1 引 言
长期以来,热带低纬度地区末次冰期时的植被与气候状态一直是科学界讨论的话题之一。关注的焦点是,此时段在东南亚暴露的巽他大陆架和南美亚马逊盆地等这些低纬地区主要被热带雨林还是被稀树草原所覆盖[1 ,2 ] ?在东南亚,Heaney[3 ] 提出“热带稀树草原廊道(savanna corridor)”的假说。主要观点是,末次冰期时稀树草原大规模扩张,其覆盖区从马来半岛向南沿着婆罗洲与爪哇岛间暴露的大陆架区域分布(图1 a)。与此相反,最近的海洋孢粉学证据表明当时古巽他大陆仍然被热带森林所覆盖[4 ,5 ] 。这一科学问题制约了我们对末次冰期时热带低纬度地区气候变化及其机制的正确认识。更重要的是,暴露的巽他大陆架面积与现代周边陆地面积相当,假如冰期时增加的陆地被热带雨林所覆盖,大气中的二氧化碳可被这部分增加的森林及其下面的土壤(主要为热带泥炭)所固定,结果可能影响我们对冰期时全球碳循环机制的理解。
末次冰期后,巽他大陆架逐渐被海水淹没,海陆变迁使得此区域的古植被重建工作较其他热带地区更为复杂。正确重建末次冰期时暴露的巽他大陆架上的植被不仅需要充足的孢粉化石记录,也需要对热带植被演替与地形及气候关系的正确理解。例如,现今的“热带稀树草原廊道”出现在巴西南部,它将南北部的亚热带雨林与热带雨林隔离开[6 ] 。对于该类植被与环境因子关系的理解有助于推测它是否可能出现在末次冰期时的古巽他大陆上。本文试图从热带稀树草原的概念入手,通过最近东南亚大量孢粉证据的总结以及与热带美洲和非洲孢粉证据的同步对比,从更广阔的视角来评估末次冰期时巽他大陆架上的植被类型。
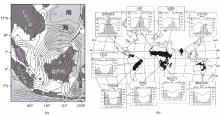
图1 东南亚地区旱季平均日降水量(a)以及现代savanna分布及其典型气候(b)[9 ]
Fig.1 Average daily precipitation during dry seasons in East Asia (a) and modern savanna distribution and its typical climate (b)[9 ]
2 热带稀树草原形成的气候条件及其组成成分
2.1 影响热带稀树草原发育的气候因素
热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] 。一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] 。其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林。非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] 。在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化。在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] 。在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] 。显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异。
在南美洲一些地区,降水也通过土壤因子(如土壤质地、保水性和矿物元素的供给等)影响savanna的分布。原因在于,沙质土壤以及隔水层上较薄的土层使得湿润季节时的水分无法长期存留,旱季土壤水分迅速散失,从而导致森林的缺失[12 ] 。
2.2 热带稀树草原的组成成分
分布在世界各地的热带稀树草原成分不同,特别是种类多样的乔木和灌木。在非洲中部,主要的乔木成分是Brachystegia ,Isoberlinia 和Julbernardia 等[6 ] 。在澳大利亚,主要乔木成分为桉树属(Eucalyptus )、白千层属(Melaleuca )和金合欢属(Acacia )。桉树属中的一些亚属,如Blakella ,Corymbaea ,Eu-desmia ,Idiogenes ,Symphyomyrtus 分布最为广泛[6 ] 。
在南美洲,乔灌木的组成成分非常丰富,种类可达774种[13 ] ,主要成分包括Caryocar brasiliense , Salvertis convallariodora , Anadanthera peregriana , Curatella americana , Tabebeuia caraiba , Byrsonima sp., Cassia sp. [14 ] 。
在亚洲,热带稀树草原被分为3种主要类型[15 ] :①落叶阔叶萨王纳(deciduous broadleaf savannahs),主要分布在南亚和东南亚(年均降水量700~2 100 mm),包括了多种群落。其中,龙脑香萨王纳群落中的乔木主要包括Shorea 和Dipterocarpus 。柚木萨王纳群落主要由Tectona 组成。混合型落叶萨王纳群落中的木本成分非常丰富,包括Anogeissus ,Lannea ,Hardwickia ,Lagerstroemia ,Pterocarpus ,Tectona ,Terminalia 和Vitex 。②Fine-leafed和spiny savannahs群落,木本植物特征为细叶和带刺,主要分布在南亚的西部和中部、东南亚的局部以及中国西南河谷地区(年均降水量400~1 000 mm)。主要成分包括Boswellia , Capparis , Acacia , Dichrostachys , Ziziphus 和Catunaregum 。③松林萨王纳(Pine savannahs),零散分布在高海拔地区(年均降水量900~3 200 mm),较其他群落更适于干燥和寒冷的环境,主要木本成分为松属[15 ] 。
综上所述,非洲savanna群落的植被组成最为简单,体现了非常干旱的气候。亚洲与美洲savanna的植被组成复杂,特别在龙脑香萨王纳群落中,Shorea 和Dipterocarpus 中的一些种也是热带低地雨林的主要成分。某些savanna类型可能与局部环境有关,例如,松林萨王纳群落更多体现了开放的森林环境。因此,基于孢粉化石判断savanna是否存在,不仅需要明确的禾本科花粉证据,还需要上述提到的乔木和灌木孢粉化石组合。
3 末次冰期时南美洲和非洲的热带雨林变化
3.1 南美洲亚马逊地区
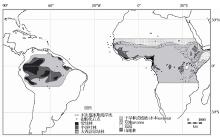
末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] 。前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成。早期的孢粉证据倾向于支持这一观点。例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] 。亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] 。然而,最近20年,越来越多的孢粉证据支持后一种解释。深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] 。在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] 。同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] 。在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] 。在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] 。
综合以上证据,末次冰期时热带南美洲的大气湿度可能有所降低,来自于现代雨林区的孢粉记录多指示当时为森林环境,而位于森林草原交错区或者高海拔的孢粉记录多表明当时为savanna环境(图2 )。现有的孢粉证据更多地指示末次冰期时热带雨林“避难所”的假设可能不成立。
3.2 非洲地区
非洲地区分布着世界最大面积的savanna。孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少。在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右。结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] 。ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 )。热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] 。除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] 。末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] 。
由于非洲热带雨林植物种类远少于热带美洲和亚洲,大量孢粉证据相对清晰地揭示了末次盛冰期以来植被带的变化特点[16 ,40 ] :撒哈拉沙漠面积扩张,热带非洲北部的植被带向南迁移,savanna的分布范围扩大,热带雨林面积减少(图2 )。
图2 末次盛冰期时南美和非洲热带地区的古植被重建[16 ]
Fig.2 Palaeovegetation reconstruction in tropical South America and Africa[16 ]
4 末次冰期时东南亚—澳大利亚北部热带地区的植被
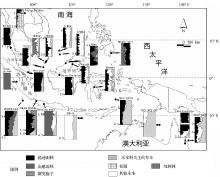
目前,对于东南亚赤道地区末次冰期时的植被与气候问题仍然存在2种截然不同的解释。最近的一些研究仍然坚持“热带稀树草原廊道”的假说,例如,Wurster等[41 ] 根据婆罗洲几个洞穴动物沉积物中的稳定性碳同位素测定,认为当时的干旱气候导致热带雨林面积大为缩小,主要分布在婆罗洲北部所谓的“避难所”。与此相反,大量的海洋和陆地孢粉记录并没有显示savanna大范围扩张,暴露的巽他大陆架与周边陆地仍被热带森林覆盖(图3 )。
