1 引言
全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] 。通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] 。前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式。其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] 。而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] 。这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主。
早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] 。目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道。本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例。
2 地质背景
石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主。上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] 。志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] 。上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内。受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b)。本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育。
石牛栏组上覆地层为韩家店组(S1-2 hj ),岩性以黄色泥岩夹亮晶砾屑灰岩为主,含少量珊瑚、层孔虫、腕足类及头足类化石,与下伏石牛栏组呈整合接触。石牛栏组下伏地层为龙马溪组(S1 l ),沉积灰色—灰黑色的含笔石页岩夹少量泥质粉砂岩,含丰富的笔石化石及少量头足类化石[25 ] 。生物礁出现在石牛栏组中上段地层中,其生物种类有珊瑚、层孔虫、腕足类和双壳类等(图1 c)。自下而上石牛栏组共可分为4个段[23 ] :第一段为灰色—深灰色泥质灰岩、瘤状灰岩,夹褐黄色含钙粉砂质泥岩,含少量生物化石和虫迹,石牛栏组早期继承了龙马溪组浅水陆棚相沉积,泥质成分含量高,代表了一种相对安静的水体环境;第二段岩性以黄色及灰色粉屑灰岩和浅灰色生屑灰岩为主,夹少量粉砂泥质灰岩,含少量生物化石,层面有虫孔和虫迹,代表了一种相对动荡的沉积环境;第三段岩性为灰黑色生物礁灰岩、灰色—浅灰色生屑灰岩及生物灰岩,含大量珊瑚(蜂巢珊瑚目及链珊瑚)、层孔虫、腕足类及双壳类,上部有紫红色灰岩砾块,夹有泥质及粉砂质条带;第四段岩性为灰色砾屑灰岩及生屑灰岩,夹少量泥质条带,具小型斜层理,代表了一种相对安静的沉积环境。总体上,石牛栏组第一段至第四段显示了从一种相对低能的沉积环境向高能环境,再向低能环境转变的一个过程。
图1 区域地质背景及地层分布图[17 ,24 ] ;(c) 石牛栏组岩性柱状图(重庆万盛机枪台剖面)
Fig.1 Regional geological background and stratigraphic distribution[17 ,24 ] ; (c) The Shiniulan Formation lithology (the Jiqiangtai section of Wansheng, Chongqing)
3 生物礁特征及发育过程
3.1 石牛栏组生物礁出露特征
石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好。生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回。其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等。从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段。横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 )。而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关。石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] 。石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积。石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] 。这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁。
图2 上扬子地区石牛栏组地层对比图
Fig.2 Coenocorrelation of the Shiniulan Formation in the Upper Yangtze RegionFig.3 a~h; ②Fig.4 a~f;③ Fig.7 a,b; ④Fig.7 c; ⑤Fig. 7 d
上扬子海洋早志留世石牛栏组生物礁分布较为广泛,主要为一些小型点礁,生物礁规模不大,厚10~30 m,延伸约10 km,礁体间存在沉积物(包括灰泥、砂岩、泥岩和少量生物碎屑)和生物骨架。石牛栏组生物礁呈层状或枕状突起与周围岩层较好区别,突起不高,礁体为一些生物格架灰岩或生物碎屑灰岩。造礁生物主要为横板珊瑚(蜂巢珊瑚目及链珊瑚为主)、四射珊瑚和层孔虫(图3 a~e),附礁生物为腕足类、三叶虫、藻类、海百合、苔藓虫和双壳等(图3 a,b,f),具良好的生物抗浪格架及格架孔。在机枪台露头剖面上,生物礁呈层状、席状夹于碳酸岩沉积物中,这些层状碳酸岩沉积物包括泥质灰岩、颗粒灰岩等,交替出现在生物礁灰岩间(图3 g),生物礁灰岩横向上厚5 m左右,长达27 m(图3 h)。
微观上,造礁生物珊瑚—层孔虫广泛发育于生物岩层中,与其他生物与沉积物构成生物礁复合体的主要部分(图4 )。生物礁的建设者四射珊瑚(如扭心珊瑚)单体分布于生物岩体空间中,呈近圆状和椭圆状(图4 a);蜂巢珊瑚复体横向上由不规则的六边形组成,纵向上向外或向上生长为放射状的复合体(图4 b)。生物礁的另一个构建者层孔虫也出现于生物礁岩层中,有零散呈板状、上凹板状、扩展状及层状的生长样式分布,也有大量的层孔虫聚集呈网格状分布于在生物岩层空间中,或因外力作用而呈零星碎片分布(图4 c, d)。双壳动物(Bi)碎屑嵌于大量方解石(C)颗粒中和藻类碎屑在生物礁灰岩中(图4 e, f)。造礁生物空间由深灰色泥晶灰岩充填,方解石晶体呈晶粒镶嵌式胶结,生物体周围有泥岩和碎屑,以及一些细小的矿物晶体(如黄铁矿)(图4 b, f)。
图3 上扬子地区石牛栏组生物礁灰岩特征Palaeofavosites shiniulanensis )属于蜂巢珊瑚目(Favositida)古巢珊瑚属(Palaeofavosites )。个体呈扩散状排列,个体横切面多角形,大小不一致,大者6~8边形,在大个体周围排列小个体;横板完整呈水平状,少数呈倾斜。(e)拉贝希层孔虫(未定种)(Labechia sp.)共骨为块状。骨骼由连续的平缓的波状细层和短支柱组成。局部泡沫板支柱主要由细层向下弯折而成, 未达底层。支柱的横断面为圆形;(f) 大量的生物碎屑如珊瑚、藻类及苔藓虫在生物礁灰岩,机枪台剖面,100 m;(g) 点礁内部含大量珊瑚、层孔虫和藻类等生物,机枪台剖面,105 m;(h) 生物礁横向上厚约5 m,长约27 m,机枪台剖面
Fig.3 Biohermal limestone characteristics of the Shiniulan Formation in the Upper Yangtze RegionPalaeofavosites shiniulanensis is arranged in a diffuse arrangement. The individual transverse section is polygonal, the large 6~8 sides in shaped, and the small individuals are arranged around the large individuals. The tabula are completely horizontal and a few are tilted. (e) Skeleton of Labechia sp. is massive and consists of gentle wavy lamina and short backbones. The local cystose pillars are mainly bent down from a thin layer and not reach the bottom layer. (f) A large number of biodetritus such as stromatoporoids, tabulate corals, algae and bryozoans in biohermal limestone of the Jiqiangtai section, 100 m; (g) Patch reefs. It contains a lot of corals, stromatoporoids and algae in the Jiqiangtai section, 105 m. (h) the layered reefs extend laterally by 27 m and 5 m in thickness of the upper part in the Shiniulan Formation of the Jiqiangtai section
图4 上扬子地区生物礁微观镜下特征
Fig.4 Microscopic photographs of reefs in the Upper Yangtze Region
3.2 生物礁发育过程
生物礁的生长规模从侧面上可以反映生物礁的成礁过程,一般来说,发育较好的生物礁共可分为5个成礁过程:定殖期(奠基期)、拓殖期、泛殖期、衰亡期和礁盖期(盖层期)。根据石牛栏组生物礁纵向及横向发育特征,选取典型习水县羊九剖面进行分析(图5 a),大致将其成礁过程分为定殖阶段、拓殖阶段、泛殖阶段、衰亡阶段共4个生长阶段[28 ,29 ] 。
图5 上扬子海洋早志留世生物礁发育过程模式图
Fig.5 The developmental pattern of reef (Early Silurian) in Paleo-Ocean of Upper Yangtze Region
(1)定殖阶段
该阶段是石牛栏组生物礁的初发育时期,在生物或非生物作用下,不稳定或软质的基底变为稳定或硬质基底[29 ] ,其大体对应石牛栏组第一段。此阶段腕足类、棘皮类(如海百合)等生物用肉柄、壳刺或其他固着器官固着于不稳定或较软质的基质之上,并使基质处于稳定状态。之后,横板珊瑚及层孔虫迁移至此,共同为礁体的发育提供一个较稳定的基底。代表这个时期的典型岩石类型是泥晶灰岩、钙质泥页岩与灰质泥岩,以及一些含棘皮类、腕足类等化石的瘤状灰岩,生物种类含有少量海百合、腕足类、珊瑚等(图5 b-I)。
(2)拓殖阶段
该阶段造礁生物已占据稳定的基底发育,造礁生物开始显现分异化,造礁生物占据该地并大量繁衍,大体上对应石牛栏组第二段。随着沉积物的堆积逐渐增加,海水进一步变浅,开始堆积大量的碳酸盐沉积物。之前在此定居的造礁生物(块状、枝状横板珊瑚及层孔虫)在良好的生存环境进一步生长发育。在此阶段造礁生物的分异度不高,但它们的功能分化已开始显现,即块状、枝状横板珊瑚及层孔虫作为沉积物的造架者,块状层孔虫作为礁架的建造者及沉积物的黏结者开始出现。代表这个时期的典型岩石类型是生物灰岩、富含生物碎屑及砂屑灰岩和泥质灰岩,含少量泥质条带,有些剖面含白云质灰岩。灰泥及生物碎屑在固着生物体之间大量沉积下来,生物作用在此阶段的沉积速度明显高于没有生物礁的地区,初期礁体地貌隆起已开始显现(图5 b-II)。
(3)泛殖阶段
在这个阶段块状、枝状横板珊瑚和层孔虫的发育进入鼎盛时期,大体上对应石牛栏组第三段。在菌藻类黏结岩和珊瑚及层孔虫造架岩的共同作用下,礁体具有初步的抗浪功能,逐渐形成隆起于周缘的地貌,礁体环境进一步变浅[29 ] 。造礁生物和联结生物(海绵、层孔虫、钙藻等)等大量繁殖,造礁生物分异化明显增强,礁体纵横向发育到了最大规模。代表这个时期的典型岩石类型是灰色块状生物礁灰岩以及亮晶生屑灰岩,局部可见沥青和油浸。此时,生物礁的礁前、礁核与礁后环境已基本显现(图5 b-III)。
(4)衰亡阶段
在这个阶段石牛栏组生物礁发育趋于衰亡,大体上对应石牛栏组第四段。随着造礁生物及附礁生物的繁盛生长,礁体迅速扩大,当礁体扩长达平均海平面时,形成极度浅水环境,生物礁间歇性暴露于水面[30 ] 。造礁生物的生长发育受生存环境的限制,造成窄盐性造礁生物群大量死亡,剩下少数的广盐性生物(如腹足类、藻类)生存下来,此时使生物分异化变弱[31 ] 。此时沉积物多带有潮坪环境特征,生物数量和种类大量减少,在纵向上礁体停止发育,顶部有少许白云石化现象。代表这个时期的典型岩生物碎屑灰岩及亮晶生屑灰岩、白云岩等(图5 b-IV)。
4 早志留世生物礁发育过程制约机制
在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关。根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响。
4.1 海平面升降对生物礁发育过程的制约机制
海平面升降是影响石牛栏组生物礁生长演化的一个重要因素。海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] 。海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等。海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化。反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] 。
图6 上扬子海洋早志留世海平面变化与生物礁发育过程
Fig.6 Sea level changes and developmental stage of reef (Early Silurian) in Paleo-Ocean of Upper Yangtze Region
以石牛栏组羊九剖面和机枪台剖面生物礁为例,其生长可分为2期(早期与晚期)成礁旋回,对应可把石牛栏组分为一段和二段(图6 a)。一个完整的成礁旋回显示序列可分为6个阶段,分别对应沉积的岩石类型(图6 b)和海平面的相对变化(图6 c):I阶段分布在石牛栏组一段下部,延续了龙马溪组岩性特征,岩性为页岩、粉砂质泥岩,生物为一些藻类、微生物和腕足动物,生物活动性较弱,可见水平层理,海水位于高水位,为浅水陆棚相沉积。II阶段分布在石牛栏组一段中下部和二段下部,为碳酸盐岩形成初期阶段,生物的作用微弱,可见一些海百合茎,岩层由瘤状灰岩和少量的泥灰岩、泥岩组成。此时海水处于较高水位期,但海平面已开始缓慢下降,为台地边缘斜坡相中的下斜坡亚相沉积。III阶段分布在石牛栏组一段中部,对应生物礁的定殖阶段,珊瑚、层孔虫、腕足类及棘皮类等生物开始大量繁殖,岩性以生物碎屑灰岩为主。海水持续下降,为台地边缘浅滩相生屑滩亚相沉积。IV阶段分布在石牛栏组一段中上部和二段中部,受海水搅动的影响,沉积物沉积速度开始加快,岩性以角砾灰岩夹杂生屑灰岩条带。海平面下降到一定时期开始回升,为台地边缘斜坡相中的上斜坡亚相沉积。V阶段分布在石牛栏组一段顶部和二段顶部,造礁生物珊瑚及层孔虫生长达到鼎盛阶段,附礁生物的多样性也达到最大值,生物礁形成小型生物礁灰岩。此时海平面逐渐上升达到一个值后又缓慢开始下降,为台地边缘生物礁相障积岩亚相沉积。VI阶段分布在石牛栏组二段中上部,生物礁的生长已到了衰退期,海平面的快速下降及障积岩大量堆积,生物礁暴露于水面,岩性主要为泥岩及泥质灰岩,发育丰富的层理构造。海水持续下降最终导致生物死亡,为局限台地相泻湖亚相沉积。
石牛栏组沉积期为早志留世,该时期正值全球古气候史上的大间冰期,此时海侵范围较小,温度升高,加里东运动使地壳上升,导致整个早志留世上扬子区海水变浅,海平面相对下降。石牛栏组早期至晚期,海平面长期处于下降的趋势,导致石牛栏组生物在晚期死亡,最终生物礁群落演化停滞;水体的这种变化导致自埃隆晚期后,在此区域内长时间无生物礁发育。这样的变化同时也影响着生物礁形态的发育、分布及规模的大小,导致生物礁在上扬子地台未形成较大规模,而多为点礁分布。
4.2 海水的温度及盐度对生物礁发育过程的制约机制
国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] 。据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] 。为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成。
(1) 古海水温度特征
δ18 O值变化可较好地反映古海洋环境的温度变化。一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] 。但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] 。因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] 。根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰。此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] 。
T =17.0 -4.52 (δ18 O calcite -δ18 Owater )+ 0.03(δ18 O calcite -δ18 O water )2 , (1)
式中:T ,δ18 O calcite 和δ18 Owater 分别代表古温度(℃)、碳酸岩形成时氧同位素值(PDB标准)和海水中的氧同位素值(SMOW标准)。
通过计算羊九村骑龙剖面石牛栏组δ18 O calcit 校正后的数值计算古温度(表1 ),其结果比较理想。
但由于碳酸盐岩总体上仍然受成岩作用影响,因此这些值并不能完全代表当时海水温度,只具有一定的参考价值。上述公式计算可得,石牛栏组沉积期海水温度为18.58~29.58 ℃,平均温度为22.46 ℃。
(2) 古海水盐度特征
古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] 。本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化:
Z =2.048 (δ13 C+50) +0.498 (δ18 O +50), (2)
式中:δ13 C和δ18 O均采用PDB标准值;Z >120‰时为海相石灰岩;Z <120‰,时为淡水石灰岩。
选取羊九村骑龙剖面共计51个样品点,采集均为微晶灰岩,受成岩作用影响小,分析δ18 O和δ13 C值变化能更准确指示古盐度与古温度(表1 )。纵向上,δ13 C值与δ18 O值呈多旋回变化。其中δ13 C值大小范围为-0.31‰~2.81‰,平均值为+1.84‰;δ18 O大小范围为-11.09‰~-8.7‰,平均值为-9.55‰。通过计算,Z 值在骑龙剖面均大于120‰,分布范围为122.05‰~127.90‰。
海相碳酸盐岩中的碳、氧等同位素演化规律研究提供了志留纪石牛栏组沉积环境的重要信息,能较为有效地反映上扬子区早志留世古海洋生物礁生存环境。根据上述温度与盐度的计算数据分析可知,最适宜生物礁生长的Z 值范围为124‰~127‰,当Z >130‰,水体环境中盐度增加或温度上升,生物将会面临死亡;最适宜生物礁生长的温度为22.13~29.58 ℃,温度过低过高都将影响各类生物的生存与造礁。这些分析表明,上扬子早志留世海洋主要为温暖或炎热的热带及亚热带气候,盐度较低的浅海沉积环境。
4.3 外来物源搅动对生物礁发育的影响机制
前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] 。石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c)。这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] 。碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] 。通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响。
图7 外来物源搅动影响下的沉积岩体
Fig.7 The sedimentary rock under the influence of external sources agitation
总的来说,外来物源的搅动大部分不利于生物繁殖,这主要由于生物礁的生长需清澈洁净、安静且富氧的水体环境,外来物源的搅动影响到水质的更新及循环。但另一方面,我们认为外来物源的搅动对生物礁的制约并不是绝对的,某种程度上也促进了生物礁的形成。如季风带来的物质中可能含有大量养分,可促进生物礁中藻类生长;另外,洋流中暖流的流动增加了水体的温度,大量喜温的生物得以大量繁殖。
4.4 构造运动对生物礁发育过程的制约机制
上扬子区志留纪生物礁的演化与所处区域的构造环境与构造运动相关,自埃隆期后,该区域地层长期暴露于水体以外,制约生物礁在志留纪发展与演化,直至二叠纪再次被水体淹没,生物礁才得以发育与演化。研究区石牛栏组的岩性、生物礁群落、沉积及其碳氧同位素含量等证实,在石牛栏组沉积期间,黔中古陆以北和东北部的上扬子海域平缓海底发生过短暂但大范围的隆升,使海水退却遭遇风化剥蚀[11 ] 。
前人研究表明,早志留世鲁丹期(龙马溪组)上扬子地台为较深水体环境,在埃隆晚期,研究区所处古大陆上升,古陆北缘逐渐向北扩展,海水的升降受其影响而变化,影响着石牛栏组生物礁的生长与演化。伴随着华夏古陆剥蚀量的增加,细碎屑物快速堆积使以往相对平静、以灰泥沉积为主的海底环境(石牛栏组上部)彻底丧失,广泛发育泥质海底,变为时而富氧、时而贫氧的沉积环境[11 ] 。在这种环境下,生物群落生长发育受到影响,生物间歇性大量死亡,整个石牛栏组生物礁系统遭受破坏。从全球范围看,埃隆晚期被大多数学者认为是全球海平面的上升期[52 ,53 ,54 ] ,若对应中国南方生物层,大致埃隆晚期海水达到最高位。如此看来,上扬子地区埃隆晚期海平面也应处于上升状态中,但实际情况却与之相反,该时期整个区域海水呈现大幅度变浅的现象。这可能有4个方面原因[11 ] :①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升。正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡。实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素。因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因。
