biological sediments in that they are generally not composed of skeletal remains. Once formed, the primary framework of the microbialite becomes the locus of secondary cementation. To distinguish them from bioherms and biostromes of skeletal origin, microbialite buildups are termed "microbial lithoherms" or "microbial lithostromes". The morphogenesis of these structures is a function of environmental influence, biologic and ecologic controls, and processes and rates of lithification. The internal structures of microbialites are best identified by descriptive terms that do not imply a particular origin. Terms used to describe internal structures of microbialites include stromatolitic, thrombolitic, oncolitic (concentrically laminated), spherulitic, and cryptic. Consideration of the term "stromatolite" shows that it is currently used in at least three distinct ways: to refer to products of microbial sedimentation in general, to describe laminated structures of probable microbial origin, or to describe discrete laminated lithified bodies. We recommend that the term be restricted to refer to microbialites with an internal structure of fine, more or less planar laminations. Assessment of published distinctions between thrombolites (microbialites characterised by a clotted internal structure) and stromatolites in the light of some present-day Australian occurrences shows that thrombolites are not always constructed by coccus-dominated BMCs, nor are stromatolites always constructed from filament-dominated BMCs. Although we have observed examples of modern thrombolitic structures forming where biogenically influenced calcification dominates, and stromatolitic structures forming where trapping and binding of either detrital sediment or seasonally precipitated carbonate dominates, both structures could be produced by other processes. It is hoped that the ability to differentiate between the processes involved in the genesis of microbialites will lead to a better understanding of such factors as the morphogenesis of microbialites, their evolution through geologic time, and their potential as tools for biostratigraphic correlation.
|
| [2] |
Microbial carbonates: The geological record of calcified bacterial-algal mats and biofilms [J]. |
| [3] |
Structure and composition of organic reefs and carbonate mud mounds: Concepts and categories [J].
Defined here as ‘essentially in place calcareous deposits created by sessile organisms’, Organic Reefs are diverse and complex structures with a long geological history. Their classification has been the subject of fierce debate, often characterized by reliance on subjective features such as wave-resistance and qualitative attempts to discriminate between ‘first’ and ‘second class’ reefs. In contrast, emphasis is here placed on the objective characteristic of the type of sedimentary support, which largely determines the sedimentary composition of the deposit. Constructional and depositional processes result in three principal sedimentary components: matrix (M), essentially in place skeletons (S) and cavity/ cement (C), whose proportions can be represented on MSC triangular plots. Separately or together, these components also provide the structural support for the reef. On these compositional and structural bases, three main categories of Organic Reef are recognized: 1. Matrix-supported reefs (Agglutinated Microbial Reefs, Cluster Reefs, Segment Reefs), 2. Skeleton-supported reefs (Frame Reefs), 3. Cement-supported reefs (Cement Reefs). Agglutinated Microbial Reefs: possess laminated, clotted, or aphanitic fabrics created by microbial trapping of particulate sediment; in place skeletons and large primary cavities are rare; early cementation may provide added support; topographic relief is limited by the need for currents to provide sediment to accreting surfaces. Cluster Reefs: skeletal reefs in which essentially in place skeletons are adjacent, but not in contact, resulting in matrix support; characterized by relatively high matrix/skeleton ratios and low volumes of extra-skeletal early cement. Sediment trapping is an important corollary of skeletal growth and Cluster Reef organisms are tolerant of loose sediment. Absence of framework limits the topographic relief that Cluster Reefs can attain relative to spatial extent, and may permit bedding to develop within the reef. Close Cluster Reefs have skeletons up to 1 unit-distance apart. Spaced Cluster Reefs have skeletons more than 1, and up to 2 unit-distances apart; with increasing separation of skeletons they grade to level-bottom communities. Segment Reefs: matrix-supported reefs in which skeletons are adjacent, and may be in contact, but are mostly disarticulated and mainly parauthochtonous. Matrix abundance is high, and early cement relatively low. Moderate relief can develop in response to intense on-reef sediment production. Frame Reefs: skeletal reefs in which essentially in place skeletons (including calcified microbes) are in contact; characterized by relatively high skeleton/matrix ratio. Skeletal support enables them to raise themselves above the substrate independently of cementation and particulate sedimentation. Simultaneously, by creating partly open shelter cavities, skeletal support may facilitate early cementation. Both relief and early lithification promote marginal talus formation. Skeletal shape and orientation distinguish: conical/stick-like, dendritic, domical, and laminar frames. Each of these may be open or filled. Open Frame Reefs: cavities remain open during the early stages of reef growth and are occupied by cryptic encrusters, early cements and internal sediment; exposed skeleton encourages endoliths. Filled Frame Reefs: inter-skeletal spaces penecontemporaneously occluded by surficial sediment during reef-growth. Cement Reefs: reefs created by cementation of essentially in place organisms. Cement provides strength and volume, mimicking skeletal growth, and can form on non-skeletal as well as skeletonized organisms. Non-skeletal Cement Reefs: created by synsedimentary cementation of essentially in place non-skeletal organisms. This converts a soft deposit with relatively poor preservation potential into a rigid lithified mass: e.g., Tufa Cement Reefs (phytoherms) in rivers and lakes and possibly Travertine Cement Reefs associated with hot springs. If the organisms are skeletal, synsedimentary cementation imparts extra strength and stability to what otherwise would be a Cluster or Frame Reef, and results in Skeleton–Cement Reefs. Cement Reefs exhibit complex relationships between cement, matrix and skeletons. Agglutinated Microbial, Cluster and Segment reefs tend to be structurally simple, have low primary relief, and may show bedding. Frame (including microbial Microframe) and Cement Reefs tend to be unbedded, structurally complex, and can have high relief. Carbonate Mud Mounds: carbonate mud-dominated deposits with topographic relief and few or no stromatolites, thrombolites or in place skeletons. Low Relief Carbonate Mud Mounds are typically thin. High Relief Carbonate Mud Mounds are thick, and internal bedding, slumping, stromatactis cavity systems, and steep marginal slopes may be common. Whereas Organic Reefs are biogenic, calcareous, and are created by essentially in place organisms, Carbonate Mud Mounds can be organic and/or inorganic in origin and it can be difficult to distinguish their origins.
|
| [4] |
Revised classification of microbial carbonates: Complementing the classification of limestones [J].微生物碳酸盐岩分类体系的修订:对灰岩成因结构分类体系的补充 [J].
微生物沉积作用在前寒武纪地层中普遍发育,在显生宙的一些地层中也较为发育。在碳酸盐岩地层之中,以叠层石为代表的微生物岩尤为引人注目。经过长期研究,2000年Riding曾经将微生物碳酸盐岩分为叠层石、凝块石、树形石和均一石4大类型。实际上,核形石以其较为广泛的发育和特殊的微组构也应该作为一种典型的微生物碳酸盐岩类型而纳入微生物碳酸盐岩的分类体系之中,而不能简单地作为球状叠层石。而那些纹理石灰岩,较厚的纹理和较深的产出沉积环境与叠层石形成明显的区别,也应该作为一种微生物碳酸盐岩的类型。生物沉积作用所形成的碳酸盐岩,以生物礁岩最为典型,在20世纪70年代曾经被Embry和Kloven归为骨架岩、障积岩、粘结岩三大类型,后来又增加了胶结岩,这是对20世纪50年代Folk、Dunham关于灰岩成因结构分类体系的良好补充。这些生物礁岩石以其高能量形成环境而有时又几乎见不到颗粒而与"颗粒含量越高沉积环境的能量越高"的基本理念不相符,所以Wright在1992年将它们归为生物作用类岩石,从而将灰岩划分为沉积作用、生物作用、成岩作用三大类。根据该分类,Folk和Dunham所描述的分类则属于沉积作用类灰岩,而Embry和Kloven所描述的生物礁岩石则归为生物作用类灰岩。微生物碳酸盐岩,总体上构成生物作用类碳酸盐岩中的粘结岩类,以其明显的微生物作用特点而具有自己的分类体系;它不但作为生物礁岩石的主要类型,而且也常常以生物礁、生物层和生物丘三种形式发育在地层之中。因此,上述概念和认识的进步,在强调微生物沉积作用的重要性的同时,有必要将微生物碳酸盐岩重新分为6大类:叠层石、凝块石、核形石、树形石、纹理石和均一石。
|
| [5] |
Rapid emergence of life shown by discovery of 3,700-million-year-old microbial structures [J].
Biological activity is a major factor in Earth's chemical cycles, including facilitating CO2 sequestration and providing climate feedbacks. Thus a key question in Earth's evolution is when did life arise and impact hydrosphere-atmosphere-lithosphere chemical cycles? Until now, evidence for the oldest life on Earth focused on debated stable isotopic signatures of 3,800-3,700 million year (Myr)-old metamorphosed sedimentary rocks and minerals1,2 from the Isua supracrustal belt (ISB), southwest Greenland3. Here we report evidence for ancient life from a newly exposed outcrop of 3,700-Myr-old metacarbonate rocks in the ISB that contain 1-4-cm-high stromatolites-macroscopically layered structures produced by microbial communities. The ISB stromatolites grew in a shallow marine environment, as indicated by seawater-like rare-earth element plus yttrium trace element signatures of the metacarbonates, and by interlayered detrital sedimentary rocks with cross-lamination and storm-wave generated breccias. The ISB stromatolites predate by 220 Myr the previous most convincing and generally accepted multidisciplinary evidence for oldest life remains in the 3,480-Myr-old Dresser Formation of the Pilbara Craton, Australia4,5. The presence of the ISB stromatolites demonstrates the establishment of shallow marine carbonate production with biotic CO2 sequestration by 3,700 million years ago (Ma), near the start of Earth's sedimentary record. A sophistication of life by 3,700 Ma is in accord with genetic molecular clock studies placing life's origin in the Hadean eon (>4,000 Ma)6.
|
| [6] |
Stromatolites in Precambrian carbonates: Evolutionary mileposts or environmental dipsticks? [J].