在位于南海南部的17964钻孔(6° 9' N,112°12' E),冰期时段主要的花粉是热带低地雨林(含量为20%~30%),其次是红树林(含量约为20%),以禾本科为主的草本花粉含量不足20%。同样的现象出现在附近的多个深海钻孔,例如,17962,SO18300,SO18302和SO18323孔(图3 )。尽管有时禾本科花粉数量略有增加,但没有出现草本植物占据主导的时段。由于这些深海钻孔位于巽他大陆架的北部边缘,孢粉基本源于邻近的陆地,它们反映了暴露的巽他大陆架上较为广阔的区域被森林所覆盖。另外,龙脑香林作为典型的热带雨林类型,其花粉表现力极低,例如,龙脑香科(Dipterocarpaceae)花粉在其表土孢粉组合中的比例也不足5%[47 ] ,在热带深海表层沉积物中的比例甚至在1%左右[61 ] 。少量的龙脑香花粉结合其他丰富的乔木花粉可以指示热带雨林的存在。因此,上述提及的深海孢粉记录充分反映了末次冰期时暴露的巽他大陆架北部地区仍然被热带森林所覆盖(图3 )。在东南亚的赤道附近,多个孢粉记录可以提供同步的海陆证据对比,如在婆罗洲西北部的Niah(3° 49' N,113°46' E)和Kelabit(3° 34' N,115°33' E)剖面,热带雨林是孢粉组合中的主导成分,在冰期时高山雨林成分的增加较为显著,以禾本科为主的草本植物花粉略有增加,含量最多不超过30%。它们反映了冰期时温度的下降,以及个别时段湿度稍有下降(图3 )。在泰国南部的Hong Thale Song Hong(HTSH, 7° 52' N,99°28' E),禾本科花粉的高值出现在全新世,而不是在冰期(图3 )。这些孢粉证据表明,末次冰期时暴露的巽他大陆架北部及周边陆地仍然被热带森林所覆盖。
在赤道附近的Danau Sentarum湖(0° 44' N,112°06' E),来自于低地的孢粉组合显示了2.6万年以来周边地区的植被组成非常稳定,低地雨林成分占据主导地位。在冰期时,孢粉组合的变化主要表现为山地雨林成分的增加,而草本花粉保持极低比例(约5%)(图3 )。在马来半岛的Nee Soon剖面(1° 24' N,103°48' E),沼泽相沉积始于末次冰期。孢粉组合显示,此时段森林植被花粉占主导,主要表现了沼泽森林环境(swamp forest)(图3 )。在苏门答腊岛西部海拔1 535 m的Danau di-Atas钻孔(1° 04' S,100°46' E),孢粉组合显示,末次冰期以来周边陆地持续被山地森林所覆盖。气候变化主要表现为冰期时山地针叶林花粉(如Dacrycarpus 和Dacrydium )的增多,而指示干旱的草本花粉并没有增加(图3 )。在接近西太平洋暖池的菲律宾南部(MD06-3075深海钻孔,6.48° N,125.83°E),冰期时的孢粉组合主要由热带山地雨林和低地雨林花粉组成(图3 )。在苏拉威西岛的Wanda剖面(2° 33' S,121°23' E),冰期时的热带低地雨林和山地雨林成分仍然占主导,以禾本科为主的草本略有增多,可能代表气候稍有干旱(图3 )。总体上,赤道附近的孢粉记录指示了当时陆地被热带森林覆盖。
图3 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Fig.3 Pollen records in Southeast Asia-north Australia regions
澳大利亚北部与东南亚赤道附近的孢粉组合存在较大的不同,此区域冰期时禾本科花粉明显增多。在苏门答腊岛西南岸外的BAR94-42钻孔(6° 4' S,102°25' E),距今3.5~2万年时段的草本花粉含量远高于全新世,达到30%~40%(图3 )。禾本科植物扩张的情况也出现在邻近的Rawa Danau(6° 11'S, 105°59'E)和Bandung Basin(7° 00'S,108°00'E)剖面以及G6-4深海钻孔(10° 47'S,105°59'E)。在澳大利亚北部的2个海洋钻孔(SHI-9014, 5° 46' S,126°58'E;MD97-2130,12° 16.01'S,138°44.9'E),末次盛冰期至全新世时段,禾本科和莎草科(Cyperaceae)花粉占据主导,含量甚至达到95%以上。在已收集的孢粉资料中,这2个钻孔中的草本花粉含量最高。在澳大利亚东北热带地区的陆地和深海孢粉组合(Euramoo, 16°10' S,145°00' E),冰期时草本花粉含量远高于全新世。该区域的孢粉记录显示了较现代更为干燥的气候,savanna可能大范围扩张。
大多数东南亚地区的孢粉记录显示,末次冰期时热带低地雨林和山地雨林成分为主导,证明了热带森林的稳定存在。在个别山地区域,以禾本科为主的草本花粉成分有所升高,表现了气候略有干燥的时段。然而,没有显著的证据显示savanna在该区域广泛而持续的扩张。savanna可能出现在接近澳大利亚北部以及泰国北部区域,前者孢粉组合受控于Poaceae,后者为Pinus (Nong pa Kho剖面)(图3 )。由于这2个区域接近或处于现代savanna的分布区(图1 ),我们可以推断,末次冰期时savanna群落有可能在现代分布范围的基础上向外扩张,但不能够广泛替代赤道附近的热带雨林。
5 冰期时东南亚地区“savanna廊道”出现的可能性
在前面的论述中,冰期时非洲地区savanna扩张的证据主要表现为禾本科花粉在孢粉谱中占据主导。然而,在马来半岛、苏门答腊岛、爪哇岛至婆罗洲(卡里曼丹岛)的海洋与陆地孢粉化石证据组合中,我们没有发现冰期时禾本科植物占主导的记录,这说明当时暴露的巽他大陆架上似乎没有广泛的savanna分布。从马来半岛至婆罗洲,现代旱季的日均降水量为2~15 mm,如果末次冰期时savanna植被在该地区广泛分布,当时的旱季降水量可能相当于现代的中南半岛北部,较现代降低数倍(图1 a)。显然,当前的孢粉化石记录没有显示如此剧烈的湿度降低现象。
东南亚与热带美洲的孢粉证据较为相似:现代热带雨林区的孢粉记录基本没有反映末次冰期时当地草本植物广泛而持续的扩张。禾本科占主导的记录主要出现在热带边缘以及现代savanna植被出现的地区,例如,中南半岛北部和澳大利亚北部。
现代亚洲savanna群落主要分布在印度、中南半岛以及澳大利亚北部,各个区域的植被组成与环境条件均不一致(图1 )。基于现代植被与环境因子(气候、土壤和海拔)关系的植被分布区模拟提供了评估不同气候条件下savanna群落动态分布的途径[15 ] :现代非洲savanna植被区的面积最大,代表的气候最为干燥(图1 )。假设在此气候条件下,亚洲的savanna群落可能主要分布在印度和中南半岛,同时也出现在澳大利亚北部的印度尼西亚地区。然而,savanna不可能出现在赤道附近的婆罗洲、马来半岛和苏门答腊岛(图4 a)。在另外2种气候条件下,亚洲的savanna群落分布范围会极大的缩小(图4 b和c)。由此可见,末次冰期时即便现代湿热的东南亚热带雨林气候转变为干燥的非洲气候,在暴露的巽他大陆架上也难以出现savanna植被。
龙脑香科是东南亚地区分布最为广泛的热带雨林植物,Raes等[62 ] 根据317个龙脑香(Dipterocarp )种分布模型推断,末次冰期时暴露的巽他大陆架可能是龙脑香林的分布中心,具有最高的种类多样性。
因此,现有的孢粉证据与植被(savanna和Dipterocarpaceae)—气候模拟结果均显示末次冰期时暴露的巽他大陆架上不存在大面积的savanna。众多研究结果均不支持巽他大陆架上的“savanna廊道”假说。
图4 基于数据模拟的方法,在不同气候条件下亚洲savanna群落的分布概率[15 ] ;图中数字指示了不同savanna群落的位置,与之对应的景观图片请见参考文献[15]
Fig.4 Predicted distribution of savannahs in Asian based on the climate envelope[15 ] ;The numbers shown are known field locations of different Asian savannahs. Images corresponding to these numbers are shown in reference[15]
6 巽他大陆架古植被重建工作面临的挑战
在东南亚,海平面下降导致巽他大陆架广泛出露,其面积相当于婆罗洲(卡里曼丹岛)、马来半岛、苏门答腊岛和爪哇岛的面积总和[62 ] (图1 a和图3 )。由于热带非洲和南美洲并不存在广阔出露的大陆架(图2 ),这使得推测东南亚地区savanna植被是否广泛扩张较其他2个地区更为困难。主要的难点在于,出露的巽他大陆架地势较为平坦,完全不同于现代东南亚地区以山地为主的地形。例如,在婆罗洲地势平坦的区域和河口处,广泛分布淡水沼泽林(freshwater swamp forest)、泥炭沼泽林(peat swamp forest)和红树林(mangrove),这些植被主要受制于局部环境,主要组成成分不同于典型的热带低地雨林 [63 ] 。因此,在暴露的巽他大陆架上,这些可能的隐域性植被可能影响我们对典型热带低地雨林(龙脑香林)的理解。
热带植被与花粉类型极其复杂,现有的资料还不能充分提供属一级的花粉鉴定参考。例如,婆罗洲拥有10 000~15 000种植物,分布在12个植被类型中[64 ] 。考虑到科一级植物属种可分布在多个植被类型中,在这一级别上的植被重建可能会导致不同植被类型信息的丢失。如能将鉴定水平从科一级上升到属一级,则可能在孢粉组合中识别出更多的植被类型,有利于精确重建古植被和古环境。然而,花粉鉴定精度的提高还有待于大量的现代孢粉形态工作。
东南亚低地雨林是热带雨林气候的典型代表,但其孢粉组合信号还是模糊的。在南海南部,低地雨林是最为广泛的森林类型,主要由龙脑香科植物组成。然而,此类植物的花粉产量极少,即便在龙脑香林的表土孢粉组合中,其百分比也不足5%[47 ] 。此外,热带雨林中仅有很少的花粉能够传播至雨林之外[65 ] 。这使得从孢粉化石组合中正确识别热带雨林植被变得困难。
相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来。但是,这也导致了判断海洋沉积花粉源区的困难。在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] 。然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少。在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] 。河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] 。现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] 。在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区。另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑。
上述问题表明,尽管现有的证据趋向于证明末次冰期时南海南部“savanna廊道”的假说不成立,但论证工作仍需要进一步完善。
The authors have declared that no competing interests exist.