5 讨 论
由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异。生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低。志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育。珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] 。中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现。Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中。晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加。晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现。总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟。
中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段。与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] 。在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] 。事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异。
加拿大东部安蒂科斯蒂岛的Menier组(埃隆晚期,兰多维列世)地层发现了生物礁,其造礁生物主要是四射珊瑚与横板珊瑚,以及较少的层孔虫,附礁生物包括海百合、腕足类、绿藻、蓝藻等[4 ] ,因其造礁生物与地层岩性等都与石牛栏组生物礁保持高度的相似性,我们认为其相当于同期的石牛栏组生物礁。通过分析可知,两地的生物礁存在几个方面的共同点:①生物礁均生长在低纬度热带大陆架的边缘地带,拥有良好的气候和适宜的海水深度;②造礁生物均为珊瑚和层孔虫,附礁生物为藻类、腕足类、海百合等,生物类型体现出高度相似性。但实际上,Menier组与石牛栏组生物礁在特征上存在很大差异性,主要体现在(图8 ):①Menier组生物礁的珊瑚、层孔虫、钙化藻呈现最大的多样性,但腕足动物、头足类和海百合的多样性却很低;石牛栏组珊瑚—层孔虫虽然大量繁殖,但整体生物多样性却较低,两地生物的种属也大相径庭。②Menier组大多数生物礁露头呈现不规则圆形,个体之间相对密集,生物礁个体或聚集的群体在露头上展布方向都是随机的,规模较大;而石牛栏组生物礁的生长模式以点礁为主,含少量生物丘和生物层的类型,规模较小。③Menier组生物礁的生物外壳、生物骨骼及层间沉积物等主要是由粉砂质灰岩至砂质灰岩构成,泥岩成分几乎没有,生物骨骼几乎完整,只在一些较浅的水域受风暴影响出现生物碎屑。而石牛栏组生物礁生物碎屑在下段地层中显示很多,生物外壳、生物骨骼及层间沉积物除一些砂质灰岩之外,还会有泥质成分。④安蒂科斯蒂岛特里奇期延续Menier组珊瑚—层孔虫礁生长趋势,在上部地层Chicotte组发育了更大规模且生物多样性更高的生物礁[66 ,67 ] ;而上扬子地台石牛栏组发育一定规模的生物礁后,特里奇期没有相应规模的生物礁发育或仅发育规模数米的小型点礁[68 ] 。
图8 加拿大Menier组与上扬子地区石牛栏组生物礁对比[4 ]
Fig.8 Comparison of the Menier Formation and the Shiniulan Formation in the Upper Yangtze Region[4 ]
造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响。奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响。早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力。鲁丹阶早期龙马溪组灰岩较薄,生物种类少。上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物。但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] 。②陆源碎屑的注入。Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] 。③海平面的升降。Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] 。埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] 。④构造运动与海水深度。志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境。地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] 。Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性。
6 结 论
(1) 上扬子区兰多维列世石牛栏期生物礁主要发育在大陆架碳酸盐缓坡地带,造礁生物为横板珊瑚、四射珊瑚和层孔虫,附礁生物包括苔藓虫、腕足类、头足类、藻类、海百合及双壳类等。三维空间展布上以小型点礁为主,生物礁生长发育分为定殖、拓殖、泛殖、衰亡4个生长阶段。
(2) 石牛栏组生物礁很大程度上受外来物源搅动、海平面升降及海水温度和盐度等因素的影响,这些因素是埃隆晚期加里东构造运动导致古陆的隆起上升和扩展所引起的。石牛栏组生物礁生长演化受这些因素的制约,在埃隆晚期大量死亡,与全球志留纪同期生物礁相比,这是上扬子地台志留纪晚期未能大规模成礁的主要原因。
(3) 生物礁在早志留世埃隆期的分布是全球性的,主要分布在气候温暖(20~28 ℃)的南北纬25°~30°碳酸盐缓坡上,同期发育的生物礁主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块。早志留世埃隆期生物礁的生长与演化,标志着自奥陶纪大规模生物灭绝后,上扬子地台志留纪生物礁大规模复苏。这不仅对志留纪华南板块生物的类型及演化研究极具参考价值,也对全球志留纪生物礁研究有重要意义。
The authors have declared that no competing interests exist.
参考文献
文献选项
[1]
Li Yue Kershaw S Chen Xu Control of carbonate sedimentation and reef growth in llandovery Sequences on the Northwestern Margin of the Yangtze Platform, South China
[J]. Gondwana Research , 2004 , 7 (4 ):937 -949 .
DOI
URL
[本文引用: 2]
[2]
Copper P Silurian and Devonian reefs: 80 million years of global greenhouse between two ice ages
[J]. Special Publications 2002 (72 ):181 -238 .
DOI
URL
[本文引用: 1]
摘要
The Silurian-Devonian (mid-Paleozoic), a time of periodic, exceptional sea level highstands, vast epicontinental sea lanes, and global greenhouse climates well above Holocene norms, also identifies the maximal extent of Phanerozoic metazoan reef development and the acme of coral and sponge reef biodiversity. Two peaks are identified for reef distribution, a mid-Silurian (Wenlock) maximum and a Mid- to Late Devonian (Eifel鈥揋ivet鈥搈iddle Frasne) maximum, with reefs spread to latitudes as high as 45 to 60 degrees, and major barrier-platform tropical reef belts stretched over more than 2000 km. Reef abundance broadly matched patterns of rise and fall in generic biodiversity for the major metazoan reef builders (corals and stromatoporoids) and algae during this 80 million year episode. The mid-Paleozoic reef community originated in the Late Ordovician, taking over from the Cambro-Ordovician mudmound-dominated reef spectrum. It was only modestly affected by the glaciation-induced end-Ordovician (Himantian) mass extinction events. Late Devonian reef extinction losses were initiated at the Givehan鈥揊rasnian boundary by major declines in coral, stromatoporoid, and brachiopod benthic components. Reefs then experienced a second global expansion in the middle Frasnian, but with reduced faunal diversity. Catastrophic reef declines began in the late Frasnian rhenana conodont Zone and continued through the final linguiformis Zone, with events probably lasting ca.1 Myr. The coral-stromatoporoid reef community was eliminated worldwide, with surviving patch reefs and reefal mud mounds in the Famennian constructed by consortia of calcimicrobes, lithistid sponges, green and red algae, and foraminiferans. The surviving Famennian corals were primarily solitary, deep-water forms that played no major reef role. The last stromatoporoids died out within the Strunian praesulcata Zone, at the Devonian-Carboniferous boundary. Widespread organic-rich carbonates of Givetian through Famennian age, normally taken as indicative of anoxia, do not show any correlation with local or global reef declines. The Frasnian-Famennian extinction events for reefs and reef faunas appear to have been second only to the end-Permian reef losses in terms of severity. Metazoan reef extinction in the Late Devonian best parallels evidence for Famennian glaciations, loss of reef accommodation space as sealevels felt, and increasing oxygenation of the atmosphere via the evolution of the first pteridophyte rainforests, forcing the terminal Famennian icehouse phases.
[3]
Antoshkina A I Late Ordovician-Early Silurian facies development and environmental changes in the Subpolar Urals
[J]. Lethaia , 2008 , 41 (2 ):163 -171 .
DOI
URL
[本文引用: 1]
[4]
Copper P Jin J Early Silurian (Aeronian) East Point Coral patch reefs of anticosti island, Eastern Canada: First reef recovery from the ordovician/silurian mass extinction in Eastern Laurentia
[J]. Geosciences , 2012 , 2 (2 ):64 -89 .
DOI
URL
[本文引用: 9]
[5]
Wang Guan Li Yue Global reef recovery after the end-Ordovician extinction: Evidence from late Aeronian coral-stromatoporoid reefs in South China
[J]. GFF , 2014 , 136 (1 ):286 -289 .
DOI
URL
[本文引用: 4]
[6]
Kershaw S Mõtus M Palaeoecology of corals and stromatoporoids in a Late Silurian biostrome in estonia
[J].Palaeontologia Polonica , 2016 , 61 (1 ):33 -50 .
DOI
URL
[本文引用: 3]
摘要
A middle Ludlow biostrome at Katri, western Estonia, the richest accumulation of corals and stromatoporoids in Estonia, is partly exposed in a coastal section. The fully marine biostrome consists of five fossiliferous layers of carbonate skeletons, grouped into Facies 1 (grainstone-packstone, Layers 1, 3, 5) and Facies 2 (wackestone, interbedded Layers 2, 4). Pressure solution degraded original sedimentary relationships and morphologies of stromatoporoids, tabulates and rugosans which constructed the biostrome, but the two facies have major faunal differences. Facies 1 is rich in stromatoporoids “Stromatopora” bekkeri and Plectostroma scaniense (low to high domical up to ca. 30 cm in basal length); and tabulate Favosites forbesi (bulbous to high domical up to ca. 25 cm wide). In Facies 2, all three taxa are less common and much smaller. Instead, the most abundant stromatoporoid is laminar Syringostromella borealis up to 30 cm basal length; the most abundant coral is erect branching Laceripora cribrosa, as...
[7]
Färber C Munnecke A Gypsum evaporites in a patch reef of the upper Slite Group in the Silurian (Wenlock) of Gotland, Sweden
[J]. GFF , 2014 , 136 (1 ):75 -79 .
DOI
URL
[本文引用: 2]
[8]
Sandström O Kershaw S Paleobiology, ecology, and distribution of stromatoporoid faunas in biostromes of the Mid-Ludlow of gotland, Sweden
[J]. Acta Palaeontologica Polonica , 2008 , 53 :293 -302 .
DOI
URL
[本文引用: 2]
[9]
Kaminskas D Micheleviǒius D Blažauskas N New evidence of an early Pridoli barrier reef in the southern part of the Baltic Silurian basin based on three-dimensional seismic survey, Lithuania
[J]. Estonian Journal of Earth Sciences , 2015 , 64 (1 ):47 -55 .
DOI
URL
[本文引用: 2]
[10]
Watts N R Riding R Growth of rigid high-relief patch reefs, Mid-Silurian, Gotland, Sweden
[J]. Sedimentology , 2010 , 47 (5 ):979 -994 .
DOI
URL
[本文引用: 1]
摘要
Patch reefs up to 35 m thick and generally 100–150 m wide, separated by bedded inter-reef sediment, dominate the H02gklint Formation (Lower Wenlock) of north-west Gotland. The spacing between adjacent patch reefs is variable, but is commonly 150–350 m. The H02gklint is a shallowing sequence, and the patch reefs exhibit a well-developed vertical succession: (1) Axelsro-type patch reefs developed in the underlying Visby Formation; (2) halysitid tabulates capped by laminar stromatoporoids; (3) domical and bulbous stromatoporoids and red algae; (4) cyanobacterial–algal reef crest. The patch reefs expand upwards from an initial bioherm phase with a small base to a laterally extensive biostrome phase. This gives them a thumb-tack appearance. In stage 2 of the bioherm phase, rigid framework development and high reef relief resulted in breakage of angular blocks up to 15 m long, which were incorporated into the reefs or fell into adjacent sediments. Poorly sorted talus haloes (Millingsklint Member) also developed adjacent to stage 2 of the bioherm phase. These include angular blocks and exhibit depositional slopes up to 40° away from the reefs. Stage 3 biostrome development was mainly non-rigid cluster reef, which shed skeletal debris (Domkyrka Member) but few lithified blocks. Stage 4 biostrome development was a reef crest with open to closed frame structure. Storm breakage and overturning produced large blocks with complex cavity fill sequences including double geopetals. Relief during the bioherm phase, indicated by fallen blocks and talus slopes, was up to at least 15 m; during the biostrome phase, it was up to 10 m.