Abstract Stromatolites are attached, lithified sedimentary growth structures, accretionary away from a point or limited surface of initiation. Though the accretion process is commonly regarded to result from the sediment trapping or precipitation-inducing activities of microbial mats, little evidence of this process is preserved in most Precambrian stromatolites. The successful study and interpretation of stromatolites requires a process-based approach, oriented toward deconvolving the replacement textures of ancient stromatolites. The effects of diagenetic recrystallization first must be accounted for, followed by analysis of lamination textures and deduction of possible accretion mechanisms. Accretion hypotheses can be tested using numerical simulations based on modem stromatolite growth processes. Application of this approach has shown that stromatolites were originally formed largely through in situ precipitation of laminae during Archean and older Proterozoic times, but that younger Proterozoic stromatolites grew largely through the accretion of carbonate sediments, most likely through the physical process of microbial trapping and binding. This trend most likely reflects long-term evolution of the earth's environment rather than microbial communities.
|
| [7] |
Microbial carbonates in space and time: Introduction [J].
Loeb space methods are used to prove existence of an optimal control for general 3D stochastic Navier–Stokes equations with multiplicative noise. The possible non-uniqueness of the solutions mean that it is necessary to utilize the notion of a non-standard approximate solution developed in the paper by N.J. Cutland and Keisler H.J. 2004, Global attractors for 3-dimensional stochastic Navier–Stokes equations, Journal of Dynamics and Differential Equations, pp. 16205–16266, for the study of attractors.
|
| [8] |
Processes of carbonate precipitation in modern microbial mats [J].DOI URL [本文引用: 11] 摘要
Microbial mats are ecosystems that arguably greatly affected the conditions of the biosphere on Earth through geological time. These laminated organosedimentary systems, which date back to > 3.4 Ga bp, are characterized by high metabolic rates, and coupled to this, rapid cycling of major elements on very small (mm- m) scales. The activity of the mat communities has changed Earth's redox conditions (i.e. oxidation state) through oxygen and hydrogen production. Interpretation of fossil microbial mats and their potential role in alteration of the Earth's geochemical environment is challenging because these mats are generally not well preserved. Preservation of microbial mats in the fossil record can be enhanced through carbonate precipitation, resulting in the formation of lithified mats, or microbialites. Several types of microbially-mediated mineralization can be distinguished, including biologically-induced and biologically influenced mineralization. Biologically-induced mineralization results from the interaction between biological activity and the environment. Biologically-influenced mineralization is defined as passive mineralization of organic matter (biogenic or abiogenic in origin), whose properties influence crystal morphology and composition. We propose to use the term organomineralization sensu lato as an umbrella term encompassing biologically influenced and biologically induced mineralization. Key components of organomineralization sensu lato are the lkalinity engine (microbial metabolism and environmental conditions impacting the calcium carbonate saturation index) and an organic matrix comprised of extracellular polymeric substances (EPS), which may provide a template for carbonate nucleation. Here we review the specific role of microbes and the EPS matrix in various mineralization processes and discuss examples of modern aquatic (freshwater, marine and hypersaline) and terrestrial microbialites.
|
| [9] |
Feature and nature of microbial-mat: Theoretical basis of microbial-mat sedimentology [J].微生物席的特征和属性:微生物席沉积学的理论基础 [J].
微生物席是一个特别的微生物群落,构成了地球上形成最早、延续时间最长的生态系,这已经成为共识。但是,由于微生物席生物构成的复杂性、发育环境和产物的多样性,对微生物席的属性还存在不同的答案。不同的答案意味着不同的研究领域,因为对一个基本问题,即“究竟什么是微生物席”,要获取一个单一的答案几乎是不可能的。微生物席定义的多重性答案,最终就成为地球生物学框架下一个年轻的沉积学分支——“微生物席沉积学”的概念体系和理论基础。最为重要的是,要穿越成岩作用“过滤器”,去了解地质历史时期的微生物席建造物的生长机理和发育过程,还存在若干的困难,这些困难表现在几个重要的现象之中,如:(1)寒武纪叠层石中的底栖鲕粒;(2)中元古界灰岩层面上特别的MISS(微生物诱发的沉积构造);(3)中元古界叠层石白云岩序列中的凝块石。因此,基于微生物席的属性和特征所蕴含的微生物席的多样化定义,介绍微生物席沉积学的理论体系,对于深入理解这一年轻分支学科的现状和进一步发展具有特别重要的意义。
|
| [10] |
Review and prospectives of microbial carbonate reservoirs [J].微生物碳酸盐岩油气储层研究现状与展望 [J].
微生物碳酸盐岩是一种重要的油气储集岩。在中国古老深埋碳酸盐岩地层中它们是主要的岩石类型。微生物碳酸盐岩通常可划分为叠层石、凝块石、树枝石、均一石和核形石等五种类型。储层孔隙系统与微生物岩的沉积结构和构造密切相关,窗格孔(洞)和格架孔(洞)是主要储集空间类型。全球中新元古界至中生界都发现了以微生物碳酸盐岩为储层的油气田,资源潜力巨大。 微生物岩储层的岩石类型、沉积构造、相序结构、沉积模式和储层有利相带尚有许多不明晰之处,是今后进一步深化研究的重点方向。
|
| [11] |
Carbonate sedimentology [J]. |
| [12] |
Oolith and Stromatolith im norddeutschen Buntsandstein [J]. |
| [13] |
Journal of researches into the geology and natural history of the various countries visited by H.M.S. Beagle [J].
ABSTRACT Journal of researches into the geology and natural history of the various countries visited by H.M.S. Beagle / Charles Darwin Note: The University of Adelaide Library eBooks @ Adelaide.
|
| [14] |
Classification and environmental significance of algal stromatolites [J].
Stromatolites are laminated structures that have been previously termed fossil algae. It is now recognized that such structures may be formed by a number of different processes and organisms. Recognizable algal stromatolites are more validly treated as organosedimentary structures than fossil organisms. This paper proposes a new classification of algal stromatolite structures based upon their geometric forms and relates these forms to their sites of growth relative to sea-level environments. The classification uses the arrangement of the basic geometric units (hemispheroids and spheroids) from which common stromatolites and oncolites are built. Three main arrangements of these geometric structures occur in Recent algal stromatolites and oncolites; (1) laterally linked hemispheroids (LLH), (2) discrete, vertically stacked hemispheroids (SH), and (3) discrete spheroids-either as randomly stacked hemispheroids or concentrically arranged spheroids (SS). The abbreviations are used to designate the various structures conveniently. Since certain restricted environments are associated with modern stromatolites, ancient environments may be interpreted by recognition of fossil stromatolite forms. For example, protected intertidal mud flats, where wave action is slight, may be inferred by the presence of type-LLH stromatolites (Collenia). Exposed, intertidal mud flats, where scouring action of waves and other interacting factors prevent growth of algal mats between stromatolites, are inferred by the presence of type-SH structures (Cryptozoon). Low intertidal areas that are exposed to waves and agitated shallow water below low-water mark are inferred by the presence of type-SS structures (oncolites). In addition, the arrangement of laminae in these structures reflects frequency of movement, frequent motion resulting in concentrically arranged spheroids and occasional motion in randomly stacked hemispheroids.
|
| [15] |
Biology of Microorganisms(7th edition) [M]. |
| [16] |
Photosynthetic microbial mats in the 3,416-Myr-old ocean [J].
Recent re-evaluations of the geological record of the earliest life on Earth have led to the suggestion that some of the oldest putative microfossils and carbonaceous matter were formed through abiotic hydrothermal processes. Similarly, many early Archaean (more than 3,400-Myr-old) cherts have been reinterpreted as hydrothermal deposits rather than products of normal marine sedimentary processes. Here we present the results of a field, petrographic and geochemical study testing these hypotheses for the 3,416-Myr-old Buck Reef Chert, South Africa. From sedimentary structures and distributions of sand and mud, we infer that deposition occurred in normal open shallow to deep marine environments. The siderite enrichment that we observe in deep-water sediments is consistent with a stratified early ocean. We show that most carbonaceous matter was formed by photosynthetic mats within the euphotic zone and distributed as detrital matter by waves and currents to surrounding environments. We find no evidence that hydrothermal processes had any direct role in the deposition of either the carbonaceous matter or the enclosing sediments. Instead, we conclude that photosynthetic organisms had evolved and were living in a stratified ocean supersaturated in dissolved silica 3,416 Myr ago.
|
| [17] |
Hydrogen-based carbon fixation in the earliest known photosynthetic organisms [J]. |
| [18] |
Hydrothermal systems: Doorways to early biosphere evolution [J]. |
| [19] |
Atmospheric composition and climate on the early Earth [J].