参考文献
文献选项
[1]
Bird M I Taylor D Hunt C. Palaeoenvironments of insular Southeast Asia during the Last Glacial Period: A savanna corridor in Sundaland?
[J]. Quaternary Science Reviews , 2005 , 24 (20 ): 2 228 -2 242 .
[本文引用: 1]
[2]
Bush M B. The resilience of Amazonian forests
[J]. Nature , 2017 , 541 : 167 -168 .
[本文引用: 1]
[3]
Heaney L R. A synopsis of climatic and vegetational change in southeast Asian
[J]. Climatic Change ,1991 , 19 (1/2 ): 53 -61 .
[本文引用: 1]
[4]
Sun X Li X Luo Y et al. The vegetation and climate at the last glaciation on the emerged continental shelf of the South China Sea
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2000 , 160 (3 ): 301 -316 .
[本文引用: 2]
[5]
Wang X M Sun X J Wang P X et al. Vegetation on the Sunda Shelf, South China Sea, during the last glacial maximum
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2009 , 278 (1 ): 88 -97 .
[本文引用: 2]
[6]
Huntley B J Walker B H. Ecology of Tropical Savannas
[M]. Berlin & New York: Springer , 1982 .
[本文引用: 3]
[7]
Wu Jihua Zhang Shen Jiang Yuan et al. Plant Geography [M]. Beijing : High Education Press , 2004 .
[本文引用: 1]
[武吉华 ,张绅 ,江源 ,等 . 植物地理学 [M]. 北京 :高等教育出版社 ,2004 .]
[本文引用: 1]
[8]
Lehmann C Archibald S Hoffmann W et al. Deciphering the distribution of the savanna biome
[J]. New Phytologist ,2011 , 191 (1 ): 197 -209 .
[本文引用: 1]
[9]
Archibold O W. Ecology of World Vegetation
[M]. Dordrecht: Springer Science & Business Media , 1995 .
[本文引用: 3]
[10]
Sankaran M Hanan N P Scholes R J et al. Determinants of woody cover in African savannas
[J].Nature , 2005 , 438 (7 069 ): 846 -849 .
[本文引用: 1]
[11]
Nix H A. Ecosystems of the World
[M]. New York: Elsevier , 1983 : 37 -61 .
[本文引用: 1]
[12]
Sarmiento G Monasterio M. A critical consideration of the environmenta1 conditions associated with the occurrence of savanna ecosystems in tropical America
[M]∥Golley F B,ed. Tropical Ecological Systems.Berlin: Springer ,1975 : 223 -250 .
[本文引用: 2]
[13]
SaarbriJcken M P Campo cerrado-forest or savanna?
[J]. GeoJournal ,1979 , 3 (1 ): 15 -25 .
[本文引用: 1]
[14]
Rodrigues J M Behling H Giesecke T. Differentiating vegetation types from eastern South American ecosystems based on modern and subfossil pollen samples: Evaluating modern analogues
[J]. Vegetation History and Archaeobotany ,2016 , 25 (4 ): 387 -403 .
[本文引用: 1]
[15]
Ratnam J Tomlinson K W Rasquinha D N et al. Savannahs of Asia: Antiquity, biogeography, and an uncertain future
[J]. Philosophical Transactions Royal Society B ,2016 , 371 (1 703 ): 20150305 .
[本文引用: 5]
[16]
Anhuf D Ledru M P Behling H et al. Paleo-environmental change in Amazonian and African rainforest during the LGM
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2006 , 239 (3 ): 510 -527 .
[本文引用: 4]
[17]
Hooghiemstra H van der Hammen T Neogene and Quaternary development of the neotropical rain forest: The forest refugia hypothesis, and a literature overview
[J]. Earth-Science Reviews ,1998 , 44 (3 ): 147 -183 .
[18]
Colinvaux P A. The ice-age Amazon
[J].Nature , 1979 , 278 :399 -400 .
[本文引用: 1]
[19]
Bush M B et al . Amazonian paleoecological histories: One hill, three watersheds
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2004 , 214 (4 ): 359 -393 .
[本文引用: 1]
[20]
Haffer J. Speciation in Amazonian forest birds
[J]. Science ,1969 , 165 (3 889 ): 131 -137 .
[本文引用: 2]
[21]
Van der Hammen T Absy M L Amazonia during the last glacial
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,1994 , 109 (2 ): 247 -261 .
[本文引用: 1]
[22]
Behling H Hooghiemstra H. Environmental history of the Colombian savannas of the Llanos Orientales since the Last Glacial Maximum from lake records El Pinal and Carimagua
[J]. Journal of Paleolimnology ,1999 , 21 (4 ): 461 -476 .
[本文引用: 1]
[23]
Haberle S G Maslin M A. Late Quaternary vegetation and climate change in the Amazon Basin based on a 50,000 year pollen record from the Amazon Fan, ODP Site 932
[J]. Quaternary Research ,1999 , 51 (1 ): 27 -38 .
[本文引用: 1]
[24]
Colinvaux P A et al . A long pollen record from lowland Amazonia: Forest and cooling in glacial times
[J]. Science ,1996 , 274 (5 284 ): 85 .
[本文引用: 1]
[25]
Van Der Hammen T Hooghiemstra H Neogene and Quaternary history of vegetation, climate, and plant diversity in Amazonia
[J]. Quaternary Science Reviews ,2000 , 19 (8 ): 725 -742 .
[本文引用: 1]
[26]
Cohen M C L Rossetti D F Pessenda L C R et al . Late Pleistocene glacial forest of Humaitá—Western Amazonia
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology , 2014 , 415 : 37 -47 .
[本文引用: 1]
[27]
Sifeddine A et al . A 21 000 cal years paleoclimatic record from Caçó Lake, northern Brazil: Evidence from sedimentary and pollen analyses
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2003 , 189 (1 ): 25 -34 .
[本文引用: 1]
[28]
Dupont L M Jahns S Marret F et al. Vegetation change in equatorial West Africa: Time-slices for the last 150 ka
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2000 , 155 (1 ): 95 -122 .
[本文引用: 1]
[29]
Rommerskirchen F Eglinton G Dupont L et al. Glacial/interglacial changes in southern Africa: Compound-specific δ13 C land plant biomarker and pollen records from southeast Atlantic continental margin sediments
[J]. Geochemistry, Geophysics, Geosystems ,2006 , 7 (8 ):1 -21 .
[本文引用: 1]
[30]
Dupont L M Behling H Jahns S et al. Variability in glacial and Holocene marine pollen records offshore from west southern Africa
[J]. Vegetation History and Archaeobotany ,2007 , 16 (2/3 ): 87 -100 .
[31]
Lezine A M Cazet J P. High-resolution pollen record from core KW31, Gulf of Guinea, documents the history of the lowland forests of West Equatorial Africa since 40,000 yr ago
[J]. Quaternary Research ,2005 , 64 (3 ): 432 -443 .
[32]
Shi N Dupont L M Beug H J et al. Vegetation and climate changes during the last 21 000 years in SW Africa based on a marine pollen record
[J]. Vegetation History and Archaeobotany ,1998 , 7 (3 ): 127 -140 .
[本文引用: 1]
[33]
Faye A Deblauwe V Mariac C et al. Phylogeography of the genus Podococcus (Palmae/Arecaceae) in Central African rain forests: Climate stability predicts unique genetic diversity
[J]. Molecular Phylogenetics and Evolution ,2016 , 105 : 126 -138 .
[本文引用: 1]
[34]
Maley J Brenac P. Vegetation dynamics, palaeoenvironments and climatic changes in the forests of western Cameroon during the last 28,000 years BP
[J]. Review of Palaeobotany and Palynology ,1998 , 99 (2 ): 157 -187 .
[35]
Dauby G Duminil J Heuertz M et al. Chloroplast DNA polymorphism and phylogeography of a Central African tree species widespread in mature rainforests: Greenwayodendron suaveolens (Annonaceae)
[J]. Tropical Plant Biology ,2010 , 3 (1 ): 4 -13 .
[36]
Elenga H Schwartz D Vincens A. Pollen evidence of late Quaternary vegetation and inferred climate changes in Congo
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,1994 , 109 (2/3/4 ): 345 -356 .