[11]
Rong Jiayu Wang Yi Zhan Renbin et al . On the Tongzi uplift: Evidence of northward expansion of Qianzhong Oldland during Aeronian, Llandovery, Silurian
[J]. Journal of Stratigraphy , 2012 , 36 (4 ), 679 -691 .
[本文引用: 6]
[戎嘉余 , 王怿 , 詹仁斌 ,等 . 论桐梓上升——志留纪埃隆晚期黔中古陆北扩的证据
[J]. 地层学杂志 , 2012 , 36 (4 ):679 -691 .]
URL
[本文引用: 6]
摘要
黔渝川边区16个县、25个志留系兰多维列统剖面的岩石、生物地层、沉积构造、风化壳及其黏土矿物成分证实,在石牛栏组最上部沉积期间(埃隆晚期),上扬子海域南部海水曾一度退尽并遭受风化剥蚀,使黔中古陆范围明显扩大。埃隆晚期全球海平面处于上升期,而研究区却出现海平面下降、基底隆升并露出海面,笔者将这次地表升隆事件命名为桐梓上升。它的风化剥蚀过程是短暂的(可能不超过0.1Myr),但海底升隆过程漫长(可能超过2Myr)。扬子区在兰多维列世发生的上升事件(宜昌上升、桐梓上升和扬子上升)均受到华南大地构造活动的控制,这些活动深深地影响了扬子海域古地理变迁。相对而言,桐梓上升的范围最窄、规模最小、时限最短,在区域地层对比意义较大。志留纪可能在埃隆晚期之后,上扬子海域海水从黔东北向南,先到达黔中贵阳和黔东南凯里一带,后侵漫到黔东南三都和独山地区。分析表明,志留纪华南大地构造活动并非处于匀变状态下,而具有强弱不等的幕式活动阶段,反映了华南大地构造运动对上扬子海域影响的时间点与强度。
[12]
Wang Haijun Meng Xianwu Liang Zirui et al . Organic reef characteristics and exploration orientation of Silurian Shiniulan Formation, southeastern Sichuan Basin
[J]. China Petroleum Exploration , 2016 , 21 (5 ):33 -41 .
[本文引用: 1]
[王海军 , 孟宪武 , 梁子锐 ,等 . 川东南志留系石牛栏组生物礁特征与勘探方向
[J]. 中国石油勘探 , 2016 , 21 (5 ):33 -41 .]
DOI
URL
[本文引用: 1]
摘要
通过对野外剖面、钻井资料及地震资料详细分析,明确了川东南地区志留系石牛栏组沉积期为一向北倾斜的大型缓坡,由南向北可分为内缓坡、中缓坡及外缓坡—陆棚。其中石牛栏组二段中缓坡相带发育,水体清澈,能量较高,发育礁滩相碳酸盐岩沉积。岩性主要为介壳灰岩、珊瑚灰岩及生屑灰岩等,地震上生物礁滩表现为地层增厚、“亮点”反射等特征,平面上主要沿叙永—西门1井—温水—隆盛2井呈北东向带状分布。元厚—四面山一带位于生物礁滩主体带,微古地貌较高,极可能发育礁滩相白云岩储层,是石牛栏组油气勘探有利方向。
[13]
Tan Xianfeng Li Zhijun Jiang Yanxia et al . Sedimentary characteristics of bioherm in the Lower Silurian Shiniulan Fm,southeastern Chongqing
[J]. Oil & Gas Geology , 2014 , 35 (1 ):56 -64 .
Magsci
[本文引用: 1]
[谭先锋 , 李志军 , 蒋艳霞 ,等 . 渝东南地区下志留统石牛栏组生物礁沉积特征
[J]. 石油与天然气地质 , 2014 , 35 (1 ):56 -64 .]
DOI
URL
Magsci
[本文引用: 1]
摘要
渝东南地区下志留统石牛栏组发育一定规模的生物礁。以重庆万盛机枪台剖面和贵州习水吼滩剖面为主要对象,利用野外观察、普通薄片、碳/氧同位素等手段,对研究区下志留统石牛栏组生物礁进行了详细研究。结果表明,研究区生物礁主要发育石牛栏组中上部,且具有一定规模;岩石结构及碳/氧同位素变化揭示了该时期海平面由缓慢上升到急剧下降的变化过程,沉积环境表现为早期浅水混积陆棚—中期生物礁滩沉积—晚期礁前斜坡沉积的演化过程,水体环境适合珊瑚礁及生物群落的大量繁殖。由此证实了渝东南地区下志留统石牛栏组生物礁发育主要受海平面变化及陆源碎屑物质注入的影响,海平面的缓慢变化是生物礁发育的基本条件,陆源碎屑物质的多少制约了生物礁的发育过程。
[14]
Zhou Kenken Mou Chuanlong Xu Xiaosong et al . Early Silurian paleogeography and source-reservoir-cap rocks of the Middle-Upper Yangtze region in South China
[J]. Petroleum Exploration & Development , 2014 , 41 (5 ):623 -632 .
[本文引用: 1]
[周恳恳 , 牟传龙 , 许效松 ,等 . 华南中上扬子早志留世古地理与生储盖层分布
[J]. 石油勘探与开发 , 2014 , 41 (5 ):623 -632 .]
DOI
URL
[本文引用: 1]
摘要
依据"构造控盆、盆地控相、相 控油气基本地质条件"思路,重建华南中上扬子地区早志留世鲁丹期、埃隆期和特列奇期岩相古地理格局,并从盆地沉积演化的角度总结生储盖层发育条件与时空分 布规律。赫南特期末期冰川消融后全球的快速海进和构造挤压造成的地壳挠曲,导致中上扬子地区在鲁丹期形成以深水陆棚为主的古地理格局,发育的多个坳陷区成 为有利烃源岩的沉积中心。埃隆期处于构造挤压的间歇期,由于相对海平面下降等因素,形成特殊的混积型碳酸盐缓坡,以礁滩相为主的碳酸盐岩储集层主要分布在 沿岸的潮下高能带。特列奇期构造运动加剧,盆地整体抬升。东部作为构造挤压的前锋带,形成滨岸/潮坪/三角洲组成的复合性海岸环境,有利于储集砂体的发 育,其中以小河坝组的临滨砂坝和三角洲前缘席状砂最具储集潜力,而广泛分布的韩家店组潮坪—陆棚相泥页岩可作为下伏含油气层系的良好封盖层。图9表3参 30
[15]
Wang Ruihua Tan Qinyin Fu Jianyuan et al . Sedimentary characteristics of the Silurian organic reefs from the Shiniulan Formation in southeastern Sichuan
[J]. Sedimentary Geology and Tethyan Geology , 2013 , 33 (2 ):10 -16 .
[本文引用: 1]
[王瑞华 , 谭钦银 , 付建元 ,等 . 川东南志留系石牛栏组生物礁沉积特征
[J]. 沉积与特提斯地质 , 2013 , 33 (2 ):10 -16 .]
[本文引用: 1]
[16]
Tan Xianfeng Jiang Yanxian Wang Jia et al . Paleo-ocean environmental fluctuations and their constraints on micro/nano porous of shale during Early Silurian in upper Yangtze Region, Southwest China
[J]. Journal of Nanoscience & Nanotechnology , 2017 , 17 (9 ):6 051 -6 066 .
[本文引用: 2]
[17]
He Li Tan Qinyin Wang Ruihua et al . Sedimentary facies, sedimentary model and evolution Shihniulan Formation of Early Silurian in southeastern Sichuan
[J]. Journal of Mineralogy & Petrology , 2013 , 33 (4 ):96 -106 .
[本文引用: 3]
[何利 , 谭钦银 , 王瑞华 ,等 . 川东南早志留世石牛栏期沉积相、沉积模式及其演化
[J]. 矿物岩石 , 2013 , 33 (4 ):96 -106 .]
DOI
URL
[本文引用: 3]
摘要
利用6口钻井、14条实测与观察剖面、大量岩心和200余块岩石薄片等资料,对川东南石牛栏组进行了沉积相和沉积微相研究,根据川东南地区石牛栏组沉积特点,将其分为局限台地相、台地边缘浅滩相、台地边缘生物礁相、台地边缘斜坡相、陆棚相等5个基本的相带。由于期间经历了两次二级海平面的升降过程,使石牛栏组发育台地边缘礁滩,礁滩沉积相带在纵横向上不断迁移、抬升,主要发育于台地的边缘,呈带状分布,明显受沉积相带控制。总的看来,川东南地区石牛栏期受海平面上升的影响,形成了碳酸盐斜坡和台地边缘礁滩沉积环境;但整体上海平面波动幅度不大,其沉积模式属于缓坡沉积模式。
[18]
Deng Xiaojie Wang Guan Huang Yong et al . Bank facies of the Shihniulan Formation(late Aeronian, Llandovery, Silurian) at the Shixi Section,Tongzi,Northern Guizhou
[J]. Guizhou Geology , 2016 , 33 (3 ):192 -198 .
[本文引用: 3]
[邓小杰 , 王冠 , 黄勇 ,等 . 黔北桐梓狮溪志留纪埃隆晚期石牛栏段的生物滩相
[J]. 贵州地质 , 2016 , 33 (3 ):192 -198 .]
URL
[本文引用: 3]
摘要
志留纪兰多维列世埃隆晚期,滇黔桂古陆以北的陆表海缓坡上沉积了石牛栏组石牛栏段浅水灰岩。珊瑚—层孔虫为主的后生动物格架岩建造的点礁分布北界仅延伸到桐梓水坝塘一带;更远岸的狮溪镇白芷垭剖面虽然远离古陆剥蚀区,层状灰岩中极低的粉砂质、泥质含量指示高清澈度的环境指标,内碎屑和少量生屑混合沉积的滩相灰岩沉积深度在最大浪基面之上,但珊瑚—层孔虫密度偏低,后生动物格架岩的丰度以及个体大小均达不到礁灰岩的标准,也不呈现生物礁特有的正向海底地貌。本文实例诠释了海底深度变化显著制约底栖群落的古生态分异,后生动物点礁栖居的深度范围有限。
[19]
Tan Xianfeng Li Zhijun Jiang Yanxia et al . Mixed sedimentation and constraints on reef of Shiniulan formation in lower Silurian, southeast of Chongqing
[J]. Petroleum Geology & Recovery Efficiency , 2014 , 21 (3 ):6 -9 .
[本文引用: 5]
[谭先锋 , 李志军 , 蒋艳霞 ,等 . 渝东南地区下志留统石牛栏组混合沉积作用及对生物礁发育的制约
[J]. 油气地质与采收率 , 2014 , 21 (3 ):6 -9 .]
DOI
URL
[本文引用: 5]
摘要
渝东南地区下志留统石牛栏组广 泛发育碳酸盐与陆源碎屑的混合沉积。通过野外观察、薄片鉴定、同位素分析等手段,对研究区石牛栏组混合沉积作用及对生物礁的制约进行了研究。结果表明,渝 东南地区石牛栏组主要发育泥质灰岩与砂质泥岩、泥晶灰岩与泥岩、生物灰岩与灰质泥岩3种混积层系和泥质灰岩、砂质/粉砂质灰岩、含粉砂生物礁灰岩(生物灰 岩)、灰质粉砂岩、灰质泥岩6种混积岩。沉积环境的演化过程表现为早期浅水混积陆棚—中期生物礁滩—晚期礁前斜坡及浅水混积陆棚;陆源碎屑含量从底部到顶 部总体为降低的过程,与海平面的演化规律存在一定的耦合。混合沉积主要发生于浅水混积陆棚及生物礁滩/斜坡2种沉积环境。在石牛栏组生物礁发育过程中,陆 源碎屑的注入是制约生物礁及生物群落发育的根本原因。
[20]
Deng Xiaojie The Paleoecology of Reef Complex the Shiniulan Formation (Upper Aeronian, Llandovery, Silurian) Tongzi, North Guizhou[D]
. Beijing: University of Chinese Academy of Sciences , 2012 .
[本文引用: 2]
[邓小杰 . 黔北桐梓志留纪埃隆晚期石牛栏组礁组合的古生态学[D]
. 北京:中国科学院大学 , 2012 .]
[本文引用: 2]
[21]
Zhang Tingshan Lan Guangzhi Kershaw S The control structure and the fluctuation of sea level of Sichuan Silurian reefs
[J]. Acta Petrolei Sinica , 1999 , 20 (3 ):19 -24 .
[本文引用: 2]
[张廷山 , 蓝光志 , Kershaw S 构造及海面波动对四川盆地志留纪生物礁的控制
[J]. 石油学报 , 1999 , 20 (3 ):19 -24 .]
DOI
URL
[本文引用: 2]
摘要
通过上扬子板块区各大地构造单元,四川盆地南北缘志留系锶同位素,碳氧同位素水平所显示的海平面变化及其与生物礁发育的关系分析得出:(1)志留纪时,四川盆地南北缘部分虽受上扬子板块内稳定古隆起(古陆)控制的陆源碎屑-碳酸盐岩缓坡环境,在靠近古陆的地区生物礁发育,而远离古陆时,由于水体逐渐加深,生物礁则不发育;(2)盆地南缘锶同位素从龙马溪组至石牛栏组呈波动下降,反映出海平面从龙马溪且至石牛栏组总体趋势
[22]
Zhang Tingshan Chen Xiaohui Bian Liceng et al . Tectonic control of the silurian reef distribution and development on upper Yangtze Platform
[J]. Acta Sedimentologica Sinica , 1996 , 14 (4 ):84 -93 .
Magsci
[本文引用: 1]
[张廷山 , 陈晓慧 , 边立曾 ,等 . 大地构造对上扬子区志留纪生物礁分布及发育的控制
[J]. 沉积学报 , 1996 , 14 (4 ):84 -93 .]
Magsci
[本文引用: 1]
摘要
根据上扬子板块的基底特征,及板块边缘特征等不同大地构造单元演化特征入手,分别对上扬子板块基底地貌、板块边缘构造及板内不同构造单元控制志留纪古沉积环境及生物礁发育演化进行了论述。在上扬子板块的北缘(南秦岭构造带),为被动大陆边缘,沉积环境适于生物礁发育,类型较多。而板块西缘(金沙江构造带)为主动大陆边缘,在岛弧区发育部分点礁以及生物层;板块内部受古隆起影响的地区(川西北、川东南、黔北),形成缓坡沉积环境,在浅缓坡区生物礁十分发育,类型众多;但是板内深断裂(龙门山,二郎山—攀西裂谷带)边缘生物礁不甚发育,仅见生物层—小型点礁组合。志留纪时,上扬子板块受全球海平面变化的影响明显,至Wenlock期后,大部分地区因海平面下降而无沉积。但是在板块边缘凹陷区,因区域构造的影响,全球海平面波动对其影响不大,Wenlock期后继续接受沉积且发育生物礁。
[23]
Ma Dongzhou Chen Hongde Zhu Lidong et al . Depositional system and lithofacies and paleogeography of the Silurian Shiniulan Formation in the southern Sichuan Basin, China
[J]. Journal of Chengdu University of Technology , 2006 , 33 (3 ):228 -232 .