Oxygen isotope data from ancient sedimentary rocks appear to suggest that the early Earth was significantly warmer than today, with estimates of surface temperatures between 45 and 85°C. We argue, following others, that this interpretation is incorrect-the same data can be explained via a change in isotopic composition of seawater with time. These changes in the isotopic composition could result from an increase in mean depth of the mid-ocean ridges caused by a decrease in geothermal heat flow with time. All this implies that the early Earth was warm, not hot. A more temperate early Earth is also easier to reconcile with the long-term glacial record. However, what triggered these early glaciations is still under debate. The Paleoproterozoic glaciations at approximately 2.4 Ga were probably caused by the rise of atmospheric O60 and a concomitant decrease in greenhouse warming by CH62. Glaciation might have occurred in the Mid-Archaean as well, at approximately 2.9 Ga, perhaps as a consequence of anti-greenhouse cooling by hydrocarbon haze. Both glaciations are linked to decreases in the magnitude of mass-independent sulphur isotope fractionation in ancient rocks. Studying both the oxygen and sulphur isotopic records has thus proved useful in probing the composition of the early atmosphere.
|
| [20] |
What are microbial mats? [M] |
| [21] |
Frommat ground structures to the primary sedimentary structures of a fifth category: Significant concepts on sedimentology [J].从席底构造到第五类原生沉积构造:沉积学中具有重要意义的概念 [J].
<p>发育微生物席或微生物膜的沉积底被形象地称为席底,在席底上由于微生物席的发育使沉积物富黏性而抗水流改造,并在沉积面和沉积层内形成一系列奇形怪状的原生沉积构造,如变余波痕、皱饰构造、微生物席碎片、纺锤状或次圆状收缩裂缝等等,这些构造都归为席底构造;与席底相对应,那些不发育微生物席或微生物膜的沉积面则称为混合底,在混合底上单个沉积颗粒受到水流剪切与变形作用的改造而形成诸如正常波痕等原生沉积构造,这些沉积构造则归为混合底构造。席底和席底构造的概念很重要,用来专指微生物沉积作用的产物而区别于混合底和混合底构造。从沉积学概念的演变方面分析,混合底构造就是1964年Pettijohn和Potter所定义的原生沉积构造,他们曾经将其归为4大类并且沿用至今;席底构造也就是微生物形成的(或相关的)原生沉积构造,由于其在形成机理和形态上与Pettijohn和Potter所定义的原生沉积构造存在较大的区别,Noffke 2001年认为其应该具有合适的分类位置,所以就将席底构造归为“第五类原生沉积构造”并纳入Pettijohn和Potter 1964年的原生沉积构造的分类体系之中。从席底构造到第五类原生沉积构造,概念的演变表明了万能的微生物在沉积作用过程中的重要性以及沉积学家对微生物沉积作用认识的深入。这些相关的沉积构造及其所形成的席底,就像叠层石(被当作正生长原生沉积构造)一样在前寒武纪地层之中较为普遍。随着后生动物的大规模出现而造成的寒武纪“农艺革命”(Agronomic revolution)的开始,显生宙的席底及其相关的席底构造多出现在那些不适宜后生动物居住的地方并大多被混合底及其相关的混合底构造所替代;因此,与微生物相关的原生沉积构造,除熟知的叠层石以外,在席底上发育的一系列原生沉积构造,由于其具有特别的指相意义并有助于古环境重塑而被很自然地归到“第五类原生沉积构造”。该类构造由于其在前寒武纪的普遍性和显生宙的局限分布性,特别是在生物大灭绝事件之后的灾难性泛滥,对于研究地球表层环境的演变具有重要意义。所以说,从席底构造到第五类原生沉积构造,是近年来沉积学发展过程中所出现的较为重要的基本概念。根据这一重要概念,对前寒武纪地层中一些常常被认为是遗迹化石、长期以来存在的“臼齿状构造谜”(Molar-tooth structure enigma)等奇形怪状的沉积构造的成因以及从前寒武纪到显生宙地层中均有分布的特殊的纹理化构造等,将会得到一些更加合理的认识;另一方面,关于微生物形成的原生沉积构造的研究,对于了解地球表层生物变化与沉积变化之间的复杂关系将会发现更多的新问题而得到进一步的思考与研究。</p>
|
| [22] |
Biomats, biofilms, and bioglue as preservational agents for arthropod trackways [J].
Due to divergent taphonomic selection, corresponding body and trace fossils are rarely found in the same rocks. In addition to this general rule, arthropod trackways are preferentially preserved in particular settings: (1) lithographic limestones, where toxic bottom waters account for the exceptional preservation of body fossils at the end of their “mortichnial” trackways; (2) estuarine and lacustrine biolaminites that yield blurred surface tracks as well as the sharper undertracks; and (3) Cambrian intertidal sands before the Precambrian/Cambrian substrate revolution had reached this environment. In all these ichnotopes, the original presence of protective microbial films can be inferred from sedimentary structures. By analogy, it is hypothesised that microbes (“bioglue”) may have been involved in the preservation of trackways in eolian dune sands. The absence of arthropod tracks in Ediacaran sands and silts means either that arthropods had not yet evolved or that they were as yet too tiny to pierce the tougher biomats of the time.
|
| [23] |
Biofilm, biodictyon, biomat microbialites, oolites, stromatolites geophysiology, global mechanism, parahistology [M] |
| [24] |
Microbial biofilms [J]. |
| [25] |
Structure and Function of Biofilms (Dahlem Workshop Reports. Life Sciences Research Report 46) [M]. |
| [26] |
Bacterial polymers: Physicochemical aspects of their interactions at interfaces [J].
How do bacteria stick to a surface? There is still not enough information about to answer this question especially at the molecular level. This question only gives rise to more questions. What is the structure of the true adhesive bacterial polymer? Is only one bacterial polymer or several polymers involved in the adhesion process? What is the role of proteins associated with the bacterial polysaccharides? What type of polymer is produced for the adhesion to hydrophobic surfaces? Is the polymer produced as a response to the surface ?
|
| [27] |
Photosynthesis in cyanobacterial mats and its relation to the sulfur cycle: A model for microbial sulfur interactions [M]∥Cohen Y, Rosenberg E, eds. |
| [28] |
The year of the slime [M]∥KrumbeinW E, Paterson D M, StalL J,ed. |
| [29] |
Bacteriogenic porosity of marine sediments—A case of biomorphogenesis of sedimentary rocks [M]∥Krumbein W E,Paterson D M,Stal L J,eds. |
| [30] |
Extracellular Polymeric Substances (EPS) [M]∥Reitner J, Thiel V,eds. |
| [31] |
The EPS matrix: The "house of biofilm cells" [J]. |
| [32] |
External controls on the distribution, fabrics and mineralization of modern microbial mats in a coastal hypersaline lagoon, Cayo Coco (Cuba) [J].
Active, carbonate-mineralizing microbial mats flourish in a tropical, highly evaporative, marine-fed lagoonal network to the south of Cayo Coco Island (Cuba). Hypersaline conditions support the development of a complex sedimentary microbial ecosystem with diverse morphologies, a variable intensity of mineralization and a potential for preservation. In this study, the role of intrinsic (i.e. microbial) and extrinsic (i.e. physicochemical) controls on microbial mat development, mineralization and preservation was investigated. The network consists of lagoons, forming in the interdune depressions of a Pleistocene aeolian substratum; they developed due to a progressive increase in sea-level since the Holocene. The hydrological budget in the Cayo Coco lagoonal network changes from west to east, increasing the salinity. This change progressively excludes grazers and increases the saturation index of carbonate minerals, favouring the development and mineralization of microbial mats in the easternmost lagoons. Detailed mapping of the easternmost lagoon shows four zones with different flooding regimes. The microbial activity in the mats was recorded using light ark shifts in conjunction with microelectrode O2 and HS profiles. High rates of O2 production and consumption, in addition to substantial amounts of exopolymeric substances, are indicative of a potentially strong intrinsic control on mineralization. Seasonal, climate-driven water fluctuations are key for mat development, mineralization, morphology and distribution. Microbial mats show no mineralization in the permanently submersed zone, and moderate mineralization in zones with alternating immersion and exposure. It is suggested that mineralization is also driven by water-level fluctuations and evaporation. Mineralized mats are laminated and consist of alternating trapping and binding of grains and microbially induced magnesium calcite and dolomite precipitation. The macrofabrics of the mats evolve from early colonizing Flat mats to complex Cerebroid or Terrace structures. The macrofabrics are influenced by the hydrodynamic regime: wind-driven waves inducing relief terraces in windward areas and flat morphologies on the leeward side of the lagoon. Other external drivers include: (i) storm events that either promote (for example, by bioclasts covering) or prevent (for example, by causing erosion) microbial mat preservation; and (ii) subsurface degassing, through mangrove roots and desiccation cracks covered by Flat mats (i.e. forming Hemispheroids and Cerebroidal structures). These findings provide in-depth insights into understanding fossil microbialite morphologies that formed in lagoonal settings.
|
| [33] |
Advances in stromatolite geobiology [J]. |
| [34] |
Production and cycling of natural microbial exopolymers (EPS) within a marine stromatolite [J].