[本文引用: 1]
[37]
Beuning K R M Talbot M R Kelts K A revised 30,000-year paleoclimatic and paleohydrologic history of Lake Albert, East Africa
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,1997 , 136 (1/2/3/4 ): 259 -279 .
[本文引用: 1]
[38]
Ivory S J Russell J. Climate, herbivory, and fire controls on tropical African forest for the last 60ka
[J]. Quaternary Science Reviews ,2016 , 148 : 101 -114 .
[本文引用: 1]
[39]
Vincens A Garcin Y Buchet G. Influence of rainfall seasonality on African lowland vegetation during the Late Quaternary: Pollen evidence from Lake Masoko, Tanzania
[J]. Journal of Biogeography , 2007 , 34 (7 ): 1 274 -1 288 .
[本文引用: 1]
[40]
Hooghiemstra H Lézine A M et al . Late Quaternary palynology in marine sediments: A synthesis of the understanding of pollen distribution patterns in the NW African setting
[J]. Quaternary International , 2006 , 148 (1 ): 29 -44 .
[本文引用: 1]
[41]
Wurster C M Bird M I Bull I D et al. Forest contraction in north equatorial Southeast Asia during the Last Glacial Period
[J]. Proceedings of the National Academy of Sciences ,2010 , 107 (35 ): 15 508 -15 511 .
[本文引用: 1]
[42]
Penny D. A 40,000 year palynological record from north-east Thailand: Implications for biogeography and palaeo-environmental reconstruction
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2001 , 171 (3 ): 97 -128 .
[本文引用: 1]
[43]
White J C Penny D Kealhofer L et al. Vegetation changes from the late Pleistocene through the Holocene from three areas of archaeological significance in Thailand
[J]. Quaternary International ,2004 , 113 (1 ): 111 -132 .
[本文引用: 1]
[44]
Stuijts I Newsome J C Flenley J R. Evidence for late Quaternary vegetational change in the Sumatran and Javan highlands
[J]. Review of Palaeobotany and Palynology ,1988 , 55 (1/2/3 ): 207 -216 .
[本文引用: 1]
[45]
Maloney B K McCormac F G. A 30,000-year pollen and radiocarbon record from Highland Sumatra as evidence for climatic change
[J]. Radiocarbon ,1995 , 37 (2 ): 181 -190 .
[本文引用: 1]
[46]
Taylor D Yen O H Sanderson P G et al. Late Quaternary peat formation and vegetation dynamics in a lowland tropical swamp: Nee Soon, Singapore
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2001 , 171 (3 ): 269 -287 .
[本文引用: 1]
[47]
Sun X Li X Luo Y. Vegetation and climate on the sunda shelf of the South China Sea during the last Glactiation—Pollen results from station 17962
[J].Acta Botanica Sinica ,2001 , 44 (6 ): 746 -752 .
[本文引用: 3]
[48]
Hunt C O Gilbertson D D Rushworth G. A 50,000-year record of late Pleistocene tropical vegetation and human impact in lowland Borneo
[J]. Quaternary Science Reviews ,2012 , 37 : 61 -80 .
[本文引用: 1]
[49]
Jones S E Hunt C O Reimer P J. A Late Pleistocene record of climate and environmental change from the northern and southern Kelabit Highlands of Sarawak, Malaysian Borneo
[J]. Journal of Quaternary Science ,2014 , 29 (2 ): 105 -122 .
[本文引用: 1]
[50]
Bian Yeping Jian Zhimin Weng Chengyu et al. A palynological and palaeoclimatological record from the southern Philippines since the Last Glacial Maximum
[J]. Chinese Science Bulletin ,2011 , 56 (22 ): 2 359 -2 365 .
[本文引用: 1]
[边叶萍 , 翦知湣 , 翁成郁 . 末次盛冰期以来菲律宾南部气候变化的孢粉记录
[J]. 科学通报 , 2011 , 56 (19 ): 1 554 -1 561 .]
[本文引用: 1]
[51]
Anshari G Peter Kershaw A et al . Environmental change and peatland forest dynamics in the Lake Sentarum area, West Kalimantan, Indonesia
[J]. Journal of Quaternary Science ,2004 , 19 (7 ): 637 -655 .
[本文引用: 1]
[52]
Hope G. Environmental change in the late Pleistocene and later Holocene at Wanda site, Soroako, South Sulawesi, Indonesia
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2001 , 171 (3 ): 129 -145 .
[本文引用: 1]
[53]
van der Kaars S Bassinot F De Deckker P et al . Changes in monsoon and ocean circulation and the vegetation cover of southwest Sumatra through the last 83,000 years: The record from marine core BAR94-42
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2010 , 296 (1 ): 52 -78 .
[本文引用: 1]
[54]
van der Kaars S Penny D Tibby J et al . Late Quaternary palaeoecology, palynology and palaeolimnology of a tropical lowland swamp: Rawa Danau, West-Java, Indonesia
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2001 , 171 (3 ): 185 -212 .
[本文引用: 1]
[55]
van der Kaars S Dam R Vegetation and climate change in West-Java, Indonesia during the last 135,000 years
[J]. Quaternary International ,1997 , 37 : 67 -71 .
[本文引用: 1]
[56]
Wang X van der Kaars S Kershaw P et al . A record of fire, vegetation and climate through the last three glacial cycles from Lombok Ridge core G6-4, eastern Indian Ocean, Indonesia
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,1999 , 147 (3 ): 241 -256 .
[本文引用: 1]
[57]
van der Kaars S Wang X Kershaw P et al . A Late Quaternary palaeoecological record from the Banda Sea, Indonesia: Patterns of vegetation, climate and biomass burning in Indonesia and northern Australia
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2000 , 155 (1 ): 135 -153 .
[本文引用: 1]
[58]
Chivas A R et al . Sea-level and environmental changes since the last interglacial in the Gulf of Carpentaria, Australia: An overview
[J]. Quaternary International , 2001 , 83 : 19 -46 .
[本文引用: 1]
[59]
Haberle S G. A 23,000-yr pollen record from Lake Euramoo, wet tropics of NE Queensland, Australia
[J].Quaternary Research ,2005 , 64 (3 ): 343 -356 .
[本文引用: 1]
[60]
Moss P T Kershaw A P. A late Quaternary marine palynological record (oxygen isotope stages 1 to 7) for the humid tropics of northeastern Australia based on ODP Site 820
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2007 , 251 (1 ): 4 -22 .
[本文引用: 1]
[61]
van der Kaars S Pollen distribution in marine sediments from the south-eastern Indonesian waters
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology ,2001 , 171 (3 ): 341 -361 .
[本文引用: 1]
[62]
Raes N Cannon C H Saw L G et al. Historical distribution of Sundaland’s Dipterocarp rainforests at Quaternary glacial maxima
[J].Proceedings of the National Academy of Sciences ,2014 ,111 (47 ):16 790 -16 795 .
[本文引用: 2]
[63]
Ivan P. Plants and Flowers of Malaysia
[M].Singapore: Times Editions ,1991 .
[本文引用: 1]
[64]
Mackinnon K Hatta G Halim H. The Ecology of Indonesia Series Volume Ⅲ: The Ecology of Kalimantan
[M]. Hong Kong: Periplus Editions , 1996 .
[本文引用: 1]
[65]
Kershaw A P van der Kaars P Flenley J The Quaternary history of far eastern rainforests
[M]∥Bush M, Flenley J, Gosling W,eds. Tropical Rainforest Responses to Climatic Change. New York: Springer-Verlag , 2007 : 77 -114 .
[本文引用: 1]
[66]
Sun X J Li X Beug H J. Pollen distribution in hemipelagic surface sediments of the South China Sea and its relation to modern vegetation distribution
[J]. Marine Geology ,1999 , 156 (1 ): 211 -226 .
[本文引用: 1]
[67]
Luo C Lin G Chen M et al. Characteristics of pollen in surface sediments from the southern South China Sea and its paleoclimatic significance
[J]. Palaeogeography, Palaeoclimatology, Palaeoecology , 2016 , 461 : 12 -28 .
[本文引用: 1]
[68]
Moss P T Kershaw A P Grindrod J. Pollen transport and deposition in riverine and marine environments within the humid tropics of northeastern Australia
[J]. Review of Palaeobotany and Palynology , 2005 , 134 (1 ): 55 -69 .
[本文引用: 1]
[69]
Xu Q H Yang X Wu C et al. Alluvial pollen on the North China plain
[J]. Quaternary Research , 1996 , 46 (3 ): 270 -280 .
[本文引用: 1]
[70]
Xu Q H Zhang S Gaillard M et al. Studies of modern pollen assemblages for pollen dispersal deposition- preservation process understanding and for pollen-based reconstructions of past vegetation, climate, and human impact: A review based on case studies in China
[J]. Quaternary Science Reviews ,1996 , 149 :151 -166 .