[本文引用: 2]
[马东洲 , 陈洪德 , 朱利东 ,等 . 川南下志留统石牛栏组沉积体系与岩相古地理
[J]. 成都理工大学学报:自然科学版 , 2006 , 33 (3 ):228 -232 .]
DOI
URL
[本文引用: 2]
摘要
四川盆地南部下志留统石牛栏组为在缓坡陆棚基础上发育起来的碳酸盐台地的沉积,可划分出4个岩性段和2个沉积旋回.沉积体系在空间展布特征上表现为由南向北的从碎屑滨岸→局限台地→开阔台地→生物礁/滩→台地边缘斜坡→浅水陆棚浅滩→泥质浅水陆棚→深水陆棚变化,在垂向上则由泥/灰质深水→浅水陆棚→碳酸盐台地的变化;海平面变化是控制沉积体系发育和空间分布的重要因素.
[24]
Zhu Zhijun Chen Hongde An analysis of sedimentary characteristics and model of Silurian Xiaoheba Formation in southeastern Sichuan Province
[J]. Geology in China , 2012 , 39 (1 ):64 -76 .
[本文引用: 2]
[朱志军 , 陈洪德 . 川东南地区早志留世晚期沉积特征及沉积模式分析
[J]. 中国地质 , 2012 , 39 (1 ):64 -76 .]
DOI
URL
[本文引用: 2]
摘要
通过对川东南地区野外露头、大量岩心、测井等资料的综合研究结果表明,研究区早志留世晚期发育2个沉积体系组,包括3种沉积相及众多亚相、微相类型。其中三角洲沉积主要发育于靠近江南—雪峰山隆起的川东南—湘西地区;碳酸盐岩台地沉积主要发育于川南—黔北地区;陆棚沉积主要发育于宜宾—重庆一带。沉积模式显示了早志留世晚期川东南地区为受古隆起控制的局限海盆,研究区东部发育一套以碎屑岩为主的小河坝组,由于江南—雪峰山不断隆起,向研究区提供了大量物源,沉积了一套向上变粗的三角洲沉积。研究区南部受黔中隆起控制,由于黔中隆起相对稳定,在其北侧主要发育了与小河坝组同时异相的石牛栏组碳酸盐岩台地沉积。该新认识有助于了解川东南地区小河坝组砂体的时空展布,以及预测该区有利油气聚集区带。对下一步油气勘探部署具有重要的理论意义和实际指导价值。
[25]
Zhou Dazhi. Sequence Stratigraphy Sedimentary Facies and Reservoir Characteristics of Shiniulan Formation in Southeast Area of Sichuan Basin[D]
. Chengdu: Chengdu University of Technology , 2010 .
[本文引用: 1]
[周大志 . 川东南地区石牛栏组层序地层、沉积相及储层特征研究[D]
. 成都:成都理工大学 , 2010 .]
[本文引用: 1]
[26]
Lan Guangzhi Zhang Tingshan Gao Weidong Classification, genesis and significance of nodular limestone of Early Silurian in NW Sichuan
[J]. Journal of Southwest —China Petroleum Institute 1994 , 16 (3 ):1 -5 .
[本文引用: 1]
[蓝光志 , 张廷山 , 高卫东 . 川西北地区早志留世瘤状灰岩的类型、成因及意义
[J]. 西南石油学院学报 , 1994 , 16 (3 ):1 -5 .]
DOI
URL
[本文引用: 1]
摘要
根据本区“瘤状灰岩”的分布、成分及特征,作者将其划分为A、E、C三种类型。A、B型的成因与蓝绿藻的活动有密切关系,与生物礁、丘一起,沉积于浅缓坡位置.C型瘤状灰岩的成因与压溶作用有关,处于深缓坡位置。本区瘤状灰岩具有一定的含油气性.
[27]
Wu Jinwei Xia Shufang Notes on the origin of the “polygonal marking” limestones
[J].Journal of Nanjing University , 1989 , 25 (1 ):136 -141 .
[本文引用: 1]
[吴劲薇 , 夏树芳 . 关于“龟裂纹灰岩”成因的探讨
[J]. 南京大学学报 , 1989 , 25 (1 ):136 -141 .]
URL
[本文引用: 1]
[28]
Alberstadt L P Walker K R Zurawski R Patch reefs in the Middle Ordovician (Carters Limestone) in Tennessee and vertical zonation in Ordovician reefs
[J].Geological Society of America.Bulletin , 1974 , 3 (85 ): 429 -441 .
[本文引用: 1]
[29]
Wang Jianpo Li Yue Zhang Yuanyuan et al . Early-middle Ordovician Calathium reef mounds:History and palaeoecology
[J]. Acta Palaeontologica Sinica , 2011 , 50 (1 ): 132 -140 .
[本文引用: 3]
[王建坡 , 李越 , 张园园 ,等 . 早—中奥陶世瓶筐石礁丘:历史和古生态学
[J]. 古生物学报 , 2011 , 50 (1 ):132 -140 .]
URL
[本文引用: 3]
摘要
瓶筐石(Calathium)是早-中奥陶世期间常见的重要的造礁生物之一,与菌藻类、海绵以不等丰度相聚集共同组成位于浅海区的礁丘群落。早期阶段(特马道克期-弗洛期早期)以瓶筐石-海绵-菌藻类礁群落为主,其后因海绵丰度减弱而逐渐演替为瓶筐石-菌藻群落,达瑞威尔期以瓶筐石为群落主体的礁丘分布已趋于局限,中奥陶世晚期伴随着珊瑚-层孔虫造礁群落的崛起,瓶筐石礁丘骤然衰减。瓶筐石在礁丘建造过程中主要是障积生屑,因而有利于形成海底正向地貌隆起,所形成点礁的生长阶段和群落的更替过程易于识别。一般分为定殖期生屑滩型的礁基,至拓殖期和泛殖期的礁核部位则多为瓶筐石和/或海绵构成的障积岩和菌藻类形成的粘结岩,与礁间相和/或礁翼相界线多显现清晰边界。经过一定距离搬运的瓶筐石也可与其他生屑颗粒共同堆积成展布宽泛的生物滩。
[30]
Li Qiufen Miao Shunde Jiang Qingchun et al . Sedimentary characteristics and reef-forming model of Changxing Formation in Panlongdong Section of Xuanhan,Sichuan
[J]. Journal of Jilin University (Earth Science Edition )2015 , 45 (5 ):1 322 -1 331 .
[本文引用: 1]
[李秋芬 , 苗顺德 , 江青春 ,等 . 四川宣汉盘龙洞长兴组生物礁沉积特征及成礁模式
[J]. 吉林大学学报:地球科学版 , 2015 , 45 (5 ):1 322 -1 331 .]
DOI
URL
[本文引用: 1]
摘要
结合野外露头剖面观察与深入剖析,对宣汉盘龙洞长兴组生物礁沉积特征和成礁模式进行了室内综合分析。该礁位于鄂西-城口海槽西侧台缘带,为相对海平面整体下降背景下形成的一套加积-进积型台地边缘生物礁。该露头出露良好,长兴组以海绵骨架礁灰岩和海绵骨架礁白云岩为主。综合岩性、古生物等沉积相标志,该区长兴组属于碳酸盐岩台地边缘沉积体系,可进一步划分出斜坡相、台地边缘生物礁和台地边缘生屑滩相,其中台地边缘生物礁又可划分为礁基、礁核、礁盖、礁前、礁后等亚相。区域沉积环境、岩性、古生物以及生物礁的内部构成分析认为,盘龙洞生物礁可分为3期成礁旋回:第一旋回礁是低能环境下形成的未经充分生长而被淹死的礁,主要由生屑泥晶灰岩和少量骨架岩组成;第二旋回礁形成于较高能环境下,经历了充分生长后暴露而死亡,暴露时间短,仅顶部发生了较弱的白云岩化;第三旋回礁则形成于高能环境下,经历了充分生长并暴露出海平面后死亡,其死亡后经波浪和水流的改造作用而形成生屑滩,长期暴露发生强烈白云岩化。
[31]
Wang Yanqi Hu Mingyi Liu Fuyan et al . Rock types and evolution of reef of Changxing Formation in Jiantianba of western Hubei
[J]. Lithologic Reservoirs , 2008 , 20 (3 ):44 -48 .
Magsci
[本文引用: 1]
[王延奇 , 胡明毅 , 刘富艳 ,等 . 鄂西利川见天坝长兴组海绵礁岩石类型及礁体演化阶段
[J]. 岩性油气藏 , 2008 , 20 (3 ):44 -48 .]
DOI
URL
Magsci
[本文引用: 1]
摘要
<p>鄂西利川见天坝长兴组生物礁位于镶边型台地陡斜的边缘上,濒临鄂西海槽!属于台地边缘</br>礁。 该礁体的最大特点是大型柱状海绵丰富,构成生物骨架&。生物礁岩石类型主要有障积岩、障积-骨</br>架岩、骨架岩、粘结岩、角砾状灰岩%5种。通过分析见天坝生物礁的生物类型、岩石宏观类型和特征!,推</br>测生物礁的发展演化经历了%5个阶段:奠基期、拓殖期、泛殖期、衰亡期和礁盖期。</p>
[32]
Wu Xichun Wang Quanfeng Chen Sizhong et al . Considering controls on development and distribution of reef reservoirs in South China Sea from the hydrocarbon accumulation potential of Tertiary reefs in the world
[J]. China Offshore Oil & Gas , 2011 , 23 (4 ):218 -224 .
[本文引用: 1]
[吴熙纯 , 王权锋 , 陈斯忠 ,等 . 从世界第三纪生物礁的油气储集潜能看中国南海生物礁储层发育和分布的控制因素
[J]. 中国海上油气 , 2011 , 23 (4 ):218 -224 .]
DOI
URL
[本文引用: 1]
摘要
在对世界第三纪生物礁的油气储集潜能、东南亚第三纪生物礁的主要成岩作用与油气储层形成机理以及中国南海第三纪生物礁储层发育和分布的控制因素进行分析的基础上,探讨了中国南海生物礁储层的油气勘探。世界第三纪生物礁发育的最鼎盛期是早中新世Burdigalian期至中中新世Serravalian期,油气主要产于晚渐新世至中中新世的生物礁储层中,以真礁的油气储集潜能为最好。东南亚第三纪生物礁一滩组合的主要成岩作用类型是近地表古岩溶、白垩化和白云化,分别形成古岩溶储层、白垩化储层及白云化储层。中国南海第三纪生物礁储层的发育和分布主要受控于洋流和海平面变化、地裂运动、礁的类型、礁的成岩相分区、礁的就地转移性和礁的沉积水深等因素,在深水区晚渐新世至中中新世地层中都有寻找礁型储层的前景,勘探时须将生物礁一滩组合作整体部署。
[33]
Wang Rui Yu Kefu Wang Yinghui et al . The diagenesis of coral reefs
[J]. Advances in Earth Science , 2017 , 32 (3 ):221 -233 .
[本文引用: 1]
[王瑞 , 余克服 , 王英辉 ,等 . 珊瑚礁的成岩作用
[J]. 地球科学进展 , 2017 , 32 (3 ):221 -233 .]
[本文引用: 1]
[34]
Wei Xi Zhu Yongjun Yin Jihong et al . Constrains and growing trend of biological reef in South China Sea Basin
[J]. Special Oil and Gas Reservoirs , 2006 , 13 (1 ):10 -15 .
[本文引用: 2]
[魏喜 , 祝永军 , 尹继红 ,等 . 南海盆地生物礁形成条件及发育趋势
[J]. 特种油气藏 , 2006 , 13 (1 ):10 -15 .]
DOI
URL
[本文引用: 2]
摘要
生物礁由生物生长堆积而成,其形成条件与造礁生物的生存环境相关,主要包括海水的温度、盐度和溶解氧的含量;海水深度、透光度和海平面的升降变化;季风、海流、热带气旋和太阳辐射。南海处于赤道以北的低纬度地区,具备生物礁形成的海水和气候等基本条件,生物礁的形成和发育趋势主要受南海盆地形成演化制约。研究表明,南海盆地生物礁形成具有南早北晚,东早西晚的发育规律,平面上北、西、南陆架和陆坡是生物礁形成的有利地区。
[35]
Qi Wentong. Evolution of Reef Ecosystem and Global Environmental Change History [M].Beijing :Peking University Press , 2002 : 9-39, 71-84, 101-139, 141-142 , 144 -148 .
[本文引用: 1]
[齐文同 . 生物礁生态系统演化和全球环境变化历史 [M]. 北京 :北京大学出版社, 2002:9-39, 71-84, 101-139, 141- 142 , 144 -148 .]
[本文引用: 1]
[36]
Yuan Fuli Effects of glacial and interglacial epoches on quaternary strata and lithologic characters
[J]. Earth Science —Journal of China University of Geosciences 1993 ,18 (6 ):686 -698 .
[本文引用: 1]
[袁复礼 . 冰期与间冰期在第四纪地层与岩性上的作用
[J]. 地球科学——中国地质大学学报 , 1993 ,18 (6 ):686 -698 .
[本文引用: 1]
[37]
Kershaw S Quaternary reefs of Northeastern Sicily: Structure and growth controls in an unstable tectonic setting
[J]. Journal of Coastal Research , 2000 , 16 (4 ):1 037 -1 062 .
[本文引用: 1]
[38]
Wu Yasheng Fan Jiasong Quantitative evaluation of the sea-level drop at the End-Permian: Based on reefs
[J]. Acta Geologica Sinica 2003 , 77 (1 ):95 -102 .
DOI
URL
[本文引用: 1]
摘要
姝 The amplitude of pre-Quaternary sea level drop, H, can be calculated by using the formula H = D + To, where To is the original thickness from the top of the tidal deposits on the reef core to the bottom of the tidal deposits on the reef front, or to the bottom of the ancient meteoric vadose zone, or to the edge of the mixed-water dolostone zone. The identity and similarity between the sea-level drop amplitudes calculated from different reefs far away from each other indicates that such sea-level changes are eustatic rather than relative changes. Evidence of an end-Permian sea-level drop has been found on the Changxingian (i.e. the end of the Palaeofusulina zone) reefs at Ziyun in South China, including algal laminated deposits, sabkha-related dolostone, desiccation cracks, dissolution collapse breccia. According to calculation based on the meteoric dissolution zone of the reef-core sequence at Ziyun, Guizhou province, the amplitude of the sea-level drop at the end-Permian is about 89.3 m. Calculation vi
[39]
Yan Zhaobin Guo Fusheng Pan Jiayong et al . Application of C, O and Sr isotopic compositions of carbonates in Paleoclimate and paleo oceanic environment
[J]. Contributions to Geology & Mineral Resources Research , 2005 , 20 (1 ):53 -56 .