Extracellular polymeric secretions (EPS) that are produced by cyanobacteria represent potential structuring agents in the formation of marine stromatolites. The abundance, production, and degradation of EPS in the upper layers of a microbial mat forming shallow subtidal stromatolites at Highborne Cay, Bahamas, were determined using 14C tracer experiments and were integrated with measurements of other microbial community parameters. The upper regions of a Type 2 [Reid, R.P., Visscher, P.T., Decho, A.W., Stolz, J., Bebout, B., MacIntyre, I.G., Dupraz, C., Pinckney, J., Paerl, H., Prufert-Bebout, L., Steppe, T., Des Marais, D., 2000. The role of microbes in accretion, lamination and early lithification of modern marine stromatolites. Nature (London) 406, 989–992] stromatolite mat exhibited a distinct layering of alternating “green” cyanobacteria-rich layers (Layers 1 and 3) and “white” layers (Layers 2 and 4), and the natural abundance of EPS varied significantly depending on the mat layer. The highest EPS abundance occurred in Layer 2. The production of new EPS, as estimated by the incorporation of 14C-bicarbonate into EPS, occurred in all layers examined, with the highest production in Layer 1 and during periods of photosynthesis (i.e., daylight hours). A large pool (i.e., up to 49%) of the total 14C-bicarbonate uptake was released as low molecular-weight (MW) dissolved organic carbon (DOC). This DOC was rapidly mineralized to CO 2 by heterotrophic bacteria. EPS degradation, as determined by the conversion of 14C-EPS to 14CO 2, was slowest in Layer 2. Results of slurry experiments, examining O 2 uptake following additions of organic substrates, including EPS, supported this degradation trend and further demonstrated selective utilization by heterotrophs of specific monomers, such as acetate, ethanol, and uronic acids. Results indicated that natural EPS may be rapidly transformed post-secretion by heterotrophic degradation, specifically by sulfate-reducing bacteria, to a more-refractory remnant polymer that is relatively slow to accumulate. A mass balance analysis suggested that a layer-specific pattern in EPS and low-MW DOC turnover may contribute to major carbonate precipitation events within stromatolites. Our findings represent the first estimate of EPS turnover in stromatolites and support an emerging idea that stromatolite formation is limited by a delicate balance between evolving microbial activities and environmental factors.
|
| [35] |
Calcified cyanobacteria [M]∥Reitner J, Thiel V,eds. |
| [36] |
|
| [37] |
The evolution of cyanobacterial symbioses [J].
In this paper, the evolution of cyanobacterial symbioses is presented, with emphasis on the molecular phylogeny of the cyanobacteria and their symbiotic associates and the number of independent origins of the symbioses. It also discusses the timing of the origin of cyanobacteria, of organisms symbiotic with cyanobacteria, and of the symbiotic associations, considering the possible evolutionary ...
|
| [38] |
Calcified cyanobacteria [M] |
| [39] |
Composition diversity of modern stromatolites: A key and window for further understanding of the formation of ancient stromatolites [J].现代叠层石的多样化构成:认识古代叠层石形成的关键和窗口 [J].
<p>自从<em>Kalkowsky</em>在<em>1908</em>年构筑了叠层石的术语之后,叠层石一直是地质学家采用不同方法研究和思考的主题,而且一直被当作证明地球早期生命历史的代表物而得到深入调查。叠层石确实为地球早期生命历史提供了间接而且复杂的证据,所以,现代叠层石确实代表着明显的生物信号而成为研究的焦点。最为引人注目的是,现代叠层石的多样化构成,确实表明了蓝细菌生物席建造了叠层石,而且进一步表明了微生物席转化成叠层石不是一个直接的作用过程。那些反映现代叠层石多样化构成的典型实例包括:(<em>1</em>)南极<em>Untersee</em>地区的湖泊相锥状泥质叠层石;(<em>2</em>)新西兰<em>North</em>群岛被称为煎锅湖的热水湖泊中以及美国黄石国家公园热泉中的硅质叠层石;(<em>3</em>)巴哈马台地、澳大利亚鲨鱼湾以及巴西东南部海湾碳酸盐沉积物构成的叠层石。由于蓝细菌微生物席是否代表了古代叠层石的形态学前体总是存在争议,而且在生命的图像中叠层石一直是一个迷惑的关键片段,因此,现代叠层石的多样化构成,将成为认识古代叠层石形成的关键和窗口。立足于前人的研究成果,追踪和总结现代叠层石的多样化构成,以及它们所代表的沉积作用和微生物新陈代谢活动丰富而复杂的信息,将不但丰富微生物沉积学的研究内容,还将拓宽沉积相分析的基本内容,对深入了解叠层石复杂的沉积学特征和生物学属性具有重要的科学意义。</p>
|
| [40] |
The nature of stromatolites: 3,500 million years of history and a century of research [J]. |
| [41] |
Organomineralization derived from the biomineralization: An important theme within the framework of geobiology [J].从生物矿化作用衍生出的有机矿化作用:地球生物学框架下重要的研究主题 [J].
Traditionally, the concept of biomineralization has been defined as the mineral formation by organism and further has been grouped into two types, i.e. the biologically controlled and the biologically induced biomineralizations. This broad term of biomineralization has been emended as the process by which living forms influence the precipitation of mineral materials; correspondingly, the term of biomineral has been redefined as the product of the selective uptake of elements from the local environment, and it can incorporate into functional structures under strict biological control. Correspondingly, the concept of “organomineralization” that is derived from the broad biomineralization is used to describe a mineral formation linked to non living organic substances, and the product resulted from the biomineralization is defined as the “organomineral”, i.e. refers to minerals that are affected by organics, mostly life related, but not directly produced by living cells. Thus, the chief difference between the biomineral and the organomineral is that the later can not incorporate into functional structures under strict biological control. When the biomineralization attract the attention by biologists and chemists, it becomes the research object for the further understanding of “the sophisticate chemical process within life system”; further, the biomineralization gradually becomes an attracting research field of many subjects beyond the geological field. At the same time, studies on the biomineralization result in the advances of the organomineralization. Because the organomineralization and its product, i.e., the organomineral, is volumetrically important constituent of the sedimentary rock record, is synonymous with paleontology and is potentially exciting tracer for life beyond Earth, studies on the organomineralization and its product become an important researching theme within the framework of geobiology.
|
| [42] |
Large sulfur bacteria and the formation of phosphorite [J]. |
| [43] |
A Color Guide to the Petrography of Carbonate Rocks: Grains, Textures, Porosity, Diagenesis, AAPG Memoir 77 [M]. |
| [44] |
Cyanobacterial calcification, carbon dioxide concentrating mechanisms, and Proterozoic-Cambrian changes in atmospheric composition [J].DOI URL [本文引用: 11] 摘要
Photosynthetic uptake of inorganic carbon can raise the pH adjacent to cyanobacterial cells, promoting CaCO 3 precipitation. This effect is enhanced by CO 2 concentrating mechanisms that actively transport into cells for carbon fixation. CO 2 concentrating mechanisms presumably developed in response to atmospheric decrease in CO 2 and increase in O 2 over geological timescales. In present-day cyanobacteria, CO 2 concentrating mechanisms are induced when the atmospheric partial pressure of CO 2 (p CO2 ) falls below 0908040.4%. Reduction in p CO2 during the Proterozoic may have had two successive effects on cyanobacterial calcification. First, fall in p CO2 below 0908041% (33 times present atmospheric level, PAL) resulted in lower dissolved inorganic carbon (DIC) concentrations that reduced pH buffering sufficiently for isolated CaCO 3 crystals to begin to nucleate adjacent to cyanobacterial cells. As a result, blooms of planktic cyanobacteria induced precipitated 'whitings' of carbonate mud in the water column whose sedimentary accumulation began to dominate carbonate platforms 09080414000900091300 Ma. Second, fall in p CO2 below 0908040.4% (10 PAL) induced CO 2 -concentrating mechanisms that further increased pH rise adjacent to cells and promoted in vivo cyanobacterial sheath calcification. Crossing of this second threshold is indicated in the fossil record by the appearance of Girvanella 750090009700 Ma. Coeval acquisition of CO 2 concentrating mechanisms by planktic cyanobacteria further stimulated whiting production. These inferences, that p CO2 fell below 0908041%09080414000900091300 Ma and below 0908040.4% 750090009700 Ma, are consistent with empirical and modelled palaeo-atmosphere estimates. Development of CO 2 concentrating mechanisms was probably temporarily slowed by global cooling 090804700090009570 Ma that favoured diffusive entry of CO 2 into cells. Lower levels of temperature and DIC at this time would have reduced seawater carbonate saturation state, also hindering cyanobacterial calcification. It is suggested that as Earth emerged from 'Snowball' glaciations in the late Neoproterozoic, global warming and O 2 rise reactivated the development of CO 2 concentrating mechanisms. At the same time, rising levels of temperature, calcium ions and DIC increased seawater carbonate saturation state, stimulating widespread cyanobacterial in vivo sheath calcification in the Early Cambrian. This biocalcification event promoted rapid widespread development of calcified cyanobacterial reefs and transformed benthic microbial carbonate fabrics.
|
| [45] |
An early-branching microbialite cyanobacterium forms intracellular carbonates [J].
Cyanobacteria have affected major geochemical cycles (carbon, nitrogen, and oxygen) on Earth for billions of years. In particular, they have played a major role in the formation of calcium carbonates (i.e., calcification), which has been considered to be an extracellular process. We identified a cyanobacterium in modern microbialites in Lake Alchichica (Mexico) that forms intracellular amorphous calcium-magnesium-strontium-barium carbonate inclusions about 270 nanometers in average diameter, revealing an unexplored pathway for calcification. Phylogenetic analyses place this cyanobacterium within the deeply divergent order Gloeobacterales. The chemical composition and structure of the intracellular precipitates suggest some level of cellular control on the biomineralization process. This discovery expands the diversity of organisms capable of forming amorphous calcium carbonates.
|
| [46] |
Evidence for $HCO_{3}^{-}$ transport by the Blue-Green Alga (Cyanobacterium) Coccochloris peniocystis [J]. |
| [47] |
Mesoproterozoic carbon dioxide levels inferred from calcified cyanobacteria [J].