[本文引用: 1]
[71]
Alqahtani F A Johnson H D et al . Nature, origin and evolution of a Late Pleistocene incised valley-fill, Sunda Shelf, Southeast Asia
[J]. Sedimentology ,2015 , 62 (4 ): 1 198 -1 232 .
[本文引用: 1]
Palaeoenvironments of insular Southeast Asia during the Last Glacial Period: A savanna corridor in Sundaland?
1
2005
... 长期以来,热带低纬度地区末次冰期时的植被与气候状态一直是科学界讨论的话题之一.关注的焦点是,此时段在东南亚暴露的巽他大陆架和南美亚马逊盆地等这些低纬地区主要被热带雨林还是被稀树草原所覆盖[1 ,2 ] ?在东南亚,Heaney[3 ] 提出“热带稀树草原廊道(savanna corridor)”的假说.主要观点是,末次冰期时稀树草原大规模扩张,其覆盖区从马来半岛向南沿着婆罗洲与爪哇岛间暴露的大陆架区域分布(图1 a).与此相反,最近的海洋孢粉学证据表明当时古巽他大陆仍然被热带森林所覆盖[4 ,5 ] .这一科学问题制约了我们对末次冰期时热带低纬度地区气候变化及其机制的正确认识.更重要的是,暴露的巽他大陆架面积与现代周边陆地面积相当,假如冰期时增加的陆地被热带雨林所覆盖,大气中的二氧化碳可被这部分增加的森林及其下面的土壤(主要为热带泥炭)所固定,结果可能影响我们对冰期时全球碳循环机制的理解. ...
The resilience of Amazonian forests
1
2017
... 长期以来,热带低纬度地区末次冰期时的植被与气候状态一直是科学界讨论的话题之一.关注的焦点是,此时段在东南亚暴露的巽他大陆架和南美亚马逊盆地等这些低纬地区主要被热带雨林还是被稀树草原所覆盖[1 ,2 ] ?在东南亚,Heaney[3 ] 提出“热带稀树草原廊道(savanna corridor)”的假说.主要观点是,末次冰期时稀树草原大规模扩张,其覆盖区从马来半岛向南沿着婆罗洲与爪哇岛间暴露的大陆架区域分布(图1 a).与此相反,最近的海洋孢粉学证据表明当时古巽他大陆仍然被热带森林所覆盖[4 ,5 ] .这一科学问题制约了我们对末次冰期时热带低纬度地区气候变化及其机制的正确认识.更重要的是,暴露的巽他大陆架面积与现代周边陆地面积相当,假如冰期时增加的陆地被热带雨林所覆盖,大气中的二氧化碳可被这部分增加的森林及其下面的土壤(主要为热带泥炭)所固定,结果可能影响我们对冰期时全球碳循环机制的理解. ...
A synopsis of climatic and vegetational change in southeast Asian
1
1991
... 长期以来,热带低纬度地区末次冰期时的植被与气候状态一直是科学界讨论的话题之一.关注的焦点是,此时段在东南亚暴露的巽他大陆架和南美亚马逊盆地等这些低纬地区主要被热带雨林还是被稀树草原所覆盖[1 ,2 ] ?在东南亚,Heaney[3 ] 提出“热带稀树草原廊道(savanna corridor)”的假说.主要观点是,末次冰期时稀树草原大规模扩张,其覆盖区从马来半岛向南沿着婆罗洲与爪哇岛间暴露的大陆架区域分布(图1 a).与此相反,最近的海洋孢粉学证据表明当时古巽他大陆仍然被热带森林所覆盖[4 ,5 ] .这一科学问题制约了我们对末次冰期时热带低纬度地区气候变化及其机制的正确认识.更重要的是,暴露的巽他大陆架面积与现代周边陆地面积相当,假如冰期时增加的陆地被热带雨林所覆盖,大气中的二氧化碳可被这部分增加的森林及其下面的土壤(主要为热带泥炭)所固定,结果可能影响我们对冰期时全球碳循环机制的理解. ...
The vegetation and climate at the last glaciation on the emerged continental shelf of the South China Sea
2
2000
... 长期以来,热带低纬度地区末次冰期时的植被与气候状态一直是科学界讨论的话题之一.关注的焦点是,此时段在东南亚暴露的巽他大陆架和南美亚马逊盆地等这些低纬地区主要被热带雨林还是被稀树草原所覆盖[1 ,2 ] ?在东南亚,Heaney[3 ] 提出“热带稀树草原廊道(savanna corridor)”的假说.主要观点是,末次冰期时稀树草原大规模扩张,其覆盖区从马来半岛向南沿着婆罗洲与爪哇岛间暴露的大陆架区域分布(图1 a).与此相反,最近的海洋孢粉学证据表明当时古巽他大陆仍然被热带森林所覆盖[4 ,5 ] .这一科学问题制约了我们对末次冰期时热带低纬度地区气候变化及其机制的正确认识.更重要的是,暴露的巽他大陆架面积与现代周边陆地面积相当,假如冰期时增加的陆地被热带雨林所覆盖,大气中的二氧化碳可被这部分增加的森林及其下面的土壤(主要为热带泥炭)所固定,结果可能影响我们对冰期时全球碳循环机制的理解. ...
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Vegetation on the Sunda Shelf, South China Sea, during the last glacial maximum
2
2009
... 长期以来,热带低纬度地区末次冰期时的植被与气候状态一直是科学界讨论的话题之一.关注的焦点是,此时段在东南亚暴露的巽他大陆架和南美亚马逊盆地等这些低纬地区主要被热带雨林还是被稀树草原所覆盖[1 ,2 ] ?在东南亚,Heaney[3 ] 提出“热带稀树草原廊道(savanna corridor)”的假说.主要观点是,末次冰期时稀树草原大规模扩张,其覆盖区从马来半岛向南沿着婆罗洲与爪哇岛间暴露的大陆架区域分布(图1 a).与此相反,最近的海洋孢粉学证据表明当时古巽他大陆仍然被热带森林所覆盖[4 ,5 ] .这一科学问题制约了我们对末次冰期时热带低纬度地区气候变化及其机制的正确认识.更重要的是,暴露的巽他大陆架面积与现代周边陆地面积相当,假如冰期时增加的陆地被热带雨林所覆盖,大气中的二氧化碳可被这部分增加的森林及其下面的土壤(主要为热带泥炭)所固定,结果可能影响我们对冰期时全球碳循环机制的理解. ...
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Ecology of Tropical Savannas
3
1982
... 末次冰期后,巽他大陆架逐渐被海水淹没,海陆变迁使得此区域的古植被重建工作较其他热带地区更为复杂.正确重建末次冰期时暴露的巽他大陆架上的植被不仅需要充足的孢粉化石记录,也需要对热带植被演替与地形及气候关系的正确理解.例如,现今的“热带稀树草原廊道”出现在巴西南部,它将南北部的亚热带雨林与热带雨林隔离开[6 ] .对于该类植被与环境因子关系的理解有助于推测它是否可能出现在末次冰期时的古巽他大陆上.本文试图从热带稀树草原的概念入手,通过最近东南亚大量孢粉证据的总结以及与热带美洲和非洲孢粉证据的同步对比,从更广阔的视角来评估末次冰期时巽他大陆架上的植被类型. ...
... 分布在世界各地的热带稀树草原成分不同,特别是种类多样的乔木和灌木.在非洲中部,主要的乔木成分是Brachystegia ,Isoberlinia 和Julbernardia 等[6 ] .在澳大利亚,主要乔木成分为桉树属(Eucalyptus )、白千层属(Melaleuca )和金合欢属(Acacia ).桉树属中的一些亚属,如Blakella ,Corymbaea ,Eu-desmia ,Idiogenes ,Symphyomyrtus 分布最为广泛[6 ] . ...
... [6 ]. ...
1
2004
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
1
2004
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
Deciphering the distribution of the savanna biome
1
2011
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
Ecology of World Vegetation
3
1995
... 东南亚地区旱季平均日降水量(a)以及现代savanna分布及其典型气候(b)[9 ]
... Average daily precipitation during dry seasons in East Asia (a) and modern savanna distribution and its typical climate (b)[9 ]
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
Determinants of woody cover in African savannas
1
2005
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
Ecosystems of the World
1
1983
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
A critical consideration of the environmenta1 conditions associated with the occurrence of savanna ecosystems in tropical America
2
1975
... 热带稀树草原是地球上重要的植被类型,英文为“savanna”,中文译为“萨王纳” [7 ] .一般被定义为分布在赤道雨林和荒漠(半荒漠)带之间,主要由C4 型草本植物和零散分布的乔灌木组成的植被类型[8 ] .其主要特点是较为开放的环境,缺乏带有密闭林冠层的森林.非洲是其最大的分布区,其次包括中美洲、南美洲、澳大利亚北部和印度,东南亚的中南半岛也有少量分布(图1 b),这种分布格局主要受到气候、水文、动物的食草作用、火和土壤性质等因素影响[9 ] .在这些因素中,水分是主导因素,包括年均降水量和降水量的季节变化.在非洲,低于650 mm的年均降水量会形成稳定的savanna[10 ] .在南美洲,年均降水量为1 000~1 500 mm[11 ] ;在东南亚,800~2 000 mm的年均降水量和5~7个月的旱季是促进savanna发育的主要因素[12 ] .显然,东南亚savanna分布区的降水量高于非洲和南美洲,这也造成了植被组成的巨大差异. ...