[本文引用: 1]
[严兆彬 , 郭福生 , 潘家永 ,等 . 碳酸盐岩C,O,Sr同位素组成在古气候、古海洋环境研究中的应用
[J]. 地质找矿论丛 , 2005 , 20 (1 ):53 -56 .]
[本文引用: 1]
[40]
Melchin M J Holmden C Carbon isotope chemostratigraphy of the Llandovery in Arctic Canada: Implications for global correlation and sea-level change
[J]. GFF , 2006 , 128 (2 ):173 -180 .
DOI
URL
[本文引用: 1]
[41]
Chen He Gong Enpu Guan Changqing et al . Application of constitution of carbon, oxygen and strontium isotopes and trace elements in study of paleoenvironment of reefs
[J]. Global Geology , 2008 , 27 (2 ):130 -136 .
[本文引用: 3]
[陈鹤 , 巩恩普 , 关长庆 ,等 . C、O、Sr同位素及微量元素组成在生物礁礁体环境研究中的应用
[J]. 世界地质 , 2008 , 27 (2 ):130 -136 .]
[本文引用: 3]
[42]
Chen Guowei Basic characteristics of the formation of bioherm and reef oil-gas pools in the South China Sea
[J]. Marine Geology Lettiers , 2003 , 19 (8 ):32 -37 .
[本文引用: 1]
[陈国威 . 南海生物礁及礁油气藏形成的基本特征
[J]. 海洋地质前沿 , 2003 , 19 (8 ):32 -37 .]
DOI
URL
[本文引用: 1]
摘要
南海是我国生物礁最发育的海区,在地质历史上,中新世已发现生物礁有百余个,第四纪生物礁有118个.生物礁类型有塔礁、补丁礁、块礁、台地边缘礁、环礁.其中中新世发育的各种类型的生物礁很多成为重要的油气聚集的场所,如曾母盆地的L礁油藏、珠江口盆地的流花11-1礁油藏.进一步研究南海生物礁形成的特征与成藏的条件,为今后南海油气勘探提供依据.
[43]
Zhang Xiulian Relationship between of carbon and oxygen stable isotope in carbonate rocks and paleosalinity and paleotemperature of seawater
[J]. Acta Sedimentologica Sinica , 1985 , 3 (4 ):17 -30 .
Magsci
[本文引用: 1]
[张秀莲 . 碳酸盐岩中氧、碳稳定同位素与古盐度、古水温的关系
[J]. 沉积学报 , 1985 , 3 (4 ):17 -30 .]
Magsci
[本文引用: 1]
摘要
一、概论 近年来,岩相古地理研究的意义已渐增大,进展甚速,其中用δ<sup>13</sup>C、δ<sup>18</sup>O值确定古盐度、古水温的方法引起了国内外学者的重视。事实证明,在一定条件下,这是恢复古环境的有效标志。
[44]
Shao Longyi The velation of theoxygen and carbon isotope in the carbonate roeks to the Paleotem Perature etc
[J]. Journal of China University of Mining & Technology , 1994 ,23 (1 ):39 -45 .
[本文引用: 1]
[邵龙义 . 碳酸盐岩氧、碳同位素与古温度等的关系
[J]. 中国矿业大学学报 , 1994 , 23 (1 ):39 -45 .]
[本文引用: 1]
[45]
Keith M L Weber J N Isotopic composition and environmental classification of selected limestones and fossils
[J]. Geochimica Cosmochimiea Acta , 1964 , 28 (10/11 ): 1 787 -1 816 .
DOI
URL
[本文引用: 2]
摘要
Average carbon and oxygen isotopic compositions of freshwater limestones show less regular changes with time of formation. Carbon isotope ratios are significantly different from those of marine limestones in sample groups as old as Carboniferous or Devonian, whereas oxygen isotope ratios differ in marine and freshwater sample groups as old as Jurassic. A graphic plot of 未C 12 versus 未O 13 gives environmental separation of more than 80 per cent of the samples, and the percentage of correct diagnosis is considerably improved in Jurassic and younger samples. It is concluded that carbon and oxygen isotopic compositions of limestones and calcareous fossils provide significant but not infallible criteria of depositional environment for samples of a wide range of geologic age. The difference in C 12 content between marine and fresh-water carbonates is attributed mainly to a variable contribution of C 13 -deficient carbon from land plants and humus, which is added to freshwater systems in several stages, but has a relatively minor influence in most marine environments.
[46]
Erez J Luz B Experimental paleotemperature equation for planktonic foraminifera
[J]. Geochimica et Cosmochimica Acta , 1983 , 47 (6 ):1 025 -1 031 .
DOI
URL
[本文引用: 1]
摘要
Small live individuals of Globigerinoides sacculifer which were cultured in the laboratory reached maturity and produced garnets. Fifty to ninety percent of their skeleton weight was deposited under controlled water temperature (14° to 30°C) and water isotopic composition, and a correction was made to account for the isotopic composition of the original skeleton using control groups. Comparison of. the actual growth temperatures with the calculated temperature based on paleotemperature equations for inorganic CaCO 3 indicate that the foraminifera precipitate their CaCO 3 in isotopic equilibrium. Comparison with equations developed for biogenic calcite give a similarly good fit. Linear regression with Craig's (1965) equation yields: t = 610.07 + 1.01 t 08 (r= 0.95) where t is the actual growth temperature and t 08 Is the calculated paleotemperature. The intercept and the slope of this linear equation show that the familiar paleotemperature equation developed originally for mollusca carbonate, is equally applicable for the planktonic foraminifer G. sacculifer. Second order regression of the culture temperature and the delta difference ( δ 18 Oc 61 δ 18 Ow) yield a correlation coefficient of r = 0.95: t 08 = 17.0 61 4.52(δ 18Oc 61 δ 18Ow) + 0.03(δ 18Oc 61 δ 18Ow) 2 t 08, δ 18Oc and δ 18 Ow are the estimated temperature, the isotopic composition of the shell carbonate and the sea water respectively. A possible cause for nonequilibnum isotopic compositions reported earlier for living planktonic foraminifera is the improper combustion of the organic matter.
[47]
Stewart D R M Pearson P N Ditchfield P W et al .Miocene tropical Indian Ocean temperatures: Evidence from three exceptionally preserved foraminiferal assemblages from Tanzania
[J]. Journal of African Earth Sciences , 2004 , 40 (3/4 ):173 -189 .
DOI
URL
[本文引用: 1]
[48]
Liu Gang Zhou Dongsheng Application of microelements analysis in identifying sedimentary environment—Taking Qianjiang formation in the Jianghan Basin as an example
[J]. Petroleum Geology & Experiment , 2007 , 29 (3 ): 307 -306 .
Magsci
[本文引用: 1]
[刘刚 , 周东升 . 微量元素分析在判别沉积环境中的应用——以江汉盆地潜江组为例
[J]. 石油实验地质 , 2007 , 29 (3 ):307 -310 .]
DOI
URL
Magsci
[本文引用: 1]
摘要
<p>湖泊沉积岩中的粘土矿物、泥岩和膏岩中的微量元素组成对沉积环境的水文变化很敏感,可以有效揭示湖平面的变化。封闭盐湖的沉积记录在垂向上以显著的岩性变化为特征,它们取决于河水补给与蒸发、沉淀和渗透之间的平衡作用,并导致湖平面的相应变化和湖岸线的频繁迁移。文中选取江汉盆地潜江组中对沉积环境反映最为敏感的硼(B)、锶(Sr)、钡(Ba)、钛(Ti)、铁(Fe)、磷(P)、锰(Mn)等微量元素指标,分析其在不同层序、不同体系域的变化特征,并讨论其环境意义。沉积岩中B含量一般比岩浆岩高,在页岩中其含量可达135×10<sup>-6</sup>,砂岩中较低,一般为(1~40)×10<sup>-6</sup>。Sr元素在咸水中含量一般为0.8‰~1‰,在淡水中的含量一般为0.1‰~0.3‰。</p>
[49]
Feng Hongzhen Yu Jianhua Fang Yiting et al. The analysis of salinity in Wufengian age of the Upper Yangtze sea region
[J]. Journal of Stratigraphy , 1993 , 17 (3 ):179 -185 .
[本文引用: 1]
[冯洪真 , 俞剑华 , 方一亭 ,等 . 五峰期上扬子海古盐度分析
[J]. 地层学杂志 , 1993 , 17 (3 ):179 -185 .]
[本文引用: 1]
[50]
Wang Jianpo Li Yue Cheng Long et al . Paleozoic reefs and their paleogeological controls in south China block
[J]. Acta Palaeontologica Sinica , 2014 , 53 (1 ): 121 -131 .
[本文引用: 2]
[王建坡 , 李越 , 程龙 ,等 . 华南板块古生代生物礁及其古地理控制因素
[J]. 古生物学报 , 2014 , 53 (1 ):121 -131 .]
[本文引用: 2]
[51]
Pan Yongxin Wang Xueqin Mud-belts serve as Criteria of sandstone in Palaeotidalites
[J]. Shanxi Mining Institute Learned Journal , 1991 , 9 (2 ):157 -161 .
[潘永信 , 王学勤 . 泥质条带作为古代潮汐砂岩的判据
[J]. 山西矿业学院学报 , 1991 , 9 (2 ):157 -161 .]
URL
[52]
Johnson M E Baarli B G Nestor H et al . Eustatic sea-level patterns from the Lower Silurian (Llandovery Series) of southern Norway and Estonia
[J]. Geological Society of America Bulletin , 1991 , 103 (3 ):315 -335 .
DOI
URL
[本文引用: 1]
[53]
Johnson M E Tracking silurian eustasy: Alignment of empirical evidence or pursuit of deductive reasoning?
[J]. Palaeogeography Palaeoclimatology Palaeoecology , 2010 , 296 (4 ):276 -284 .
DOI
URL
[本文引用: 1]
摘要
Sea level is not static, but liable to fluctuations due to addition or subtraction of water in the world's oceans, as well as changes to the shape and holding capacity of ocean basins. Relative changes in sea level are well supported by the rock record on a regional scale. Whether or not global (eustatic) changes are evident and how frequently they occurred during any given interval of time is a matter of contention among stratigraphers. Opinions have evolved over the last century with arguments based on refinements in biostratigraphy, chemostratigraphy, radiometric dating, and conceptual advances in sequence stratigraphy derived from technological advances in seismic stratigraphy. The Pulsation Theory of A.W. Grabau (1936) attributed to Paleozoic strata a global history of 11 highstands distributed through a sequence with 21 subdivisions. In 1977, Peter Vail and associates from the Exxon Production Research Company independently interpreted a similar Paleozoic history showing 10 second-order highstands but distributed over 19 subdivisions. Vail's approach was model-based and followed a deductive path, while Grabau's was based on inductive reasoning. Recent refinements in a Paleozoic sea-level curve by Haq and Schutter (2008) are based on the same deductive approach taken by the Vail group, but pinned to patterns in sequence stratigraphy. Drawing on the Silurian System as a Paleozoic sample, the timing, frequency, and magnitude of sea-level highstands deduced by Haq and Schutter are compared with those promulgated by the author from the mid-1980s onward using empirical evidence more in line with Grabau's methodology. Both apply the concept of geographic reference areas, but Haq and Schutter identify 50% more Silurian highstands over an interval lasting 27.7 million. years. Eight out of 10 Silurian highstands identified by this author match or overlap 8 out of 15 highstands recognized by Haq and Schutter. At issue is which, if any, qualify as eustatic signals with re
[54]
Haq B U Schutter S R A chronology of Paleozoic sea-level changes
[J]. Science , 2008 , 322 (5 898 ):64 -68 .
DOI
URL
PMID
[本文引用: 1]
摘要
Sea levels have been determined for most of the Paleozoic Era (542 to 251 million years ago), but an integrated history of sea levels has remained unrealized. We reconstructed a history of sea-level fluctuations for the entire Paleozoic by using stratigraphic sections from pericratonic and cratonic basins. Evaluation of the timing and amplitude of individual sea-level events reveals that the magnitude of change is the most problematic to estimate accurately. The long-term sea level shows a gradual rise through the Cambrian, reaching a zenith in the Late Ordovician, then a short-lived but prominent withdrawal in response to Hirnantian glaciation. Subsequent but decreasingly substantial eustatic highs occurred in the mid-Silurian, near the Middle/Late Devonian boundary, and in the latest Carboniferous. Eustatic lows are recorded in the early Devonian, near the Mississippian/Pennsylvanian boundary, and in the Late Permian. One hundred and seventy-two eustatic events are documented for the Paleozoic, varying in magnitude from a few tens of meters to 鈭125 meters.
[55]
Alroy J Geographical, environmental and intrinsic biotic controls on Phanerozoic marine diversification
[J]. Palaeontology , 2010 , 53 (6 ):1 211 -1 235 .
DOI
URL
[本文引用: 1]
摘要
Abstract: The Paleobiology Database now includes enough data on fossil collections to produce useful time series of geographical and environmental variables in addition to a robust global Phanerozoic marine diversity curve. The curve is produced by a new 090004shareholder quorum090005 method of sampling standardization that removes biases but avoids overcompensating for them by imposing entirely uniform data quotas. It involves drawing fossil collections until the taxa that have been sampled at least once (the 090004shareholders090005) have a summed total of frequencies (i.e. coverage) that meets a target (the 090004quorum090005). Coverage of each interval090005s entire data set is estimated prior to subsampling using a variant of a standard index, Good090005s u. This variant employs counts of occurrences of taxa described in only one publication instead of taxa found in only one collection. Each taxon090005s frequency within an interval is multiplied by the interval090005s index value, which limits the maximum possible sampling level and thereby creates the need for subsampling. Analyses focus on a global diversity curve and curves for northern, southern and 090004tropical090005 (3000°N to 3000°S) palaeolatitudinal belts. Tropical genus richness is remarkably static, so most large shifts in the curve reflect trends at higher latitudes. Changes in diversity are analysed as a function of standing diversity; the number, spacing and palaeolatitudinal position of sampled geographical cells; the mean onshore090009offshore position of cells; and proportions of cells from carbonate, onshore and reefal environments. Redundancy among the variables is eliminated by performing a principal components analysis of each data set and using the axis scores in multiple regressions. The key factors are standing diversity and the dominance of onshore environments such as reefs. These factors combine to produce logistic growth patterns with slowly changing equilibrium values. There is no evidence of unregulated exponential growth across any long stretch of the Phanerozoic, and in particular there was no large Cenozoic radiation beyond the Eocene. The end-Ordovician, Permo090009Triassic and Cretaceous090009Palaeogene mass extinctions had relatively short-term albeit severe effects. However, reef collapse was involved in these events and also may have caused large, longer term global diversity decreases in the mid-Devonian and across the Triassic/Jurassic boundary. Conversely, the expansion of reef ecosystems may explain newly recognized major radiations in the mid-Permian and mid-Jurassic. Reef ecosystems are particularly vulnerable to current environmental disturbances such as ocean acidification, and their decimation might prolong the recovery from today090005s mass extinction by millions or even tens of millions of years.
[56]
Nestor H Silurian
[M]∥Raukas A , Teedumäe A,eds. Geology and Mineral Resources of Estonia. Tallinn: Estonian Academy Publishers , 1997 :89 -106 .