Filamentous and shrub-like carbonate fabrics produced by in vivo cyanobacterial sheath calcification in stromatolites of the ca. 1200 Ma Society Cliffs Formation, Baffin and Bylot Islands, Arctic Canada, are 400 m.y. older than previously reported examples. In vivo sheath calcification is promoted by carbon dioxide concentrating mechanisms (CCMs) and is a direct ecophysiological link to atmospheric COconcentration. CCMs are induced in present-day cyanobacteria under experimental conditions when pCOis below 0.36% ( 10 times present atmospheric level, PAL). Society Cliffs calcified cyanobacteria consequently imply pCOlevels of <0.36% at ca. 1200 Ma. This inference is consistent with marine carbon isotope modeling that suggests pCOof 7-10 PAL in the late Mesoproterozoic. Combined, petro-graphic, experimental, and modeling results therefore suggest that Mesoproterozoic pCOconcentrations were not substantially different from Phanerozoic values and were significantly less than previous estimates of up to 200 PAL. Assuming 10% lower solar luminosity in the late Mesoproterozoic, pCOlevels of 10 PAL or less require the presence of additional greenhouse gases for maintenance of an ice-free Earth. At 10 PAL pCO, methane concentrations of 100-200 ppm would have been sufficient to sustain warm Earth surface conditions. The low atmospheric oxygen and limited marine sulfate concentrations required to sustain atmospheric methane provide additional support for sulfur isotope models that suggest protracted oxygenation of Earth's Proterozoic biosphere.
|
| [48] |
Cyanobacterial precipitation of gypsum, calcite, and magnesite from natural alkaline lake water [J]. |
| [49] |
Microbial carbonates: A review and perspectives [J].微生物碳酸盐岩研究:回顾与展望 [J].
在查阅国内外大量文献资料的基础上,回顾与综述了微生物碳酸盐岩近几年的研究现状及进展。微生物碳酸盐岩是由钙化微生物(蓝绿菌、微藻等)在微生物作用、沉积作用及成岩作用的共同作用下形成的,其种类繁多,包括叠层石、凝块石、树形石、均一石、核形石和纹理石等。它在各个地质历史时期均有发育,但主要繁盛于中新元古代和早古生代;自显生宙以来,随着后生动物的繁育微生物碳酸盐岩总体呈衰退趋势。另外,微生物对白云石的直接沉淀、碳酸盐泥丘及第五类原生沉积构造的形成具有重要作用。近年来,对微生物碳酸盐岩研究的发展使得碳酸盐岩的成因及分类得到了补充与完善。
|
| [50] |
Bio-ore formation and biomineralization [J].生物成矿作用与生物矿化作用 [J].
About fifty--five kinds of biominerals are listed and divided into four stages of amorphous,intermediate, matured and fossilized factes. The biominerals are mainly character lied by weak alkali and weak acid 'salts,calcareous compounds (25 kinds),oxides (12) andoxysalts (32), hydrated (25) or hydroxylated (6) minerals and uniaxial negative crystalsor approximately the like.
|
| [51] |
|
| [52] |
Non-biologically supported organomineralization [J]. |
| [53] |
Defining biominerals and organominerals: Direct and indirect indicators of life [J].
By introducing the new term ‘organomineral’ to apply to minerals that are affected by organics, mostly life-related, but not directly produced by living cells, we hope to increase the accuracy of the terminology in present usage. We believe that the term ‘biomineral’ does not describe all mineral deposits precisely enough and offer case studies to support instances where the use of the new term ‘organomineral’ is more appropriate. We provide examples of some materials that are biominerals such as those in diatoms and human bones and which perform a functional purpose. We then attempt to show that terminology can sometimes mislead investigators, drive the direction and prejudice interpretations of scientific investigation. This is achieved by presenting case studies of minerals that have been investigated as biominerals although they may not actually be directly controlled by biology. For this reason we pay special attention to desert varnish, hot-spring siliceous deposits, stromatolites, and bacterial encrustations. We go on to explain why we regard biominerals as direct indictors of life whereas organominerals should be treated as indirect indicators of life that require careful characterization to establish the origin of contained organics and whether these are biotic in origin. How organic compounds are preserved and transformed in sedimentary environments and in the mineral matrix is as highly relevant to the study of modern Earth environments as it is in the search for the oldest evidence of life in the terrestrial rock record. It is also important to many disciplines such as microbiology and archaeology and especially to scientists planning missions to look for evidence of ‘life’ on other planetary surfaces.
|
| [54] |
Function, structure and composition of cell walls and external layers [M]∥Carr N G, Whittnm B A, eds. |
| [55] |
The Genera Beggiatoa,Thioploca [M] |
| [56] |
Microbial mats as bioreactors: Populations, processes, and products [J].
Microbial mats are dynamic and complex ecosystems exhibiting spatial and temporal heterogeneity. The physical/chemical environment is typified by steep gradients and distinct microenvironments. These microenvironments support a great diversity of species with a wide range of metabolic processes. These processes often result in coupled reactions and biogeochemical cycles, and produce important end products such as trace gases and mineral precipitates. The latter can impact the composition and character of the sediment, imparting a iosignature. These biosignatures can be preserved in the rock record and are useful in the interpretation of fossil record on Earth and possibly as an indication of life on other planetary bodies. The modern marine stromatolites of the Exuma Cays, Bahamas, provide an ideal system for studying the populations, processes, and products in a microbial ecosystem using a multidisciplinary approach. In order to acquire redox energy, microbial populations need to carry out metabolic reactions at rates faster than the equivalent chemical (abiotic) reactions. As such, microbes can be viewed as bioreactors that preferably oxidize carbon to CO 2 to maximize the energy yield. The study of the microbial role in carbonate sedimentation and lithification in these stromatolites provides a picture of microbial mats as bioreactors producing a biosignature.
|
| [57] |
Microelectrode measurements in stromatolites: Unraveling the Earth's past? [J]. |
| [58] |
Biogeochemistry: Space for hydrogen [J]. |
| [59] |
Photosynthesis-induced biofilm calcification and calcium concentrations in Phanerozoic Oceans [J].
Photosynthetic carbon assimilation is commonly invoked as the cause of calcium carbonate precipitation in cyanobacterial biofilms that results in the formation of calcareous stromatolites. However, biofilm calcification patterns in recent lakes and simulation of photosynthetically induced rise in calcium carbonate supersaturation demonstrate that this mechanism applies only in settings low in dissolved inorganic carbon and high in calcium. Taking into account paleo-partial pressure curves for carbon dioxide, we show that Phanerozoic oceans sustaining calcified cyanobacteria must have had considerably higher calcium concentrations than oceans of today. In turn, the enigmatic lack of calcified cyanobacteria in stromatolite-bearing Precambrian sequences can now be explained as a result of high dissolved inorganic carbon concentrations.
|
| [60] |
Exopolymeric substances of sulfate-reducing bacteria: Interactions with calcium at alkaline pH and implication for formation of carbonate minerals [J].
ABSTRACT Sulfate-reducing bacteria (SRB) have been recognized as key players in the precipitation of calcium carbonate in lithifying microbial communities. These bacteria increase the alkalinity by reducing sulfate ions, and consuming organic acids. SRB also produce copious amounts of exopolymeric substances (EPS). All of these processes influence the morphology and mineralogy of the carbonate minerals. Interactions of EPS with metals, calcium in particular, are believed to be the main processes through which the extracellular matrix controls the precipitation of the carbonate minerals. SRB exopolymers were purified from lithifying mat and type cultures, and their potential role in CaCO 3 precipitation was determined from acid-base titrations and calcium-binding experiments. Major EPS characteristics were established using infrared spectroscopy and gas chromatography to characterize the chemical functional groups and the sugar monomers composition. Our results demonstrate that all of the three SRB strains tested were able to produce large amounts of EPS. This EPS exhibited three main buffering capacities, which correspond to carboxylic acids (pK a = 3.0), sulfur-containing groups (thiols, sulfonic and sulfinic acids - pK a = 7.0-7.1) and amino groups (pK a = 8.4-9.2). The calcium-binding capacity of these exopolymers in solution at pH 9.0 ranged from 0.12g Ca g EPS -1 -0.15 g Ca g EPS -1 . These results suggest that SRB could play a critical role in the formation of CaCO 3 in lithifying microbial mats. The unusually high sulfur content, which has not been reported for EPS before, indicates a possible strong interaction with iron. In addition to changing the saturation index through metabolic activity, our results imply that SRB affect the rock record through EPS production and its effect on the CaCO 3 precipitation. Furthermore, EPS produced by SRB may account for the incorporation of metals (e.g. Sr, Fe, Mg) associated with carbonate minerals in the rock record.
|
| [61] |
A revised classification of limestones [J].
The most widely used classifications of limestones are now thirty years old and our appreciation of the diagenetic effects on limestone textures is now much greater. A revision of the classifications of Dunham (1962) and Embry and Klovan (1971) is offered and new "diagenetic" categories are proposed. The confusing term lime mudstone is replaced by calcimudstone. Two categories of diagenetic textures are recognized: non-obliterative and obliterative. The former contains limestones which are almost wholly composed of diagenetic cement (cementstones) and those whose fabric is the result of intergranular pressure solution (condensed and fitted grainstones). Obliterative categories include limestones and dolomites whose original fabric has been destroyed; sub-categories include sparstones (or dolosparstones) (crystals > 10 m in size) and microsparstones (dolomicrosparstones) (crystals 4-10 m in size) and dolomicrostones crystals (< 4 m in size). Many diagenetic processes can produce fabrics which mimic depositional ones and these are reviewed.
|
| [62] |
Spectral subdivision of limestone type [M]∥Ham W E,ed. |
| [63] |
Classification of carbonates rocks according to Deposicional texture [J]. |
| [64] |
A late Devonian reef tract on northeastern Banks Island, N.W.T [J]. |
| [65] |
A comment on the systematic confusion of thrombolites [J]. |
| [66] |
Formative mechanisms, depositional processes, and geological implications of Furongian (late Cambrian) reefs in the North China Platform [J].
