First author:Weng Chengyu(1965-),male, Kunming City, Yunnan Province, Professor. Research areas include quaternary geology, palaeoecology and palaeogeography.E-mail:weng@tongji.edu.cn
Sunda region, located in the tropical region of Southeast Asia, is one of the three main regions of the tropical rainforests with the highest biodiversity in the world, and also the most endangered ranges of species extinction. The high biodiversity in the region was due to several reasons: ①the lucky geographical location in the warm and moist tropics, ②joint zone between the two large tectonic plates Eurasia and India-Australia, ③with abundant of islands separated with different distances. ④In the cycles of glacial-interglacial during the geological history, the variations of the temperature and the fluctuations of the sea level created opportunities for the species interactions and gene mixture, therefore resulting in the formation of new species and contributing more species to the region. In particular, during Quaternary period, the continental shelves exposed repeatedly during the glacial times, and the many islands were often merged into one or a few continuous and large territories, making the gene flows within species easier. During the interglacials, the sea-level rose and the continental shelved were submerged, and the scattered and isolated territories might make the speciation and extinction occurred more frequently. Biological refugia might be important for many species’ survival. Today, with the rapid global warming and intensive human disturbance, the refugia may be more crucial for many species to survive. However, the extinction of many species may be inevitable.
WengChengyu. The Influences of Geological History, Climatic Variations and the Environment Changes on the Terrestrial Biodiversity of Sunda Region[J]. Advances in Earth Science, 2017, 32(11): 1163-1173 https://doi.org/10.11867/j.issn.1001-8166.2017.11.1163
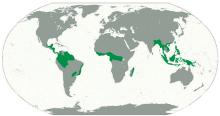
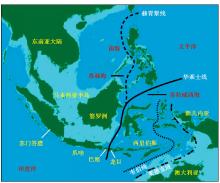
中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31]。但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33]。已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36]。而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限。这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27]。虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2)。而且差异不光存在于陆上生物,也存在于海洋生物中[40]。来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41]。华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论。
Fig.2 Geographical map of the Southeast Asia, showing the lands (green), continental shelves (light-blue region)[27] The black solid line is the Wallace’s Line, dashed lines is the other proposed boundary lines thereafter
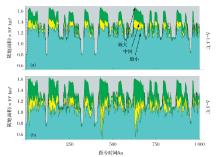
巽他的隔绝状态不是固定不变的,尤其是在第四纪时期,这就给这一区域带来了许多复杂因素。第四纪期间(大约2.6 Ma BP),全球经历了多次周期性气候冷—暖变化,即常说的冰期—间冰期气候旋回。在不同的旋回中,由于影响气候的众因素(尤其是太阳辐射量)不同,气候变化的幅度有所不同,但总的来说都是在冰期时气温降低,两极冰盖加大,伴随着的是全球海平面下降。反之,在间冰期时,海平面则上升。
Fig.3 Simulated oscillations of the land area of Sunda region in the cycles of glacial-interglacials during last one million years for two different scenarios[49] (a)Temperature dropped 1.4 ℃;(b)Temperature dropped 3 ℃
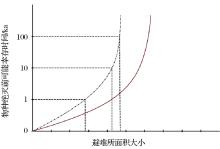
Fig.4 Conceptual relationship between size of a potential refugium and the time to extinction[61] The slope of the line is also determined by body size, generation time and the trophy level, which is represented by two different lines in the figure
Phylogeographic pattern of Rhizophora (Rhizophoraceae) reveals the importance of both vicariance and long-distance oceanic dispersal to modern mangrove distribution
Historical biogeography of Aganope, Brachypterum and Derris (Fabaceae, tribe Millettieae): Insights into the origins of Palaeotropical intercontinental disjunctions and general biogeographical patterns in Southeast Asia
[J]. Journal of Biogeography, 2014, 41(5): 882-893.
Phylogenetic systematics of the Rana signata complex of Philippine and Bornean stream frogs: Reconsideration of Huxley’s modification of Wallace’s Line at the Oriental-Australian faunal zone interface
[J]. Biological Journal of the Linnean Society, 2002, 76(3): 393-461.
[39]
Esselstyn JA, Oliveros CH, Moyle RG, et al.
Integrating phylogenetic and taxonomic evidence illuminates complex biogeographic patterns along Huxley’s modification of Wallace’s Line
[J]. Journal of Biogeography, 2010, 37(11): 2 054-2 066.
Vincent A C J. A marine fish follows Wallace’s Line: The phylogeography of the three-spot seahorse (Hippocampus trimaculatus, Syngnathidae, Teleostei) in Southeast Asia
[J]. Journal of Biogeography, 2004, 31(12): 1 975-1 985.
Biogeography and conservation in Southeast Asia: How 2.7 million years of repeated environmental fluctuations affect today’s patterns and the future of the remaining refugial-phase biodiversity
[J]. Biodiversity and Conservation, 2010, 19(4): 919-941.
Pleistocene glacial refugia across the Appalachian Mountains and coastal plain in the millipede genus Narceus: Evidence from population genetic, phylogeographic, and paleoclimatic data
A long-standing Pleistocene refugium in southern Africa and a mosaic of refugia in East Africa: Insights from mtDNA and the common eland antelope
[J]. Journal of Biogeography, 2010, 37(3): 571-581.
[59]
BornC, AlvarezN, McKEY D,et al.