[本文引用: 1]
[57]
Cole S R Haynes J T Lucas P C et al . Faunal and sedimentological analysis of a latest Silurian stromatoporoid biostrome from the central Appalachian Basin
[J]. Facies , 2015 , 61 (3 ): 2 -16 .
DOI
URL
[本文引用: 1]
[58]
Rhebergen F Munnecke A Jarochowska E First report of (Porifera) from the Wenlock of Gotland, Sweden
[J]. Geologiska Fãreningen I Stockholm Fãrhandlingar , 2016 , 138 (3 ):1 -6 .
[本文引用: 1]
[59]
Antoshkina A I Soja C M Late Silurian reconstruction indicated by migration of reef biota between Alaska, Baltica (Urals), and Siberia (Salair)
[J]. GFF , 2006 , 128 (2 ):75 -78 .
DOI
URL
[本文引用: 1]
[60]
Freitas T A D Nowlan G S A new, major Silurian reef tract and overview of regional Silurian reef development, Canadian Arctic and north Greenland
[J]. Bulletin of Canadian Petroleum Geology , 1998 , 46 (3 ):327 -349 .
[本文引用: 1]
[61]
Antoshkina A I Organic buildups and reefs on the Palaeozoic carbonate platform margin, Pechora Urals, Russia
[J]. Sedimentary Geology , 1998 , 118 (1/4 ):187 -211 .DOI:10.2110/pec.02.72 .
URL
[本文引用: 2]
摘要
In the Pechora Urals, Palaeozoic organic structures are widespread and are represented by various types, from small bioherms to large barrier reefs which can be observed throughout Upper Ordovician–Lower Permian strata. In the pre-orogenic history of Palaeozoic carbonate sedimentation and reef growth three stages are distinguished. Each stage is characterized by a distinct reef geometry, growth rate and depositional setting. (1) The reefs of the lower Palaeozoic (Ashgillian–lower Emsian) stage were restricted to the outer shelf margin, and they were wave-resistant stromatolite-rich frameworks, except for upper Llandoverian knoll and patch reefs. The total thickness of the lower Palaeozoic reefs is 60–1200 m. (2) The middle Palaeozoic (middle Frasnian–lower Tournaisian) organic buildups are quiet-water carbonate bank-type structures generally between 300 and 700 m thick. They were formed on both the outer and the inner shelf margin; however, they are better developed and more characteristic of the latter setting. Stromatoporoid–microbial and microbial associations form the main buildup facies separated by bedded interbuildup sediments. The upper Palaeozoic (upper Visean–Sakmarian) organic structures are represented by both isolated wave-resistant shelf margin reefs, up to 200 m thick, and quiet-water carbonate banks, up to 330 m thick. These buildups were located in the outer and inner shelf margin. Framework-builders were mainly branching and platy stromatoporoids associated with stromatolites in the reefs, and calcified microbes and phylloid algae in the carbonate banks. The distribution and geometry of the buildups was mainly determined by regional tectonic events caused by the passive-to-active margin transition of the northeastern European Platform. The size of the reefs depended on the magnitude and duration of sea-level fluctuations.
[62]
Golonka J Plate-tectonic maps of the Phanerozoic
[M]∥Kieesling W, Flügel E, Golonka J, eds. Phanerozoic Reef Patterns , 2013 :1 772 -1 781 .DOI:10.2110/pec.02.72 .
[63]
Zadoroshnaya N M Nikitin I F Kazakhstanskaya skladcataya oblast
[M]∥Belenitskaya G A, Zadoroshaya N M, eds. Rifogennye i sulfatonosnye formatsii Fanerozaoya SSSR(Phanerozoic Reefal and Sulfate bearing Formations of the USSR). Ministerstvo Geologii SSSR, Moscow, Nedra , 1990 : 41 -52 .
[本文引用: 2]
[64]
Schneider K A Ausich W I Paleoecology of framebuilders in Early Silurian reefs (Brassfield Formation, Southwestern Ohio)
[J]. Palaios , 2002 , 17 (3 ):237 -248 .
DOI
URL
[本文引用: 1]
[65]
Suchy D R Stearn C W Lower Silurian sequence stratigraphy and sea-level history of the Hudson Bay Platform
[J]. Bulletin of Canadian Petroleum Geology , 1992 , 40 (4 ):335 -355 .
DOI
URL
[本文引用: 1]
摘要
Trends in relative sea-level change, shown by curves drawn from drill core and outcrop sections, were used to correlate individual parasequences among widely separated localities in Llandoverian carbonates of the Hudson Bay Platform. Correlations show that two sequences, bounded by regional disconformities, are present: (1) the Severn River Formation and (2) the Ekwan River and Attawapiskat formations, and possibly part of the Kenogami River Formation (listed in ascending order). Initial onlap of marine facies at the base of the Severn River Formation progressed from north to south, occurring first in the Hudson Bay Basin then in the Moose River Basin; final retreat of the seas at the end of Attawapiskat time was in the opposite direction. The Moose River Basin was always shallower than the Hudson Bay Basin, but both basins were remarkably shallow and flat. The most extensive inundations occurred during Ekwan River and Attawapiskat depositional times. For the remainder of the Silurian following Attawapiskat time, only supratidal facies or subaerial exposure surfaces are recorded on the platform, indicating that relative sea level was very low at that time. Correlations based on matching trends in relative sea-level change show that strata of the upper Ekwan River Formation in the southern Moose River Basin are time equivalents of the Attawapiskat reefs in the northern part of the basin and that reefal outcrops along the Attawapiskat River represent one interval of reef growth. Large-scale trends in the relative sea-level curves delineate four major Early Silurian sea-level highstands (late Rhuddanian, late Aeronian, early Telychian and late Telychian) also recognized in other basins across North America.
[66]
Munneckea Axel Männikb Peep New biostratigraphic and chemostratigraphic data from the Chicotte Formation (Llandovery, Anticosti Island, Laurentia) compared with the Viki core (Estonia, Baltica)
[J]. Estonian Journal of Earth Sciences , 2009 , 58 (3 ):159 -169 .
DOI
URL
[本文引用: 2]
[67]
Brunton F R Copper P Paleoecologic, temporal, and spatial analysis of early Silurian Reefs of the Chicotte Formation, Anticosti Island, Quebec, Canada
[J]. Facies , 1994 , 31 (1 ):57 -79 .
DOI
URL
[本文引用: 1]
[68]
Li Qijian Wang Yuanyuan Li Yue et al. Embryonic patchy reefs from the silurian of guizhou—An example of muddy sediments constraining reef-bank growth
[J]. Acta Palaeontologica Sinica , 2012 , 51 (1 ): 127 -136 .
[本文引用: 1]
[李启剑 , 王媛媛 , 李越 ,等 . 泥质对志留系礁滩生长的抑制作用:黔北桐梓韩家店组的例证
[J]. 古生物学报 , 2012 ,51 (1 ):127 -136 .]
URL
[本文引用: 1]
摘要
黔北桐梓的戴家沟剖面和狮溪剖面志留系兰多维列统特列奇阶下部的韩家店组出露完好,该组泥岩、粉砂岩中夹有厚度1—3m、直径4—7m的小型点礁。礁核相多具典型的障积格架岩特征,但生长时限短暂,群落分异度低,仅见床板珊瑚、单体四射珊瑚、苔藓虫和海百合茎,礁间为珊瑚、苔藓虫和海百合茎碎片堆积的滩相,伴生丰富的遗迹化石。在陆源碎屑快速沉积的背景下,浑浊海水频繁的富营养化过程限制了礁体纵横向生长和朝高分异度群落发展的可能性。
[69]
Li Yue Wang Jianpo Zhang Yuanyuan et al . Carbonates on Ordovician-Silurian transition of South China and its paleoclimate meaning
[J]. Progress in Natural Science , 2008 , 18 (11 ):1 264 -1 270 .
[本文引用: 1]
[李越 , 王建坡 , 张园园 ,等 . 华南奥陶—志留纪之交的碳酸盐岩对古气候的诠释
[J]. 自然科学进展 , 2008 , 18 (11 ):1 264 -1 270 .]
DOI
URL
[本文引用: 1]
摘要
华南板块晚奥陶世到早志留世冰川事件前后所记录的碳酸盐岩,其地理分布和成因依时而变.文中所讨论的5个时间片段实例包括:晚奥陶世凯迪晚期(全球暖期)华夏古陆北缘浙赣台地上的珊瑚-层孔海绵-菌藻类礁组合;凯迪末期(冰川初期)扬子地台西缘川西汉源康滇古陆边缘的隐藻-钙藻灰泥丘;奥陶纪末赫南特期(主冰期)广布于上扬子区的观音桥层灰岩分远岸冷水型和近岸局部暖水型;志留纪初鲁丹早期(冰川消融)灰岩在黔中古陆边缘首现,之后鲁丹晚期向生物层复苏.这些例证显现出华南近岸区的碳酸盐岩建造特征对奥陶-志留纪之交主冰期前和主冰期后全球暖期的响应更加明显.在主冰期,古陆对来自高纬度地区凉水团具阻障作用,暖/凉水型碳酸盐岩生产力和沉积相分异显著,海水降温主要发生于远岸区,而滇黔桂古陆的近岸区仍然局部地维持着暖水沉积;主冰期后的海进也许是快速的,但要回暖到适宜造礁的程度则是缓慢的.
[70]
Munnecke A Männik P New biostratigraphic and chemostratigraphic data from the Chicotte Formation (Llandovery, Anticosti Island, Laurentia) compared with the Viki core (Estonia, Baltica)
[J]. Estonian Journal of Earth Sciences , 2009 , 58 (3 ):159 -169 .
DOI
URL
[本文引用: 1]
Control of carbonate sedimentation and reef growth in llandovery Sequences on the Northwestern Margin of the Yangtze Platform, South China
2
2004
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
Silurian and Devonian reefs: 80 million years of global greenhouse between two ice ages
1
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
Late Ordovician-Early Silurian facies development and environmental changes in the Subpolar Urals
1
2008
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
Early Silurian (Aeronian) East Point Coral patch reefs of anticosti island, Eastern Canada: First reef recovery from the ordovician/silurian mass extinction in Eastern Laurentia
9
2012
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... [4 ,5 ,8 ],珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
... [4 ,5 ,6 ,57 ].中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
... 中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段.与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] .在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
... [4 ,64 ,65 ];在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
... 加拿大东部安蒂科斯蒂岛的Menier组(埃隆晚期,兰多维列世)地层发现了生物礁,其造礁生物主要是四射珊瑚与横板珊瑚,以及较少的层孔虫,附礁生物包括海百合、腕足类、绿藻、蓝藻等[4 ] ,因其造礁生物与地层岩性等都与石牛栏组生物礁保持高度的相似性,我们认为其相当于同期的石牛栏组生物礁.通过分析可知,两地的生物礁存在几个方面的共同点:①生物礁均生长在低纬度热带大陆架的边缘地带,拥有良好的气候和适宜的海水深度;②造礁生物均为珊瑚和层孔虫,附礁生物为藻类、腕足类、海百合等,生物类型体现出高度相似性.但实际上,Menier组与石牛栏组生物礁在特征上存在很大差异性,主要体现在(图8 ):①Menier组生物礁的珊瑚、层孔虫、钙化藻呈现最大的多样性,但腕足动物、头足类和海百合的多样性却很低;石牛栏组珊瑚—层孔虫虽然大量繁殖,但整体生物多样性却较低,两地生物的种属也大相径庭.②Menier组大多数生物礁露头呈现不规则圆形,个体之间相对密集,生物礁个体或聚集的群体在露头上展布方向都是随机的,规模较大;而石牛栏组生物礁的生长模式以点礁为主,含少量生物丘和生物层的类型,规模较小.③Menier组生物礁的生物外壳、生物骨骼及层间沉积物等主要是由粉砂质灰岩至砂质灰岩构成,泥岩成分几乎没有,生物骨骼几乎完整,只在一些较浅的水域受风暴影响出现生物碎屑.而石牛栏组生物礁生物碎屑在下段地层中显示很多,生物外壳、生物骨骼及层间沉积物除一些砂质灰岩之外,还会有泥质成分.④安蒂科斯蒂岛特里奇期延续Menier组珊瑚—层孔虫礁生长趋势,在上部地层Chicotte组发育了更大规模且生物多样性更高的生物礁[66 ,67 ] ;而上扬子地台石牛栏组发育一定规模的生物礁后,特里奇期没有相应规模的生物礁发育或仅发育规模数米的小型点礁[68 ] . ...
... 加拿大Menier组与上扬子地区石牛栏组生物礁对比[4 ] ...
... Comparison of the Menier Formation and the Shiniulan Formation in the Upper Yangtze Region[4 ] ...
Global reef recovery after the end-Ordovician extinction: Evidence from late Aeronian coral-stromatoporoid reefs in South China
4
2014
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... ,5 ,8 ],珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
... 中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段.与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] .在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
Palaeoecology of corals and stromatoporoids in a Late Silurian biostrome in estonia
3
2016
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... [6 ,9 ,10 ].这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
Gypsum evaporites in a patch reef of the upper Slite Group in the Silurian (Wenlock) of Gotland, Sweden
2
2014
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
Paleobiology, ecology, and distribution of stromatoporoid faunas in biostromes of the Mid-Ludlow of gotland, Sweden
2
2008
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... [8 ,9 ].而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
New evidence of an early Pridoli barrier reef in the southern part of the Baltic Silurian basin based on three-dimensional seismic survey, Lithuania
2
2015
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
... ,9 ,10 ].这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
Growth of rigid high-relief patch reefs, Mid-Silurian, Gotland, Sweden
1
2010
... 全球生物礁生态系统在奥陶纪末期遭受破坏,直至早志留世气温回暖,全球大部分地区才开始形成一定规模生物礁[1 ,2 ] .通过前人研究发现,全球志留纪生物礁,其分布时代为埃隆期和特里奇期(早志留世)、申伍德期和侯默期(中志留世)及勒德洛和普里道利期(晚志留世),主要发育于北美大陆、扬子地台、中亚及波罗的海盆地地层中[3 ,4 ,5 ,6 ,7 ] .前人在加拿大东部安蒂科斯蒂岛埃隆阶Menier组地层、美国俄亥俄西南部及安大略省发现大规模珊瑚—层孔虫礁[4 ,5 ,8 ] ,珊瑚—层孔虫礁成为全球志留纪生物礁系统的主要造礁形式.其中,早志留世造礁生物主要以珊瑚—层孔虫类型为主,中—晚志留世,层孔虫、微生物、藻类等生物大量发育,参与生物礁主要造礁环节[8 ,9 ] .而在欧洲板块波罗的海盆地中,志留纪生物礁的恢复相对于全球其他地区稍晚一些,如瑞典哥特兰岛和爱沙尼亚境内直至中志留世文洛克时期才发现大型珊瑚—层孔虫礁;在欧洲东北部地区到晚志留世Pridoli期才发现大型层孔虫礁[6 ,9 ,10 ] .这些研究表明,全球生物礁在志留纪的发育较好,分布范围广泛,造礁生物多以珊瑚和层孔虫为主. ...