61Describe various microbial–metazoan and microbial reefs from the Furongian61Provide a model of reef development in response to sea-level changes61Reveal an abrupt transition in reef components from Cambrian Series 3 to Furongian
|
| [67] |
The middle-late Cambrian reef transition and related geological events: A review and new view [J].
The reefs of the middle ate Cambrian (Cambrian Epoch 3 urongian) have long been simply considered as microbialites that flourished in the aftermath of the archaeocyath extinction. Thorough review of the reefs in these periods shows, however, that the Cambrian Epoch 3 and the Furongian actually yield different types of reefs. The Cambrian Epoch 3 reefs are dominated by thrombolites and dendrolites, largely constructed by the calcified microbes Epiphyton and Renalcis. On the other hand, the Furongian reefs consist mainly of maze-like maceriate reefs and columnar stromatolites. The maceriate reefs most likely formed by siliceous sponges and calcified microbes including Girvanella and Tarthinia, whereas the columnar stromatolites were mainly constructed by Girvanella, Tarthinia, and minor siliceous sponges. Other microbial reefs (e.g., non-columnar stromatolites) persisted during the Cambrian Epoch 3 and the Furongian, mainly as small patch reefs or reefal crusts. Lithistid sponge-microbial reefs initially formed in the Cambrian Epoch 3 and occurred throughout the Furongian, but occupied only a minor portion during these periods. Several geological events occurred across the boundary between the Cambrian Epoch 3 and the Furongian, including positive carbon and sulfur isotope excursions, a eustatic sea-level drop, major faunal turnover of trilobites, and diversification of new organisms. The coincidence of these events and the transition in reefal fabrics suggest that reef ecosystems were significantly influenced by at least some of these events. Among the events, it is likely that sea-level change would have affected reefs, although further studies are required in order to test whether and how the other events influenced the reef transition. Epiphyton and Renalcis (and thrombolites and dendrolites formed by them) declined greatly most likely due to sea-level fall and thus decreased in shallow marine habitat at the end of Cambrian Epoch 3. The Cambrian Epoch 3 thrombolites and dendrolites were gradually replaced by a sponge-microbial association of the maze-like, maceriate reefs and the columnar stromatolites in the Furongian, coincident with a sea-level rise.
|
| [68] |
Carbonate factories: A conundrum in sedimentary geology [J].
Describing, characterizing and interpreting the nearly infinite variety of carbonate rocks are conundrums – intricate and difficult problems having only conjectural answers – that have occupied geologists for more than two centuries. Depositional features including components, rock textures, lithofacies, platform types and architecture, all vary in space and time, as do the results of diagenetic processes on those primary features. Approaches to the study of carbonate rocks have become progressively more analytical. One focus has evolved from efforts to build reference models for specific Phanerozoic windows to scrutinize the effect of climate and long-term oscillations of the ocean–atmosphere system in influencing the mineralogy of carbonate components. This paper adds to the ongoing lively debates by attempting to understand changes in the predominant types of carbonate-producing organisms during the Mesozoic–Cenozoic, while striving to minimize the uniformitarian bias. Our approach integrates estimates of changes in Ca 2+ concentration in seawater and atmospheric CO 2, with biological evolution and ecological requirements of characteristic carbonate-producing marine communities. The underlying rationale for our approach is the fact that CO 2 is basic to both carbonates and organic matter, and that photosynthesis is a fundamental biological process responsible for both primary production of organic matter and providing chemical environments that promote calcification. Gross photosynthesis and hypercalcification are dependent largely upon sunlight, while net primary production and, e.g., subsequent burial of organic matter typically requires sources of new nutrients (N, P and trace elements). Our approach plausibly explains the changing character of carbonate production as an evolving response to changing environmental conditions driven by the geotectonic cycle, while identifying uncertainties that deserve further research. With metazoan consumer diversity reduced by the end-Permian extinctions, excess photosynthesis by phytoplankton and microbial assemblages in surface waters, induced by moderately high CO 2 and temperature during the Early Mesozoic, supported proliferation of non-tissular metazoans (e.g., sponges) and heterotrophic bacteria at the sea floor. Metabolic activity by those microbes, especially sulfate reduction, resulted in abundant biologically-induced geochemical carbonate precipitation on and within the sea floor. For example, with the opening of Tethyan seaways during the Triassic, massive sponge/microbe boundstones (the benthic automicrite factory) formed steep, massive and thick progradational slopes and, locally, mud-mounds. As tectonic processes created shallow epicontinental seas, photosynthesis drove lime-mud precipitation in the illuminated zone of the water column. The resulting neritic lime-mud component of the shallow-water carbonate factory became predominant during the Jurassic, paralleling the increase in atmospheric pCO 2, while the decreasing importance of the benthic automicrite factory parallels the diversification of calcifying metazoans, phytoplankton and zooplankton. With atmospheric pCO 2 declining through the Cretaceous, the potential habitats for neritic lime-mud precipitation declined. At the same time, peak oceanic Ca 2+ concentrations promoted biotically-controlled calcification by the skeletal factory. With changes produced by extinctions and turnovers at the Cretaceous–Tertiary boundary, adaptations to decreasing Ca 2+ and pCO 2, coupled with increasing global temperature gradients (i.e., high-latitude and deep-water cooling), and strategies that efficiently linked photosynthesis and calcification, promoted successive changes of the dominant skeletal factory through the Cenozoic: larger benthic foraminifers (protist–protist symbiosis) during the Paleogene, red algae during the Miocene and modern coral reefs (metazoan–protist symbiosis) since Late Miocene.
|
| [69] |
Stromatolites [M]. |
| [70] |
Stromatolite morphogenesis: Progress and problems [J]. |
| [71] |
Respect for stromatolites [J]. |
| [72] |
Classification andenvironmental significance of cryptalgal Limestones and dolomites, with illustrations from the cambrian and ordovician of southwestern Alberta [J]. |
| [73] |
|
| [74] |
Characteristic and Geological Significance of Dendrolite from Zhangxia Formation,Cambrian, Jiulongshan Section [D].莱芜九龙山寒武系张夏组树形石特征及地质意义 [D]. |
| [75] |
Remarks on the systematic position of Lithocodium, Elliott, a problematic microorganism from the Mesozoic carbonate platforms of the Tethyan realm [J].
The encrusting genus Lithocodium Elliott, widespread in Tethyan Triassic-Cretaceous carbonate platforms and interpreted in the past as a codiacean alga, is presently regarded as a colony of calcified cyanobacteria, a microorganism incertae sedis or a loftusiacean foraminifer. The absence of apertures connecting neighbouring cavities, as well as the very irregular form and arrangement of these hollows within a frequently massive micritic matrix, excludes assigning Lithocodium to the foraminifers, but could rather suggest colonies of calcified cyanobacteria.
|
| [76] |
The jurassic Lithocodium aggregatum-Troglotella incrustans foraminiferal consortium [J]. |
| [77] |
Calcified cyanobacterias within the stromatolotic bioherm for the Cambrian Furongian Series in the northeastern margin of the North-China Platform [J].华北地台东北缘寒武系芙蓉统叠层石生物丘中的钙化蓝细菌 [J].
华北地台东北缘的芙蓉统,大致为长山组和凤山组所组成,可以进一步划分为3个三级沉积层序;层序划分主要基于沉积相序列的旋回性所代表的沉积趋势,较深水的陆棚相钙质泥岩和深缓坡相条带状泥晶灰岩和泥灰岩组成的凝缩作用序列、与高水位体系域和强迫型海退体系域的中至浅缓坡相碳酸盐岩组成的总体向上变浅序列,是这些三级沉积层序的基本构成,从而形成了较为典型的淹没不整合型层序。那些典型的叠层石生物丘,类似于微生物礁,主要发育在长山组和凤山组下部构成的三级层序的强迫型海退体系域之中,代表了缓坡型台地中相对海平面下降阶段的沉积记录。这些叠层石生物丘中的叠层石,泥晶和微亮晶是其基本构成,最为特征的是发育着一些典型的钙化蓝细菌化石,表明了这些寒武纪芙蓉世的叠层石生长于蓝细菌主导的微生物席的钙化作用之中。最为重要的是,在构成叠层石生物丘的粗糙纹层柱状和穹窿状叠层石中,较为普遍地发育着"石松藻(Lithocodium)";这种谜一样的钙化蓝细菌化石,与其他的钙化蓝细菌化石一起,表明了寒武纪叠层石形成过程中复杂的微生物沉淀作用,成为窥视叠层石生长和石化过程中重要的微生物作用信号。就像其名称所蕴含着的高级绿藻中的松藻(Codium)的涵义一样,"石松藻(Lithocodium)"状的钙化蓝细菌,多描述于中生代的微生物碳酸盐岩中,而且还常常被解释为结壳状有孔虫或"海绵骨针的网状物",其生物亲和性还存在着剧烈的争论。因此,华北地台东北缘寒武系芙蓉统中的叠层石生物丘,特殊的层序地层位置代表了较为典型的强迫型海退沉积记录,特别的钙化微生物构成代表着叠层石生长和石化过程中复杂的微生物作用信号,成为深入了解"寒武纪-早奥陶世微生物碳酸盐岩复苏期"和"显生宙早期第一幕蓝细菌钙化作用事件"中的微生物造礁和成丘作用的典型实例。
|
| [78] |
Proterozoic stromatolite zonation [J]. |
| [79] |
|
| [80] |
Sulfate reducing bacteria in microbial mats: Changing paradigms, new discoveries [J].