... 在南美洲一些地区,降水也通过土壤因子(如土壤质地、保水性和矿物元素的供给等)影响savanna的分布.原因在于,沙质土壤以及隔水层上较薄的土层使得湿润季节时的水分无法长期存留,旱季土壤水分迅速散失,从而导致森林的缺失[12 ] . ...
Campo cerrado-forest or savanna?
1
1979
... 在南美洲,乔灌木的组成成分非常丰富,种类可达774种[13 ] ,主要成分包括Caryocar brasiliense , Salvertis convallariodora , Anadanthera peregriana , Curatella americana , Tabebeuia caraiba , Byrsonima sp., Cassia sp. [14 ] . ...
Differentiating vegetation types from eastern South American ecosystems based on modern and subfossil pollen samples: Evaluating modern analogues
1
2016
... 在南美洲,乔灌木的组成成分非常丰富,种类可达774种[13 ] ,主要成分包括Caryocar brasiliense , Salvertis convallariodora , Anadanthera peregriana , Curatella americana , Tabebeuia caraiba , Byrsonima sp., Cassia sp. [14 ] . ...
Savannahs of Asia: Antiquity, biogeography, and an uncertain future
5
2016
... 在亚洲,热带稀树草原被分为3种主要类型[15 ] :①落叶阔叶萨王纳(deciduous broadleaf savannahs),主要分布在南亚和东南亚(年均降水量700~2 100 mm),包括了多种群落.其中,龙脑香萨王纳群落中的乔木主要包括Shorea 和Dipterocarpus .柚木萨王纳群落主要由Tectona 组成.混合型落叶萨王纳群落中的木本成分非常丰富,包括Anogeissus ,Lannea ,Hardwickia ,Lagerstroemia ,Pterocarpus ,Tectona ,Terminalia 和Vitex .②Fine-leafed和spiny savannahs群落,木本植物特征为细叶和带刺,主要分布在南亚的西部和中部、东南亚的局部以及中国西南河谷地区(年均降水量400~1 000 mm).主要成分包括Boswellia , Capparis , Acacia , Dichrostachys , Ziziphus 和Catunaregum .③松林萨王纳(Pine savannahs),零散分布在高海拔地区(年均降水量900~3 200 mm),较其他群落更适于干燥和寒冷的环境,主要木本成分为松属[15 ] . ...
... [15 ]. ...
... 现代亚洲savanna群落主要分布在印度、中南半岛以及澳大利亚北部,各个区域的植被组成与环境条件均不一致(图1 ).基于现代植被与环境因子(气候、土壤和海拔)关系的植被分布区模拟提供了评估不同气候条件下savanna群落动态分布的途径[15 ] :现代非洲savanna植被区的面积最大,代表的气候最为干燥(图1 ).假设在此气候条件下,亚洲的savanna群落可能主要分布在印度和中南半岛,同时也出现在澳大利亚北部的印度尼西亚地区.然而,savanna不可能出现在赤道附近的婆罗洲、马来半岛和苏门答腊岛(图4 a).在另外2种气候条件下,亚洲的savanna群落分布范围会极大的缩小(图4 b和c).由此可见,末次冰期时即便现代湿热的东南亚热带雨林气候转变为干燥的非洲气候,在暴露的巽他大陆架上也难以出现savanna植被. ...
... 基于数据模拟的方法,在不同气候条件下亚洲savanna群落的分布概率[15 ] ;图中数字指示了不同savanna群落的位置,与之对应的景观图片请见参考文献[15] ...
... Predicted distribution of savannahs in Asian based on the climate envelope[15 ] ;The numbers shown are known field locations of different Asian savannahs. Images corresponding to these numbers are shown in reference[15] ...
Paleo-environmental change in Amazonian and African rainforest during the LGM
4
2006
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
... 由于非洲热带雨林植物种类远少于热带美洲和亚洲,大量孢粉证据相对清晰地揭示了末次盛冰期以来植被带的变化特点[16 ,40 ] :撒哈拉沙漠面积扩张,热带非洲北部的植被带向南迁移,savanna的分布范围扩大,热带雨林面积减少(图2 ). ...
... 末次盛冰期时南美和非洲热带地区的古植被重建[16 ] ...
... Palaeovegetation reconstruction in tropical South America and Africa[16 ] ...
Neogene and Quaternary development of the neotropical rain forest: The forest refugia hypothesis, and a literature overview
0
1998
The ice-age Amazon
1
1979
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Amazonian paleoecological histories: One hill, three watersheds
1
2004
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Speciation in Amazonian forest birds
2
1969
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
... [20 ,24 ].同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Amazonia during the last glacial
1
1994
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Environmental history of the Colombian savannas of the Llanos Orientales since the Last Glacial Maximum from lake records El Pinal and Carimagua
1
1999
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Late Quaternary vegetation and climate change in the Amazon Basin based on a 50,000 year pollen record from the Amazon Fan, ODP Site 932
1
1999
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
A long pollen record from lowland Amazonia: Forest and cooling in glacial times
1
1996
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Neogene and Quaternary history of vegetation, climate, and plant diversity in Amazonia
1
2000
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Late Pleistocene glacial forest of Humaitá—Western Amazonia
1
2014
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
A 21 000 cal years paleoclimatic record from Caçó Lake, northern Brazil: Evidence from sedimentary and pollen analyses
1
2003
... 末次盛冰期时南美洲亚马逊热带低地雨林区是何种植被的问题,至今仍存在2种不同的解释[16 ~18 ] :①热带雨林面积减少,仅分布在局部所谓的“避难所”,其周围被savanna植被包围;②热带雨林仍然保持较为广阔的分布,但气候变化导致了其组成成分的改变[19 ] .前一种解释是Haffer[20 ] 基于鸟类的物种多样性提出的,他认为,冰期时热带雨林被savanna植被分割为众多孤立的“避难所”,多样的生境促进了许多鸟类新种的形成.早期的孢粉证据倾向于支持这一观点.例如,位于亚马逊雨林西南部(9°S, 63°W)的孢粉记录显示,冰期时savanna取代了现代的湿润森林 [21 ] .亚马逊雨林北部边缘的湖泊孢粉记录也显示当时的植被类型为savanna[22 ] .然而,最近20年,越来越多的孢粉证据支持后一种解释.深海孢粉记录(ODP932孔, 5°12.7'N, 47°1.8'W)显示了末次冰期时孢粉组合受控于木本,例如,Alchornea , Melastomataceae, Myrtaceae和Moraceae/Urticaceae,禾本科花粉较少,这证明了当时周边陆地被森林所覆盖[23 ] .在赤道雨林地区,连续的陆地孢粉记录(0°16'N, 66° 41'W)反映出亚马逊热带森林在末次冰期时是持续存在的,但气候变冷导致Podocarpus 等高山植被成分向低地入侵[20 ,24 ] .同样的结果也出现在附近3.6万年以来的孢粉记录(0° 42'S ,72° 04' W)[25 ] .在亚马逊雨林西部的森林与savanna交错区(7°55' 26″S, 63° 04' 59″W和8°46' 43″S, 63° 56'48″W,海拔1 000 m),末次冰期时的乔木花粉含量仍然很高,主要成分是Hedyosmum ,Alnus ,Fabaceae 等,显示了当时高海拔地区的森林并没有消失[26 ] .在亚马逊雨林西部的森林与savanna交错区(2° 58'S, 43° 25'W),末次盛冰期时的乔木花粉含量超过25%,尽管当时草本花粉占据主导优势并高于全新世,这一组合仍然证明了当时的森林并没有消失[27 ] . ...