Insights into the biogeographical history of the Lower Guinea Forest Domain: Evidence for the role of refugia in the intraspecific differentiation of Aucoumea klaineana
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
... Geographical map of the Southeast Asia, showing the lands (green), continental shelves (light-blue region)[27] The black solid line is the Wallace’s Line, dashed lines is the other proposed boundary lines thereafter ...
Gondwana dispersion and Asian accretion: Tectonic and palaeogeographic evolution of eastern Tethys
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Phylogeographic pattern of Rhizophora (Rhizophoraceae) reveals the importance of both vicariance and long-distance oceanic dispersal to modern mangrove distribution
1
2014
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Palaeozoic and Mesozoic tectonic evolution and palaeogeography of East Asian crustal fragments:The Korean Peninsula in context
0
2006
Tectonic framework and Phanerozoic evolution of Sundaland
1
2011
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
The palaeogeography of Sundaland and Wallacea since the Late Jurassic
0
2013
Phylogeography of the Southeast Asian stone oaks (Lithocarpus)
1
2003
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Historical biogeography of Aganope, Brachypterum and Derris (Fabaceae, tribe Millettieae): Insights into the origins of Palaeotropical intercontinental disjunctions and general biogeographical patterns in Southeast Asia
1
2014
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
West to east dispersal and subsequent rapid diversification of the mega-diverse genus Begonia (Begoniaceae) in the Malesian archipelago
1
2012
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Tectonic activities shape the spatial patchiness in the distribution of global biological diversity
1
2002
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Wallace’s line revisited: Has vicariance or dispersal shaped the distribution of Malesian hawkmoths (Lepidoptera: Sphingidae)?
1
2006
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Phylogenetic systematics of the Rana signata complex of Philippine and Bornean stream frogs: Reconsideration of Huxley’s modification of Wallace’s Line at the Oriental-Australian faunal zone interface
0
2002
Integrating phylogenetic and taxonomic evidence illuminates complex biogeographic patterns along Huxley’s modification of Wallace’s Line
1
2010
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Vincent A C J. A marine fish follows Wallace’s Line: The phylogeography of the three-spot seahorse (Hippocampus trimaculatus, Syngnathidae, Teleostei) in Southeast Asia
1
2004
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
Wallace’s Line and plant distributions: Two or three phytogeographical areas and where to group Java?
1
2011
... 中生代以前,环绕东南亚的是特提斯洋,海洋生物应该主要是特提斯洋的组成,陆地环境及其上的生物区系(也包括海岸环境的红树林)的形成则更受到欧亚板块与特提斯洋的影响[28~31].但是,在中生代后,印度与澳大利亚脱离冈瓦纳而向北推进,逐渐与欧亚大陆趋近,大约4千万年前印度与欧亚大陆直接碰撞对接,而澳大利亚虽然还没有完全对接,却也向北推挤着东南亚运移,并使一些区域抬升成为陆地或岛屿,生物之间也已开始互相交流与迁移[29~33].已直接相连的印度次大陆与欧亚大陆上的生物发生迁移与交流,并有部分向东南亚方向迁移与扩散,这个过程如果从两大板块开始接触算起,已经有数千万年的历史,这个时期可以进化出大量的新物种[34~36].而东侧的澳大利亚与东亚也一直在趋近,但大部分区域还没有直接接触,生物的交汇似乎还处于初级阶段,交流量有限.这两大生物区系主要在交界带有部分融合,但还不可能完全混合,华莱士(Wallace A R)认为它们之间存在巨大差别,从而勾勒一条界线,认为是两大区系的分界(但当时他并没有地质板块的知识,只是认识到生物学上的差异)(图2)[27].虽然后来的学者从不同生物种类中对于这两大区系的分界线的具体位置有不同的调整与认识,但总体都意识到差异的存在[37~39](图2).而且差异不光存在于陆上生物,也存在于海洋生物中[40].来自两大板块的2个区系都可能为这一区域提供了物种组成,并可能存在基因交流而共享更多的新物种[35,41].华莱士也正是根据在这一区域的工作而与达尔文几乎同时提出了进化论. ...
The hypothesis of sympatric speciation as the dominant generator of endemism in a global hotspot of biodiversity
Biogeography and conservation in Southeast Asia: How 2.7 million years of repeated environmental fluctuations affect today’s patterns and the future of the remaining refugial-phase biodiversity
... Simulated oscillations of the land area of Sunda region in the cycles of glacial-interglacials during last one million years for two different scenarios[49] (a)Temperature dropped 1.4 ℃;(b)Temperature dropped 3 ℃ ...
Pleistocene glacial refugia across the Appalachian Mountains and coastal plain in the millipede genus Narceus: Evidence from population genetic, phylogeographic, and paleoclimatic data
A long-standing Pleistocene refugium in southern Africa and a mosaic of refugia in East Africa: Insights from mtDNA and the common eland antelope
0
2010
Insights into the biogeographical history of the Lower Guinea Forest Domain: Evidence for the role of refugia in the intraspecific differentiation of Aucoumea klaineana
... Conceptual relationship between size of a potential refugium and the time to extinction[61] The slope of the line is also determined by body size, generation time and the trophy level, which is represented by two different lines in the figure ...
Historical distribution of Sundaland’s Dipterocarp rainforests at Quaternary glacial maxima




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}