Sulfate reducing bacteria (SRB) have existed throughout much of Earth's history and remain major contributors to carbon cycling in modern systems. Despite their importance, misconceptions about SRB are prevalent. In particular, SRB are commonly thought to lack oxygen tolerance and to exist only in anoxic environments. Through the last two decades, researchers have discovered that SRB can, in fact, tolerate and even respire oxygen. Investigations of microbial mat systems have demonstrated that SRB are both abundant and active in the oxic zones of mats. Additionally, SRB have been found to be highly active in the lithified zones of microbial mats, suggesting a connection between sulfate reduction and mat lithification. In the present paper, we review recent research on SRB distribution and present new preliminary findings on both the diversity and distribution of -proteobacterial SRB in lithifying and non-lithifying microbial mat systems. These preliminary findings indicate the unexplored diversity of SRB in a microbial mat system and demonstrate the close microspatial association of SRB and cyanobacteria in the oxic zone of the mat. Possible mechanisms and further studies to elucidate mechanisms for carbonate precipitation via sulfate reduction are also discussed.
|
| [81] |
Bacterial roles in the precipitation of carbonate minerals [M] |
| [82] |
Microscale observations of sulfate reduction: Correlation of microbial activity with lithified micritic laminae in modern marine stromatolites [J].
We report for the first time micrometer-scale correlation of geologic and microbial processes in modern marine stromatolites. Precipitation of micritic laminae in these stromatolites was studied by comparing microstructure, as observed in petrographic thin sections, with microbial sulfate-reduction activity. Two-dimensional mapping of sulfate-reduction rates was implemented by incubating a vertical section of a stromatolite face on silver foil coated with SO. Our results show that sulfate-reduction activity is high in zones of CaCOprecipitation and indicate that microbial activity produces lithified micritic laminae near the surface of the stromatolites. Similarities with micritic laminae in ancient stromatolites suggest that sulfate reduction may also have been an important mechanism of carbonate precipitation in these fossilized structures.
|
| [83] |
Seasonal records of climatic change in annually laminated tufas: Short review and future prospects [J].
Many Recent and fossil freshwater tufa stromatolites contain millimetre-scale, alternating laminae of dense micrite and more porous or sparry crystalline calcites. These alternating laminae have been interpreted to represent seasonally controlled differences in the biotic activity of microbes, and/or seasonally controlled changes in the rate of calcification. Either way, couplets of these microbially mediated alternating calcified laminae are generally agreed to represent annual seasonality. Combined stable isotope (18O and 13C) and trace element (Mg, Sr, Ba) geochemistry from Recent tufa stromatolites show that seasonal climatic information is available from these calcites. Variability in 18O (and in one case Mg concentration) has been shown to be controlled primarily by stream temperature change, usually driven by solar insolation. In arid climates, seasonal evaporation can also cause 18O enrichment by at least 1. Variability in 13C results potentially from: (1) seasonal change in plant uptake of 12C-enriched CO2; (2) seasonal change in degassing of 12C-enriched CO2 in the aquifer system; and (3) precipitation of calcite along the aquifer or river flow path, a process that increases 13C of dissolved inorganic carbon (DIC) in the remaining water. Mechanisms 2 and 3 are linked because calcite precipitates in aquifers where degassing occurs, e.g. air pockets. The latter mechanism for 13C enrichment has also been shown to cause sympathetic variation between trace element/Ca ratios and 13C because trace elements with partition coefficients much greater than 1 (e.g. Sr, Ba) remain preferentially in solution. Since degassing in air pockets will be enhanced during decreased recharge when water saturation of the aquifer is lowest, sympathetic variation in trace element/Ca ratios and 13C is a possible index of recharge and therefore precipitation intensity. High-resolution geochemical data from well-dated tufa stromatolites have great potential for Quaternary palaeoclimate reconstructions, possibly allowing recovery of annual seasonal climatic information including water temperature variation and change in rainfall intensity. However, careful consideration of diagenetic effects, particularly aggrading neomorphism, needs to be the next step. Copyright 2005 John Wiley & Sons, Ltd.
|
| [84] |
Evidence of microbiocoenosis in the formation of laminae in modern stromatolites [J].
The first appearance of laminated stromatolites in sedimentary rocks around 3.5 billion years ago is generally accepted as evidence for the earliest advanced ecosystem. However, the bio-mechanisms leading to the construction of stromatolites continue to be poorly understood. Using a combination of microscopic and advanced spectroscopic techniques (STXM/NEXAFS/EELS) to analyze lithified laminae, living biomass, reference microorganisms and their byproducts, it is demonstrated that lithified laminae contain the bio-molecular signatures of sulphide oxidizers and sulphate reducers, as well as cyanobacterial extracellular polymeric substances. From this, it is concluded that a combination of microbial processes, such as the degradation of cyanobacterial biomass, phototrophic sulphide oxidation, and sulphate reduction, results in the formation of laminae, a process here called amalgamation. In summary, the described microbialite formation process is biochemically controlled with minimal external particle input and serves as an alternative biotic explanation for the trapping and binding model of lamina formation.
|
| [85] |
The microbial communities of the modern marine stromatolites at Highborn Cay, Bahamas [J]. |
| [86] |
Discovery of large conical stromatolites in Lake Untersee, Antarctica [J].
Lake Untersee is one of the largest (11.4 km2) and deepest (>160 m) freshwater lakes in East Antarctica. Located at 7100°S the lake has a perennial ice cover, a water column that, with the exception of a small anoxic basin in the southwest of the lake, is well mixed, supersaturated with dissolved oxygen, alkaline (pH 10.4) and exceedingly clear. The floor of the lake is covered with photosynthetic microbial mats to depths of at least 100 m. These mats are primarily composed of filamentous cyanophytes and form two distinct macroscopic structures, one of which 090009 cm-scale cuspate pinnacles dominated by Leptolyngbya spp. 090009 is common in Antarctica, but the second 090009 laminated, conical stromatolites that rise up to 0.5 m above the lake floor, dominated by Phormidium spp. 090009 has not previously been reported in any modern environment. The laminae that form the conical stromatolites are 0.20900090.8 mm in thickness consisting of fine clays and organic material; carbon dating implies that laminations may occur on near decadal timescales. The uniformly steep sides (59.6 00± 2.500°) and the regular laminar structure of the cones suggest that they may provide a modern analog for growth of some of the oldest well-described Archean stromatolites. Mechanisms underlying the formation of these stromatolites are as yet unclear, but their growth is distinct from that of the cuspate pinnacles. The sympatric occurrence of pinnacles and cones related to microbial communities with distinct cyanobacterial compositions suggest that specific microbial behaviors underpin the morphological differences in the structures.
|
| [87] |
Brief introduction on new advances on the origin of oids [J].鲕粒成因研究的新进展 [J].
鲕粒的成因一直是一个谜一样的沉积学难题。Brehm等在2006年的实验室研究的结果表明,将鲕粒的形成可以与叠层石进行类比,是一个特殊的球状微生物席的产物,从而将鲕粒归为微生物成因。最近,来自于巴哈马现代鲕粒的研究,Duguid等在2010年认为,鲕粒形成与微生物活动不存在一个直接的关系,重新强调了鲕粒形成的化学过程,即:当鲕粒处在沉积物—水界面(活跃相)的时候,凭借一个在鲕粒上的非晶质钙碳酸盐(ACC)沉淀物的微薄层,鲕粒开始形成;这个ACC微薄层后来结晶成文石针,从而形成新的鲕粒皮层(静止相)。两种观点的直接对立,代表了对鲕粒成因研究的新进展。来自于华南下三叠统的巨鲕、以及华北寒武系鲕粒,特殊的显微组构似乎更多地支持了鲕粒形成的微生物作用机制的学说。
|
| [88] |
Hot spring siliceous stromatolites from Yellowstone National Park: Assessing growth rate and laminae formation [J].
Stromatolites are commonly interpreted as evidence of ancient microbial life, yet stromatolite morphogenesis is poorly understood. We apply radiometric tracer and dating techniques, molecular analyses and growth experiments to investigate siliceous stromatolite morphogenesis in Obsidian Pool Prime (OPP), a hot spring in Yellowstone National Park. We examine rates of stromatolite growth and the environmental and/or biologic conditions that affect lamination formation and preservation, both difficult features to constrain in ancient examples. The 090008main body090009 of the stromatolite is composed of finely laminated, porous, light090009dark couplets of erect (surface normal) and reclining (surface parallel) silicified filamentous bacteria, interrupted by a less-distinct, well-cemented 090008drape090009 lamination. Results from dating studies indicate a growth rate of 10900095 cm year0908081; however, growth is punctuated. 14C as a tracer demonstrates that stromatolite cyanobacterial communities fix CO2 derived from two sources, vent water (radiocarbon dead) and the atmosphere (modern 14C). The drape facies contained a greater proportion of atmospheric CO2 and more robust silica cementation (vs. the main body facies), which we interpret as formation when spring level was lower. Systematic changes in lamination style are likely related to environmental forcing and larger scale features (tectonic, climatic). Although the OPP stromatolites are composed of silica and most ancient forms are carbonate, their fine lamination texture requires early lithification. Without early lithification, whether silica or carbonate, it is unlikely that a finely laminated structure representing an ancient microbial mat would be preserved. In OPP, lithification on the nearly diurnal time scale is likely related to temperature control on silica solubility.
|
| [89] |
Brief history and current status of stromatolite study in China [J].中国叠层石研究的历史和现状 [J]. |
| [90] |
Taking advantage of disorder: Amorphous calcium carbonate and its roles in biomineralization [J].