论桐梓上升——志留纪埃隆晚期黔中古陆北扩的证据
6
2012
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... [11 ],生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 上扬子区志留纪生物礁的演化与所处区域的构造环境与构造运动相关,自埃隆期后,该区域地层长期暴露于水体以外,制约生物礁在志留纪发展与演化,直至二叠纪再次被水体淹没,生物礁才得以发育与演化.研究区石牛栏组的岩性、生物礁群落、沉积及其碳氧同位素含量等证实,在石牛栏组沉积期间,黔中古陆以北和东北部的上扬子海域平缓海底发生过短暂但大范围的隆升,使海水退却遭遇风化剥蚀[11 ] . ...
... 前人研究表明,早志留世鲁丹期(龙马溪组)上扬子地台为较深水体环境,在埃隆晚期,研究区所处古大陆上升,古陆北缘逐渐向北扩展,海水的升降受其影响而变化,影响着石牛栏组生物礁的生长与演化.伴随着华夏古陆剥蚀量的增加,细碎屑物快速堆积使以往相对平静、以灰泥沉积为主的海底环境(石牛栏组上部)彻底丧失,广泛发育泥质海底,变为时而富氧、时而贫氧的沉积环境[11 ] .在这种环境下,生物群落生长发育受到影响,生物间歇性大量死亡,整个石牛栏组生物礁系统遭受破坏.从全球范围看,埃隆晚期被大多数学者认为是全球海平面的上升期[52 ,53 ,54 ] ,若对应中国南方生物层,大致埃隆晚期海水达到最高位.如此看来,上扬子地区埃隆晚期海平面也应处于上升状态中,但实际情况却与之相反,该时期整个区域海水呈现大幅度变浅的现象.这可能有4个方面原因[11 ] :①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
... [11 ]:①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
论桐梓上升——志留纪埃隆晚期黔中古陆北扩的证据
6
2012
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... [11 ],生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 上扬子区志留纪生物礁的演化与所处区域的构造环境与构造运动相关,自埃隆期后,该区域地层长期暴露于水体以外,制约生物礁在志留纪发展与演化,直至二叠纪再次被水体淹没,生物礁才得以发育与演化.研究区石牛栏组的岩性、生物礁群落、沉积及其碳氧同位素含量等证实,在石牛栏组沉积期间,黔中古陆以北和东北部的上扬子海域平缓海底发生过短暂但大范围的隆升,使海水退却遭遇风化剥蚀[11 ] . ...
... 前人研究表明,早志留世鲁丹期(龙马溪组)上扬子地台为较深水体环境,在埃隆晚期,研究区所处古大陆上升,古陆北缘逐渐向北扩展,海水的升降受其影响而变化,影响着石牛栏组生物礁的生长与演化.伴随着华夏古陆剥蚀量的增加,细碎屑物快速堆积使以往相对平静、以灰泥沉积为主的海底环境(石牛栏组上部)彻底丧失,广泛发育泥质海底,变为时而富氧、时而贫氧的沉积环境[11 ] .在这种环境下,生物群落生长发育受到影响,生物间歇性大量死亡,整个石牛栏组生物礁系统遭受破坏.从全球范围看,埃隆晚期被大多数学者认为是全球海平面的上升期[52 ,53 ,54 ] ,若对应中国南方生物层,大致埃隆晚期海水达到最高位.如此看来,上扬子地区埃隆晚期海平面也应处于上升状态中,但实际情况却与之相反,该时期整个区域海水呈现大幅度变浅的现象.这可能有4个方面原因[11 ] :①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
... [11 ]:①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
川东南志留系石牛栏组生物礁特征与勘探方向
1
2016
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
川东南志留系石牛栏组生物礁特征与勘探方向
1
2016
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
渝东南地区下志留统石牛栏组生物礁沉积特征
1
2014
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
渝东南地区下志留统石牛栏组生物礁沉积特征
1
2014
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
华南中上扬子早志留世古地理与生储盖层分布
1
2014
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
华南中上扬子早志留世古地理与生储盖层分布
1
2014
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
川东南志留系石牛栏组生物礁沉积特征
1
2013
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
川东南志留系石牛栏组生物礁沉积特征
1
2013
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
Paleo-ocean environmental fluctuations and their constraints on micro/nano porous of shale during Early Silurian in upper Yangtze Region, Southwest China
2
2017
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
川东南早志留世石牛栏期沉积相、沉积模式及其演化
3
2013
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 区域地质背景及地层分布图[17 ,24 ] ;(c) 石牛栏组岩性柱状图(重庆万盛机枪台剖面) ...
... Regional geological background and stratigraphic distribution[17 ,24 ] ; (c) The Shiniulan Formation lithology (the Jiqiangtai section of Wansheng, Chongqing) ...
川东南早志留世石牛栏期沉积相、沉积模式及其演化
3
2013
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 区域地质背景及地层分布图[17 ,24 ] ;(c) 石牛栏组岩性柱状图(重庆万盛机枪台剖面) ...
... Regional geological background and stratigraphic distribution[17 ,24 ] ; (c) The Shiniulan Formation lithology (the Jiqiangtai section of Wansheng, Chongqing) ...
黔北桐梓狮溪志留纪埃隆晚期石牛栏段的生物滩相
3
2016
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... [18 ],陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
黔北桐梓狮溪志留纪埃隆晚期石牛栏段的生物滩相
3
2016
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... [18 ],陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
渝东南地区下志留统石牛栏组混合沉积作用及对生物礁发育的制约
5
2014
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] .石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... [19 ],镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... [19 ].碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
渝东南地区下志留统石牛栏组混合沉积作用及对生物礁发育的制约
5
2014
... 早志留世龙马溪组沉积期,上扬子海洋为安静和缺氧的浅海陆棚环境,发育大量笔石动物群,其他类型的生物鲜有发育;埃隆晚期黔中古陆的上升和扩展使沉积环境变为滨浅海相沉积,温暖的气候条件下生物多样性增加,在碳酸盐台地上开始发育一定规模的生物礁[11 ] ,生物礁的类型主要为点礁[12 ,13 ,14 ] ,其造礁生物以复体四射珊瑚、横板珊瑚和层孔虫为主,大体沿四川盆地碳酸盐台地边缘缓坡地带分布,主要位于黔北和渝南一带的埃隆晚期石牛栏组中,特别是石牛栏组上部的灰岩中[15 ,16 ,17 ,18 ] ,华南的构造运动促使扬子地区上升,海水大幅度变浅对生物群落发育造成影响[11 ] ,生物礁的发育受到大地构造及海平面升降的影响[18 ] ,陆源碎屑注入也是制约生物礁演化的因素之一[19 ] .目前的研究成果表明,上扬子地区早志留世生物礁研究尚未对早志留世生物礁的发育演化做系统论述,特别是全球同期生物礁演化的差异性对比方面尚未见报道.本文通过大量野外地质考察,系统分析上扬子海洋早志留世生物礁的发育过程,并开展全球同期生物礁的差异性对比,为上扬子地区早志留世生物礁油气勘探提供借鉴,也为志留纪生物礁研究提供新的实例. ...
... 前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] .石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... [19 ],镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... [19 ].碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
黔北桐梓志留纪埃隆晚期石牛栏组礁组合的古生态学[D]
2
2012
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
黔北桐梓志留纪埃隆晚期石牛栏组礁组合的古生态学[D]
2
2012
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
构造及海面波动对四川盆地志留纪生物礁的控制
2
1999
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
构造及海面波动对四川盆地志留纪生物礁的控制
2
1999
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
大地构造对上扬子区志留纪生物礁分布及发育的控制
1
1996
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
大地构造对上扬子区志留纪生物礁分布及发育的控制
1
1996
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
川南下志留统石牛栏组沉积体系与岩相古地理
2
2006
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
... 石牛栏组上覆地层为韩家店组(S1-2 hj ),岩性以黄色泥岩夹亮晶砾屑灰岩为主,含少量珊瑚、层孔虫、腕足类及头足类化石,与下伏石牛栏组呈整合接触.石牛栏组下伏地层为龙马溪组(S1 l ),沉积灰色—灰黑色的含笔石页岩夹少量泥质粉砂岩,含丰富的笔石化石及少量头足类化石[25 ] .生物礁出现在石牛栏组中上段地层中,其生物种类有珊瑚、层孔虫、腕足类和双壳类等(图1 c).自下而上石牛栏组共可分为4个段[23 ] :第一段为灰色—深灰色泥质灰岩、瘤状灰岩,夹褐黄色含钙粉砂质泥岩,含少量生物化石和虫迹,石牛栏组早期继承了龙马溪组浅水陆棚相沉积,泥质成分含量高,代表了一种相对安静的水体环境;第二段岩性以黄色及灰色粉屑灰岩和浅灰色生屑灰岩为主,夹少量粉砂泥质灰岩,含少量生物化石,层面有虫孔和虫迹,代表了一种相对动荡的沉积环境;第三段岩性为灰黑色生物礁灰岩、灰色—浅灰色生屑灰岩及生物灰岩,含大量珊瑚(蜂巢珊瑚目及链珊瑚)、层孔虫、腕足类及双壳类,上部有紫红色灰岩砾块,夹有泥质及粉砂质条带;第四段岩性为灰色砾屑灰岩及生屑灰岩,夹少量泥质条带,具小型斜层理,代表了一种相对安静的沉积环境.总体上,石牛栏组第一段至第四段显示了从一种相对低能的沉积环境向高能环境,再向低能环境转变的一个过程. ...
川南下志留统石牛栏组沉积体系与岩相古地理
2
2006
... 石牛栏组主要分布在上扬子区黔北和渝南一带,其构造位置为上扬子地台黔中隆起北缘及川中隆起以南的部分区域(图1 a),岩石类型以礁灰岩、生物碎屑灰岩、泥岩和砂岩为主.上扬子海洋在奥陶纪末期经历了生物大灭绝以后,直至志留纪气温回暖,生物才得以大量繁殖,此时上扬子海洋由南向北(黔北—渝南一带)其古地理格局呈现由近岸到远延展的陆表海碳酸盐台地缓坡带,这为石牛栏期生物礁的生长发育提供了有利的地势条件[18 ,20 ] .志留纪时期的上扬子区整体表现为陆棚—碳酸盐缓坡沉积环境,石牛栏组发育于陆表海的中缓坡地带,发育了一定规模的生物礁[21 ,22 ] .上扬子海洋自南向北依次发育滨岸、内缓坡,中缓坡、外缓坡、浅水砂质陆棚、浅水混合陆棚和深水陆棚,石牛栏组生物礁为中缓坡相沉积,主要集中在上扬子黔北—渝东南区域内.受沉积相带控制,生物礁分布范围横向上向东北延伸至重庆秀山一带,向西延伸至四川文山一带,由南向北自重庆万盛到贵州桐梓尖灭,总体呈串珠状分散在这一曲折的相带内[23 ] (图1 b).本文以上扬子地区主要是贵州省习水县吼滩剖面和重庆市万盛区机枪台剖面为例,结合习水县羊九、骑龙等剖面来综合观察和分析石牛栏组生物礁的生长发育. ...
... 石牛栏组上覆地层为韩家店组(S1-2 hj ),岩性以黄色泥岩夹亮晶砾屑灰岩为主,含少量珊瑚、层孔虫、腕足类及头足类化石,与下伏石牛栏组呈整合接触.石牛栏组下伏地层为龙马溪组(S1 l ),沉积灰色—灰黑色的含笔石页岩夹少量泥质粉砂岩,含丰富的笔石化石及少量头足类化石[25 ] .生物礁出现在石牛栏组中上段地层中,其生物种类有珊瑚、层孔虫、腕足类和双壳类等(图1 c).自下而上石牛栏组共可分为4个段[23 ] :第一段为灰色—深灰色泥质灰岩、瘤状灰岩,夹褐黄色含钙粉砂质泥岩,含少量生物化石和虫迹,石牛栏组早期继承了龙马溪组浅水陆棚相沉积,泥质成分含量高,代表了一种相对安静的水体环境;第二段岩性以黄色及灰色粉屑灰岩和浅灰色生屑灰岩为主,夹少量粉砂泥质灰岩,含少量生物化石,层面有虫孔和虫迹,代表了一种相对动荡的沉积环境;第三段岩性为灰黑色生物礁灰岩、灰色—浅灰色生屑灰岩及生物灰岩,含大量珊瑚(蜂巢珊瑚目及链珊瑚)、层孔虫、腕足类及双壳类,上部有紫红色灰岩砾块,夹有泥质及粉砂质条带;第四段岩性为灰色砾屑灰岩及生屑灰岩,夹少量泥质条带,具小型斜层理,代表了一种相对安静的沉积环境.总体上,石牛栏组第一段至第四段显示了从一种相对低能的沉积环境向高能环境,再向低能环境转变的一个过程. ...
川东南地区早志留世晚期沉积特征及沉积模式分析
2
2012
... 区域地质背景及地层分布图[17 ,24 ] ;(c) 石牛栏组岩性柱状图(重庆万盛机枪台剖面) ...
... Regional geological background and stratigraphic distribution[17 ,24 ] ; (c) The Shiniulan Formation lithology (the Jiqiangtai section of Wansheng, Chongqing) ...
川东南地区早志留世晚期沉积特征及沉积模式分析
2
2012
... 区域地质背景及地层分布图[17 ,24 ] ;(c) 石牛栏组岩性柱状图(重庆万盛机枪台剖面) ...
... Regional geological background and stratigraphic distribution[17 ,24 ] ; (c) The Shiniulan Formation lithology (the Jiqiangtai section of Wansheng, Chongqing) ...
川东南地区石牛栏组层序地层、沉积相及储层特征研究[D]
1
2010
... 石牛栏组上覆地层为韩家店组(S1-2 hj ),岩性以黄色泥岩夹亮晶砾屑灰岩为主,含少量珊瑚、层孔虫、腕足类及头足类化石,与下伏石牛栏组呈整合接触.石牛栏组下伏地层为龙马溪组(S1 l ),沉积灰色—灰黑色的含笔石页岩夹少量泥质粉砂岩,含丰富的笔石化石及少量头足类化石[25 ] .生物礁出现在石牛栏组中上段地层中,其生物种类有珊瑚、层孔虫、腕足类和双壳类等(图1 c).自下而上石牛栏组共可分为4个段[23 ] :第一段为灰色—深灰色泥质灰岩、瘤状灰岩,夹褐黄色含钙粉砂质泥岩,含少量生物化石和虫迹,石牛栏组早期继承了龙马溪组浅水陆棚相沉积,泥质成分含量高,代表了一种相对安静的水体环境;第二段岩性以黄色及灰色粉屑灰岩和浅灰色生屑灰岩为主,夹少量粉砂泥质灰岩,含少量生物化石,层面有虫孔和虫迹,代表了一种相对动荡的沉积环境;第三段岩性为灰黑色生物礁灰岩、灰色—浅灰色生屑灰岩及生物灰岩,含大量珊瑚(蜂巢珊瑚目及链珊瑚)、层孔虫、腕足类及双壳类,上部有紫红色灰岩砾块,夹有泥质及粉砂质条带;第四段岩性为灰色砾屑灰岩及生屑灰岩,夹少量泥质条带,具小型斜层理,代表了一种相对安静的沉积环境.总体上,石牛栏组第一段至第四段显示了从一种相对低能的沉积环境向高能环境,再向低能环境转变的一个过程. ...