Vegetation change in equatorial West Africa: Time-slices for the last 150 ka
1
2000
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Glacial/interglacial changes in southern Africa: Compound-specific δ13 C land plant biomarker and pollen records from southeast Atlantic continental margin sediments
1
2006
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Variability in glacial and Holocene marine pollen records offshore from west southern Africa
0
2007
High-resolution pollen record from core KW31, Gulf of Guinea, documents the history of the lowland forests of West Equatorial Africa since 40,000 yr ago
0
2005
Vegetation and climate changes during the last 21 000 years in SW Africa based on a marine pollen record
1
1998
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Phylogeography of the genus Podococcus (Palmae/Arecaceae) in Central African rain forests: Climate stability predicts unique genetic diversity
1
2016
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Vegetation dynamics, palaeoenvironments and climatic changes in the forests of western Cameroon during the last 28,000 years BP
0
1998
Chloroplast DNA polymorphism and phylogeography of a Central African tree species widespread in mature rainforests: Greenwayodendron suaveolens (Annonaceae)
0
2010
Pollen evidence of late Quaternary vegetation and inferred climate changes in Congo
1
1994
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
A revised 30,000-year paleoclimatic and paleohydrologic history of Lake Albert, East Africa
1
1997
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Climate, herbivory, and fire controls on tropical African forest for the last 60ka
1
2016
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Influence of rainfall seasonality on African lowland vegetation during the Late Quaternary: Pollen evidence from Lake Masoko, Tanzania
1
2007
... 非洲地区分布着世界最大面积的savanna.孢粉证据表明,末次冰期时的savanna分布范围扩大,热带雨林减少.在赤道附近的许多深海钻孔,例如,GIK16776孔(3°44'N,11°24'W)、GIK16856孔(4°48'N,3°24'E)、GeoB1008孔(6°35'S,10°19'W)和KW31孔(3°31'N,5°34'E),末次盛冰期时段的禾本科(Poaceae)花粉数量剧增,最高达到60%左右.结果指示了内陆地区savanna的广泛扩张,一定含量的雨林乔木花粉表明了热带雨林存在于赤道附近沿岸地区[28 ] .ODP1075(4°47'S,0°04'E),GeoB6518(5°35'S,11°13'E),GeoB1007(6°23'S,10°57'E)和KW31(3°31'N,05°34'E) 钻孔的孢粉记录显示末次冰期时热带雨林主要分布在刚果河下游流域和大西洋沿岸地区[29 ~32 ] (图2 ).热带雨林重要成分Podococcus 和Greenwayodendron suaveolens 的基因多样性研究和陆地孢粉证据表明,冰期时非洲的热带雨林可能收缩在中、西非洲沿岸和刚果河盆地区域的几个“避难所”[33 ~36 ] .除雨林外,在内陆高山地区仍然零散分布着常绿和半落叶乔木组成的森林[37 ,38 ] .末次冰期时,赤道南部非洲的森林植被减小程度小于北部,这可能是由于热带辐合带(Intertropical Convergence Zone,ITCZ)向南移动导致降水增多造成[39 ] . ...
Late Quaternary palynology in marine sediments: A synthesis of the understanding of pollen distribution patterns in the NW African setting
1
2006
... 由于非洲热带雨林植物种类远少于热带美洲和亚洲,大量孢粉证据相对清晰地揭示了末次盛冰期以来植被带的变化特点[16 ,40 ] :撒哈拉沙漠面积扩张,热带非洲北部的植被带向南迁移,savanna的分布范围扩大,热带雨林面积减少(图2 ). ...
Forest contraction in north equatorial Southeast Asia during the Last Glacial Period
1
2010
... 目前,对于东南亚赤道地区末次冰期时的植被与气候问题仍然存在2种截然不同的解释.最近的一些研究仍然坚持“热带稀树草原廊道”的假说,例如,Wurster等[41 ] 根据婆罗洲几个洞穴动物沉积物中的稳定性碳同位素测定,认为当时的干旱气候导致热带雨林面积大为缩小,主要分布在婆罗洲北部所谓的“避难所”.与此相反,大量的海洋和陆地孢粉记录并没有显示savanna大范围扩张,暴露的巽他大陆架与周边陆地仍被热带森林覆盖(图3 ). ...
A 40,000 year palynological record from north-east Thailand: Implications for biogeography and palaeo-environmental reconstruction
1
2001
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Vegetation changes from the late Pleistocene through the Holocene from three areas of archaeological significance in Thailand
1
2004
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Evidence for late Quaternary vegetational change in the Sumatran and Javan highlands
1
1988
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
McCormac F G. A 30,000-year pollen and radiocarbon record from Highland Sumatra as evidence for climatic change
1
1995
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Late Quaternary peat formation and vegetation dynamics in a lowland tropical swamp: Nee Soon, Singapore
1
2001
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Vegetation and climate on the sunda shelf of the South China Sea during the last Glactiation—Pollen results from station 17962
3
2001
... 在位于南海南部的17964钻孔(6° 9' N,112°12' E),冰期时段主要的花粉是热带低地雨林(含量为20%~30%),其次是红树林(含量约为20%),以禾本科为主的草本花粉含量不足20%.同样的现象出现在附近的多个深海钻孔,例如,17962,SO18300,SO18302和SO18323孔(图3 ).尽管有时禾本科花粉数量略有增加,但没有出现草本植物占据主导的时段.由于这些深海钻孔位于巽他大陆架的北部边缘,孢粉基本源于邻近的陆地,它们反映了暴露的巽他大陆架上较为广阔的区域被森林所覆盖.另外,龙脑香林作为典型的热带雨林类型,其花粉表现力极低,例如,龙脑香科(Dipterocarpaceae)花粉在其表土孢粉组合中的比例也不足5%[47 ] ,在热带深海表层沉积物中的比例甚至在1%左右[61 ] .少量的龙脑香花粉结合其他丰富的乔木花粉可以指示热带雨林的存在.因此,上述提及的深海孢粉记录充分反映了末次冰期时暴露的巽他大陆架北部地区仍然被热带森林所覆盖(图3 ).在东南亚的赤道附近,多个孢粉记录可以提供同步的海陆证据对比,如在婆罗洲西北部的Niah(3° 49' N,113°46' E)和Kelabit(3° 34' N,115°33' E)剖面,热带雨林是孢粉组合中的主导成分,在冰期时高山雨林成分的增加较为显著,以禾本科为主的草本植物花粉略有增加,含量最多不超过30%.它们反映了冰期时温度的下降,以及个别时段湿度稍有下降(图3 ).在泰国南部的Hong Thale Song Hong(HTSH, 7° 52' N,99°28' E),禾本科花粉的高值出现在全新世,而不是在冰期(图3 ).这些孢粉证据表明,末次冰期时暴露的巽他大陆架北部及周边陆地仍然被热带森林所覆盖. ...
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
... 东南亚低地雨林是热带雨林气候的典型代表,但其孢粉组合信号还是模糊的.在南海南部,低地雨林是最为广泛的森林类型,主要由龙脑香科植物组成.然而,此类植物的花粉产量极少,即便在龙脑香林的表土孢粉组合中,其百分比也不足5%[47 ] .此外,热带雨林中仅有很少的花粉能够传播至雨林之外[65 ] .这使得从孢粉化石组合中正确识别热带雨林植被变得困难. ...