Amorphous calcium carbonate (ACC) in its pure form is highly unstable, yet some organisms produce stable ACC, and cases are known in which ACC functions as a transient precursor of more stable crystalline aragonite or calcite. Studies of biogenic ACC show that there are significant structural differences, including the observation that the stable forms are hydrated whereas the transient forms are not. The many different ways in which ACC can be formed in vitro shed light on the possible mechanisms involved in stabilization, destabilization, and transformation of ACC into crystalline forms of calcium carbonate. We show here that ACC is a fascinating form of calcium carbonate that may well be of much interest to materials science and biomineralization.
|
| [91] |
|
| [92] |
The "Carbonate Factory" revisited: A reexamination of sediment production functions used to model deposition on carbonate platforms [J].
There are two widely used models of sediment production on carbonate shelves: (1) production varying with depth, and (2) production as a function of depth and distance from the platform margin. Here we recalculate carbonate production on Great Bahama Bank west and northwest of Andros Island using Broecker and Takahashi's (1966) pioneering geochemical study. In the calculations, residence time and depth are used to calculate yearly carbonate production based on equilibrium thermodynamics of CaCO 3 loss from water over the banks. Whereas Broecker and Takahashi assumed a uniform depth of 4.5 m in their work, here we use realistic depths and residence times interpolated over the bank. Our results suggest that sediment production is on the order of 4-5 kg/m 2/y on the bank margins and that production rapidly drops off into the platform, approaching zero in the shallowest, innermost parts of the platform. Thus, for a large platform such as Great Bahama Bank with long residence times, the shallowest areas of the platform may become sinks, not sources, of sediment.
|
| [93] |
The carbonate factory of Middle Triassic buildups in the Dolomites, Italy: A quantitative analysis [J].
Middle Triassic carbonate buildups of the Dolomites were high in relief (500 1000m) and small in size (one to a few square kilometres in area). A paradox results from the carbonate platform model that invokes the platform top, including reef rims, as the carbonate factory and flanking beds as talus deposits. Most buildups consist largely of clinoforms (inclined at 10-50 ) whereas massive reef rocks and stratified buildup interiors are poorly developed or absent. Facies and modal analysis of 323 thin sections from buildups of the Marmolada indicate that clinoforms are: (i) predominantly composed of in situ boundstones (56% of all samples); (ii) primarily made up of early cements (37 vol.%), microbial crusts (17 vol.%), micritic intraclasts (10 vol.%) and Tubiphytes (8 vol.%); and (iii) contain diagnostic shallow water grains (dasyclads, coated grains) that are less abundant by 1-2 orders of magnitude compared with buildup interior facies. These data suggest that the clinoforms themselves were the main carbonate factory of the Triassic buildups. Stratified buildup interior rocks and massive reef rocks were apparently not a prerequisite for buildup growth and clinoform progradation.
|
| [94] |
The macroalgal carbonate factory at a cool-to-warm temperate marine transition, Southern Australia [J]. |
| [95] |
Benthic carbonate factories of the Phanerozoic [J].
Marine carbonate precipitation occurs in three basic modes: abiotic (or quasi-abiotic), biotically induced, and biotically controlled. On a geologic scale, these precipitation modes combine to form three carbonate production systems, or "factories" in the benthic environment: (1) tropical shallow-water factory, dominated by biotically controlled (mainly photo-autotrophic) and abiotic precipitates; (2) cool-water factory, dominated by biotically controlled (mainly heterotrophic) precipitates; and (3) mud-mound factory, dominated by biotically induced (mainly microbial) and abiotic precipitates. Sediment accumulations of the factories differ in composition, geometry, and facies patterns, and some of these differences appear prominently in seismic data, thus facilitating subsurface prediction. The characteristic accumulation of the tropical factory is the flat-topped, often reef-rimmed platform. In cool-water systems, reefs in high-energy settings are scarce and hydrodynamic influence dominates, producing seaward-sloping shelves and deep-water sediment drifts often armored by skeletal framework. The typical accumulation of the mud-mound factory is groups of mounds in deeper water. Where the mud-mound factory expands into shallow water, it forms rimmed platforms similar to the tropical factory. The tropical factory is most productive; the mud-mound factory reaches 80 90%, and the cool-water factory 20 30% of the tropical growth rate. The three factories represent end members connected by transitions in space. Transitions in time are linked to biotic evolution.
|
| [96] |
Geology of the Shark Bay area, Western Australia [R]∥Berry P F, Bradshaw S D, Wilson B R, eds. |

/
| 〈 |
|
〉 |

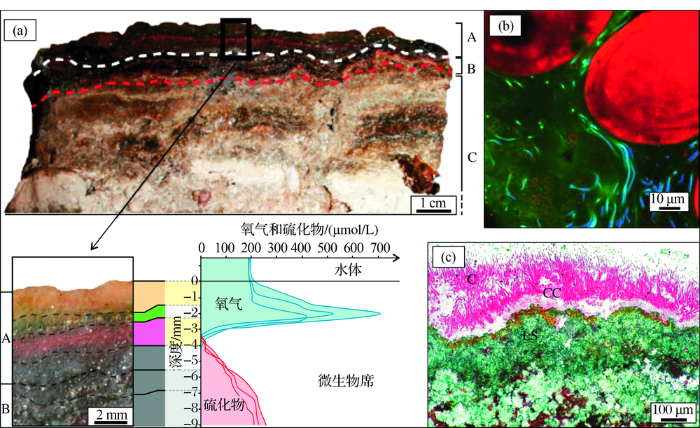

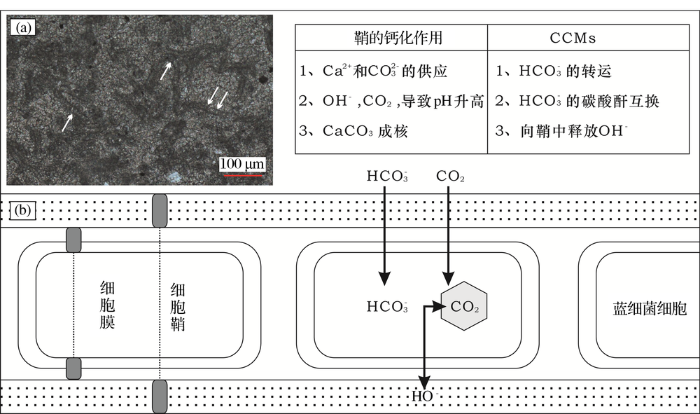
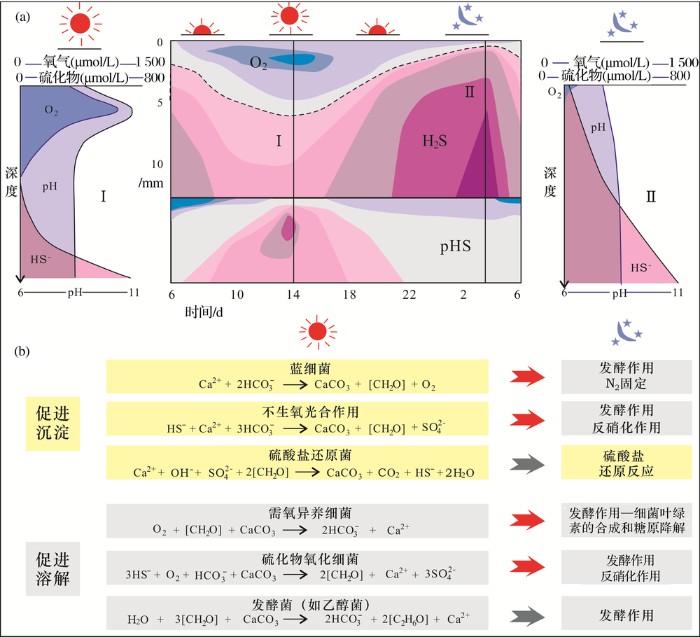
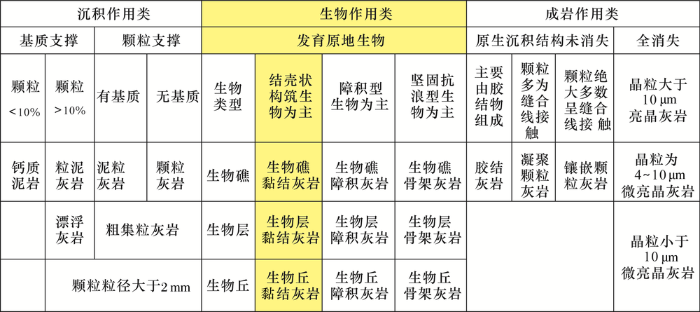



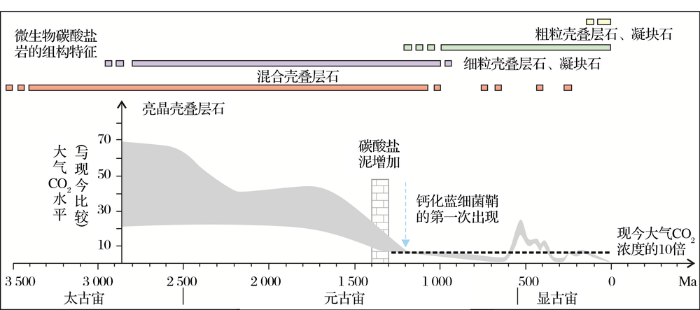
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}