A 50,000-year record of late Pleistocene tropical vegetation and human impact in lowland Borneo
1
2012
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
A Late Pleistocene record of climate and environmental change from the northern and southern Kelabit Highlands of Sarawak, Malaysian Borneo
1
2014
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
末次盛冰期以来菲律宾南部气候变化的孢粉记录
1
2011
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
末次盛冰期以来菲律宾南部气候变化的孢粉记录
1
2011
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Environmental change and peatland forest dynamics in the Lake Sentarum area, West Kalimantan, Indonesia
1
2004
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Environmental change in the late Pleistocene and later Holocene at Wanda site, Soroako, South Sulawesi, Indonesia
1
2001
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Changes in monsoon and ocean circulation and the vegetation cover of southwest Sumatra through the last 83,000 years: The record from marine core BAR94-42
1
2010
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Late Quaternary palaeoecology, palynology and palaeolimnology of a tropical lowland swamp: Rawa Danau, West-Java, Indonesia
1
2001
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Vegetation and climate change in West-Java, Indonesia during the last 135,000 years
1
1997
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
A record of fire, vegetation and climate through the last three glacial cycles from Lombok Ridge core G6-4, eastern Indian Ocean, Indonesia
1
1999
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
A Late Quaternary palaeoecological record from the Banda Sea, Indonesia: Patterns of vegetation, climate and biomass burning in Indonesia and northern Australia
1
2000
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Sea-level and environmental changes since the last interglacial in the Gulf of Carpentaria, Australia: An overview
1
2001
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
A 23,000-yr pollen record from Lake Euramoo, wet tropics of NE Queensland, Australia
1
2005
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
A late Quaternary marine palynological record (oxygen isotope stages 1 to 7) for the humid tropics of northeastern Australia based on ODP Site 820
1
2007
... 东南亚—澳大利亚北部地区的孢粉记录[42 ] ,NTSH[43 ] ,Danau di-Atas[44 ,45 ] ,Nee soon[46 ] ,17964[4 ] ,18302,18300,18323[5 ] ,17962[47 ] ,Niah[48 ] ,Kelabit(Ba钻孔)[49 ] ,MD06-3075[50 ] ,Danau Sentarum(B钻孔)[51 ] ,Wanda[52 ] ,BAR94-42[53 ] ,Rawa Danau[54 ] ,Bandung basin[55 ] ,G6-4[56 ] ,SHI-9014[57 ] ,MD97-2130[58 ] ,lake Euramoo[59 ] ,ODP820[60 ]
Pollen distribution in marine sediments from the south-eastern Indonesian waters
1
2001
... 在位于南海南部的17964钻孔(6° 9' N,112°12' E),冰期时段主要的花粉是热带低地雨林(含量为20%~30%),其次是红树林(含量约为20%),以禾本科为主的草本花粉含量不足20%.同样的现象出现在附近的多个深海钻孔,例如,17962,SO18300,SO18302和SO18323孔(图3 ).尽管有时禾本科花粉数量略有增加,但没有出现草本植物占据主导的时段.由于这些深海钻孔位于巽他大陆架的北部边缘,孢粉基本源于邻近的陆地,它们反映了暴露的巽他大陆架上较为广阔的区域被森林所覆盖.另外,龙脑香林作为典型的热带雨林类型,其花粉表现力极低,例如,龙脑香科(Dipterocarpaceae)花粉在其表土孢粉组合中的比例也不足5%[47 ] ,在热带深海表层沉积物中的比例甚至在1%左右[61 ] .少量的龙脑香花粉结合其他丰富的乔木花粉可以指示热带雨林的存在.因此,上述提及的深海孢粉记录充分反映了末次冰期时暴露的巽他大陆架北部地区仍然被热带森林所覆盖(图3 ).在东南亚的赤道附近,多个孢粉记录可以提供同步的海陆证据对比,如在婆罗洲西北部的Niah(3° 49' N,113°46' E)和Kelabit(3° 34' N,115°33' E)剖面,热带雨林是孢粉组合中的主导成分,在冰期时高山雨林成分的增加较为显著,以禾本科为主的草本植物花粉略有增加,含量最多不超过30%.它们反映了冰期时温度的下降,以及个别时段湿度稍有下降(图3 ).在泰国南部的Hong Thale Song Hong(HTSH, 7° 52' N,99°28' E),禾本科花粉的高值出现在全新世,而不是在冰期(图3 ).这些孢粉证据表明,末次冰期时暴露的巽他大陆架北部及周边陆地仍然被热带森林所覆盖. ...
Historical distribution of Sundaland’s Dipterocarp rainforests at Quaternary glacial maxima
2
2014
... 龙脑香科是东南亚地区分布最为广泛的热带雨林植物,Raes等[62 ] 根据317个龙脑香(Dipterocarp )种分布模型推断,末次冰期时暴露的巽他大陆架可能是龙脑香林的分布中心,具有最高的种类多样性. ...
... 在东南亚,海平面下降导致巽他大陆架广泛出露,其面积相当于婆罗洲(卡里曼丹岛)、马来半岛、苏门答腊岛和爪哇岛的面积总和[62 ] (图1 a和图3 ).由于热带非洲和南美洲并不存在广阔出露的大陆架(图2 ),这使得推测东南亚地区savanna植被是否广泛扩张较其他2个地区更为困难.主要的难点在于,出露的巽他大陆架地势较为平坦,完全不同于现代东南亚地区以山地为主的地形.例如,在婆罗洲地势平坦的区域和河口处,广泛分布淡水沼泽林(freshwater swamp forest)、泥炭沼泽林(peat swamp forest)和红树林(mangrove),这些植被主要受制于局部环境,主要组成成分不同于典型的热带低地雨林 [63 ] .因此,在暴露的巽他大陆架上,这些可能的隐域性植被可能影响我们对典型热带低地雨林(龙脑香林)的理解. ...
Plants and Flowers of Malaysia
1
1991
... 在东南亚,海平面下降导致巽他大陆架广泛出露,其面积相当于婆罗洲(卡里曼丹岛)、马来半岛、苏门答腊岛和爪哇岛的面积总和[62 ] (图1 a和图3 ).由于热带非洲和南美洲并不存在广阔出露的大陆架(图2 ),这使得推测东南亚地区savanna植被是否广泛扩张较其他2个地区更为困难.主要的难点在于,出露的巽他大陆架地势较为平坦,完全不同于现代东南亚地区以山地为主的地形.例如,在婆罗洲地势平坦的区域和河口处,广泛分布淡水沼泽林(freshwater swamp forest)、泥炭沼泽林(peat swamp forest)和红树林(mangrove),这些植被主要受制于局部环境,主要组成成分不同于典型的热带低地雨林 [63 ] .因此,在暴露的巽他大陆架上,这些可能的隐域性植被可能影响我们对典型热带低地雨林(龙脑香林)的理解. ...
The Ecology of Indonesia Series Volume Ⅲ: The Ecology of Kalimantan
1
1996
... 热带植被与花粉类型极其复杂,现有的资料还不能充分提供属一级的花粉鉴定参考.例如,婆罗洲拥有10 000~15 000种植物,分布在12个植被类型中[64 ] .考虑到科一级植物属种可分布在多个植被类型中,在这一级别上的植被重建可能会导致不同植被类型信息的丢失.如能将鉴定水平从科一级上升到属一级,则可能在孢粉组合中识别出更多的植被类型,有利于精确重建古植被和古环境.然而,花粉鉴定精度的提高还有待于大量的现代孢粉形态工作. ...
The Quaternary history of far eastern rainforests
1
2007
... 东南亚低地雨林是热带雨林气候的典型代表,但其孢粉组合信号还是模糊的.在南海南部,低地雨林是最为广泛的森林类型,主要由龙脑香科植物组成.然而,此类植物的花粉产量极少,即便在龙脑香林的表土孢粉组合中,其百分比也不足5%[47 ] .此外,热带雨林中仅有很少的花粉能够传播至雨林之外[65 ] .这使得从孢粉化石组合中正确识别热带雨林植被变得困难. ...
Pollen distribution in hemipelagic surface sediments of the South China Sea and its relation to modern vegetation distribution
1
1999
... 相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来.但是,这也导致了判断海洋沉积花粉源区的困难.在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] .然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少.在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] .河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] .现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] .在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区.另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑. ...
Characteristics of pollen in surface sediments from the southern South China Sea and its paleoclimatic significance
1
2016
... 相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来.但是,这也导致了判断海洋沉积花粉源区的困难.在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] .然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少.在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] .河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] .现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] .在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区.另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑. ...
Pollen transport and deposition in riverine and marine environments within the humid tropics of northeastern Australia
1
2005
... 相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来.但是,这也导致了判断海洋沉积花粉源区的困难.在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] .然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少.在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] .河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] .现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] .在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区.另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑. ...
Alluvial pollen on the North China plain
1
1996
... 相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来.但是,这也导致了判断海洋沉积花粉源区的困难.在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] .然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少.在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] .河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] .现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] .在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区.另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑. ...
Studies of modern pollen assemblages for pollen dispersal deposition- preservation process understanding and for pollen-based reconstructions of past vegetation, climate, and human impact: A review based on case studies in China
1
1996
... 相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来.但是,这也导致了判断海洋沉积花粉源区的困难.在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] .然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少.在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] .河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] .现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] .在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区.另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑. ...
Nature, origin and evolution of a Late Pleistocene incised valley-fill, Sunda Shelf, Southeast Asia
1
2015
... 相对于陆地孢粉分析,深海沉积的孢粉化石组合更能反映区域植被概况,这是由于深海沉积的孢粉由风和水流从周边陆地长途搬运而来.但是,这也导致了判断海洋沉积花粉源区的困难.在南海南部,表层沉积物、空气和海水中的孢粉分析和观测工作显示了孢粉颗粒在海洋上的传播机制[66 ,67 ] .然而,对于花粉如何从植物群落向海洋传输的环节关注依然偏少.在降水丰富的热带地区,河流是花粉向海洋传播的主导途径[68 ] .河流传播的花粉受洪水、季节等多种要素的影响,但基本反映了上游流域的植被状况[69 ,70 ] .现代海底地貌观测显示,在巽他大陆架上存在多条古河道,证明当时暴露的巽他大陆架上存在着多条大河,包括North-Sunda河、Siam河和Johore河等[71 ] .在上述古河流河口处的孢粉沉积可能反映上游较为广阔区域的植被面貌,然而,当前的深海钻孔并没有覆盖这些古河口区.另外,海洋孢粉化石数据的正确解释,热带雨林孢粉信号的揭示还有待于更多的现代孢粉传播机制研究予以支撑. ...





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}