川东南地区石牛栏组层序地层、沉积相及储层特征研究[D]
1
2010
... 石牛栏组上覆地层为韩家店组(S1-2 hj ),岩性以黄色泥岩夹亮晶砾屑灰岩为主,含少量珊瑚、层孔虫、腕足类及头足类化石,与下伏石牛栏组呈整合接触.石牛栏组下伏地层为龙马溪组(S1 l ),沉积灰色—灰黑色的含笔石页岩夹少量泥质粉砂岩,含丰富的笔石化石及少量头足类化石[25 ] .生物礁出现在石牛栏组中上段地层中,其生物种类有珊瑚、层孔虫、腕足类和双壳类等(图1 c).自下而上石牛栏组共可分为4个段[23 ] :第一段为灰色—深灰色泥质灰岩、瘤状灰岩,夹褐黄色含钙粉砂质泥岩,含少量生物化石和虫迹,石牛栏组早期继承了龙马溪组浅水陆棚相沉积,泥质成分含量高,代表了一种相对安静的水体环境;第二段岩性以黄色及灰色粉屑灰岩和浅灰色生屑灰岩为主,夹少量粉砂泥质灰岩,含少量生物化石,层面有虫孔和虫迹,代表了一种相对动荡的沉积环境;第三段岩性为灰黑色生物礁灰岩、灰色—浅灰色生屑灰岩及生物灰岩,含大量珊瑚(蜂巢珊瑚目及链珊瑚)、层孔虫、腕足类及双壳类,上部有紫红色灰岩砾块,夹有泥质及粉砂质条带;第四段岩性为灰色砾屑灰岩及生屑灰岩,夹少量泥质条带,具小型斜层理,代表了一种相对安静的沉积环境.总体上,石牛栏组第一段至第四段显示了从一种相对低能的沉积环境向高能环境,再向低能环境转变的一个过程. ...
川西北地区早志留世瘤状灰岩的类型、成因及意义
1
1994
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
川西北地区早志留世瘤状灰岩的类型、成因及意义
1
1994
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
关于“龟裂纹灰岩”成因的探讨
1
1989
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
关于“龟裂纹灰岩”成因的探讨
1
1989
... 石牛栏组生物礁在上扬子地台地区许多露头剖面中都有很好的出露,其中贵州习水羊九、重庆万盛机枪台和綦江观音桥、重庆骑龙村几个剖面生物礁出露较好.生物礁出露于石牛栏组的中上段,有的剖面可见生物造礁经历了2期旋回.其造礁生物主要为四射珊瑚、横板珊瑚(蜂巢珊瑚目和链珊瑚)与层孔虫,一些附礁生物包括苔藓虫、腕足类、藻类、海百合、双壳类及头足类等.从纵向剖面上来看,自下而上,石牛栏组泥质含量越来越少,生物种类及数量逐渐增多,其生物礁发育于石牛栏组的中上段.横向上,礁在不同剖面生物礁出露的段存在一定差异性,其中吼滩剖面相应段不发育生物礁,为生屑滩沉积,而铁索桥及羊九等剖面生物礁发育较好,分布石牛栏组的中上段(图2 ).而铁索桥与鱼化剖面生物礁灰岩出露于石牛栏组中下段,这种分布可能与当时海底地形及海水深度相关.石牛栏组下段多发育深灰色—灰色瘤状灰岩,基质为泥质,瘤体多为泥晶灰岩,这些特征指示岩石形成于相对低能静水环境中[26 ] .石牛栏组中部开始出现大量的生物活动痕迹和生物碎屑,表明水体已经开始变浅,大量的生物繁殖和生长需要充足的氧气、适宜的光照与海水深度[1 ,21 ] ,海水变浅已达生物生长最佳水体深度,石牛栏组中上段为生物礁灰岩沉积.石牛栏沉积晚期,其顶部开始出现白云岩、“龟裂纹”等浅水沉积构造,说明此时岩层及生物礁系统已开始暴露于水面之上[27 ] .这些现象表明,自下而上,石牛栏组总体表现为沉积水体由深变浅的趋势,即由浅水陆棚相沉积逐渐演化为碳酸盐缓坡相沉积,发育一定规模的生物礁. ...
Patch reefs in the Middle Ordovician (Carters Limestone) in Tennessee and vertical zonation in Ordovician reefs
1
1974
... 生物礁的生长规模从侧面上可以反映生物礁的成礁过程,一般来说,发育较好的生物礁共可分为5个成礁过程:定殖期(奠基期)、拓殖期、泛殖期、衰亡期和礁盖期(盖层期).根据石牛栏组生物礁纵向及横向发育特征,选取典型习水县羊九剖面进行分析(图5 a),大致将其成礁过程分为定殖阶段、拓殖阶段、泛殖阶段、衰亡阶段共4个生长阶段[28 ,29 ] . ...
早—中奥陶世瓶筐石礁丘:历史和古生态学
3
2011
... 生物礁的生长规模从侧面上可以反映生物礁的成礁过程,一般来说,发育较好的生物礁共可分为5个成礁过程:定殖期(奠基期)、拓殖期、泛殖期、衰亡期和礁盖期(盖层期).根据石牛栏组生物礁纵向及横向发育特征,选取典型习水县羊九剖面进行分析(图5 a),大致将其成礁过程分为定殖阶段、拓殖阶段、泛殖阶段、衰亡阶段共4个生长阶段[28 ,29 ] . ...
... 该阶段是石牛栏组生物礁的初发育时期,在生物或非生物作用下,不稳定或软质的基底变为稳定或硬质基底[29 ] ,其大体对应石牛栏组第一段.此阶段腕足类、棘皮类(如海百合)等生物用肉柄、壳刺或其他固着器官固着于不稳定或较软质的基质之上,并使基质处于稳定状态.之后,横板珊瑚及层孔虫迁移至此,共同为礁体的发育提供一个较稳定的基底.代表这个时期的典型岩石类型是泥晶灰岩、钙质泥页岩与灰质泥岩,以及一些含棘皮类、腕足类等化石的瘤状灰岩,生物种类含有少量海百合、腕足类、珊瑚等(图5 b-I). ...
... 在这个阶段块状、枝状横板珊瑚和层孔虫的发育进入鼎盛时期,大体上对应石牛栏组第三段.在菌藻类黏结岩和珊瑚及层孔虫造架岩的共同作用下,礁体具有初步的抗浪功能,逐渐形成隆起于周缘的地貌,礁体环境进一步变浅[29 ] .造礁生物和联结生物(海绵、层孔虫、钙藻等)等大量繁殖,造礁生物分异化明显增强,礁体纵横向发育到了最大规模.代表这个时期的典型岩石类型是灰色块状生物礁灰岩以及亮晶生屑灰岩,局部可见沥青和油浸.此时,生物礁的礁前、礁核与礁后环境已基本显现(图5 b-III). ...
早—中奥陶世瓶筐石礁丘:历史和古生态学
3
2011
... 生物礁的生长规模从侧面上可以反映生物礁的成礁过程,一般来说,发育较好的生物礁共可分为5个成礁过程:定殖期(奠基期)、拓殖期、泛殖期、衰亡期和礁盖期(盖层期).根据石牛栏组生物礁纵向及横向发育特征,选取典型习水县羊九剖面进行分析(图5 a),大致将其成礁过程分为定殖阶段、拓殖阶段、泛殖阶段、衰亡阶段共4个生长阶段[28 ,29 ] . ...
... 该阶段是石牛栏组生物礁的初发育时期,在生物或非生物作用下,不稳定或软质的基底变为稳定或硬质基底[29 ] ,其大体对应石牛栏组第一段.此阶段腕足类、棘皮类(如海百合)等生物用肉柄、壳刺或其他固着器官固着于不稳定或较软质的基质之上,并使基质处于稳定状态.之后,横板珊瑚及层孔虫迁移至此,共同为礁体的发育提供一个较稳定的基底.代表这个时期的典型岩石类型是泥晶灰岩、钙质泥页岩与灰质泥岩,以及一些含棘皮类、腕足类等化石的瘤状灰岩,生物种类含有少量海百合、腕足类、珊瑚等(图5 b-I). ...
... 在这个阶段块状、枝状横板珊瑚和层孔虫的发育进入鼎盛时期,大体上对应石牛栏组第三段.在菌藻类黏结岩和珊瑚及层孔虫造架岩的共同作用下,礁体具有初步的抗浪功能,逐渐形成隆起于周缘的地貌,礁体环境进一步变浅[29 ] .造礁生物和联结生物(海绵、层孔虫、钙藻等)等大量繁殖,造礁生物分异化明显增强,礁体纵横向发育到了最大规模.代表这个时期的典型岩石类型是灰色块状生物礁灰岩以及亮晶生屑灰岩,局部可见沥青和油浸.此时,生物礁的礁前、礁核与礁后环境已基本显现(图5 b-III). ...
四川宣汉盘龙洞长兴组生物礁沉积特征及成礁模式
1
2015
... 在这个阶段石牛栏组生物礁发育趋于衰亡,大体上对应石牛栏组第四段.随着造礁生物及附礁生物的繁盛生长,礁体迅速扩大,当礁体扩长达平均海平面时,形成极度浅水环境,生物礁间歇性暴露于水面[30 ] .造礁生物的生长发育受生存环境的限制,造成窄盐性造礁生物群大量死亡,剩下少数的广盐性生物(如腹足类、藻类)生存下来,此时使生物分异化变弱[31 ] .此时沉积物多带有潮坪环境特征,生物数量和种类大量减少,在纵向上礁体停止发育,顶部有少许白云石化现象.代表这个时期的典型岩生物碎屑灰岩及亮晶生屑灰岩、白云岩等(图5 b-IV). ...
四川宣汉盘龙洞长兴组生物礁沉积特征及成礁模式
1
2015
... 在这个阶段石牛栏组生物礁发育趋于衰亡,大体上对应石牛栏组第四段.随着造礁生物及附礁生物的繁盛生长,礁体迅速扩大,当礁体扩长达平均海平面时,形成极度浅水环境,生物礁间歇性暴露于水面[30 ] .造礁生物的生长发育受生存环境的限制,造成窄盐性造礁生物群大量死亡,剩下少数的广盐性生物(如腹足类、藻类)生存下来,此时使生物分异化变弱[31 ] .此时沉积物多带有潮坪环境特征,生物数量和种类大量减少,在纵向上礁体停止发育,顶部有少许白云石化现象.代表这个时期的典型岩生物碎屑灰岩及亮晶生屑灰岩、白云岩等(图5 b-IV). ...
鄂西利川见天坝长兴组海绵礁岩石类型及礁体演化阶段
1
2008
... 在这个阶段石牛栏组生物礁发育趋于衰亡,大体上对应石牛栏组第四段.随着造礁生物及附礁生物的繁盛生长,礁体迅速扩大,当礁体扩长达平均海平面时,形成极度浅水环境,生物礁间歇性暴露于水面[30 ] .造礁生物的生长发育受生存环境的限制,造成窄盐性造礁生物群大量死亡,剩下少数的广盐性生物(如腹足类、藻类)生存下来,此时使生物分异化变弱[31 ] .此时沉积物多带有潮坪环境特征,生物数量和种类大量减少,在纵向上礁体停止发育,顶部有少许白云石化现象.代表这个时期的典型岩生物碎屑灰岩及亮晶生屑灰岩、白云岩等(图5 b-IV). ...
鄂西利川见天坝长兴组海绵礁岩石类型及礁体演化阶段
1
2008
... 在这个阶段石牛栏组生物礁发育趋于衰亡,大体上对应石牛栏组第四段.随着造礁生物及附礁生物的繁盛生长,礁体迅速扩大,当礁体扩长达平均海平面时,形成极度浅水环境,生物礁间歇性暴露于水面[30 ] .造礁生物的生长发育受生存环境的限制,造成窄盐性造礁生物群大量死亡,剩下少数的广盐性生物(如腹足类、藻类)生存下来,此时使生物分异化变弱[31 ] .此时沉积物多带有潮坪环境特征,生物数量和种类大量减少,在纵向上礁体停止发育,顶部有少许白云石化现象.代表这个时期的典型岩生物碎屑灰岩及亮晶生屑灰岩、白云岩等(图5 b-IV). ...
从世界第三纪生物礁的油气储集潜能看中国南海生物礁储层发育和分布的控制因素
1
2011
... 在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关.根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响. ...
从世界第三纪生物礁的油气储集潜能看中国南海生物礁储层发育和分布的控制因素
1
2011
... 在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关.根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响. ...
珊瑚礁的成岩作用
1
2017
... 在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关.根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响. ...
珊瑚礁的成岩作用
1
2017
... 在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关.根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响. ...
南海盆地生物礁形成条件及发育趋势
2
2006
... 在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关.根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响. ...
... 前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] .石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
南海盆地生物礁形成条件及发育趋势
2
2006
... 在研究南海现代生物礁的生长发育,前人提出生物礁发育和分布与洋流和海平面变化、礁的沉积水深、礁的类型[32 ] ,成岩作用[33 ] ,海水的温度、盐度、溶氧量、透光度[34 ] 等因素密切相关.根据前人研究成果,再加上现代生物礁的生长环境,我们认为石牛栏组生物礁生长与消亡的主要受控于4个方面因素:外来物源搅动、海平面变化、海水的温度和盐度以及构造运动影响. ...
... 前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] .石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
1
2002
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
1
2002
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
冰期与间冰期在第四纪地层与岩性上的作用
1
1993
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
冰期与间冰期在第四纪地层与岩性上的作用
1
1993
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
Quaternary reefs of Northeastern Sicily: Structure and growth controls in an unstable tectonic setting
1
2000
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
Quantitative evaluation of the sea-level drop at the End-Permian: Based on reefs
1
2003
... 海平面升降是影响石牛栏组生物礁生长演化的一个重要因素.海平面不断变化影响着生物礁的外部形态及内部结构、组分特征、生长速率和堆积型式等[16 ,35 ] .海平面的变化与地球的各种地质构造运动紧密相关,包括海底地震及火山喷发,全球的冰期与间冰期更替[36 ] ,地壳升降运动[37 ] 等.海平面长期升高使海水变深,生物礁被淹没,生物窒息死亡,附礁生物藻类的光和作用减弱,从而影响礁生态系统的演化.反之,长期海平面下降则会使生物礁礁体长期暴露于水面,造礁生物大量死亡,从而抑制生物礁的生长及演化[38 ] . ...
碳酸盐岩C,O,Sr同位素组成在古气候、古海洋环境研究中的应用
1
2005
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
碳酸盐岩C,O,Sr同位素组成在古气候、古海洋环境研究中的应用
1
2005
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
Carbon isotope chemostratigraphy of the Llandovery in Arctic Canada: Implications for global correlation and sea-level change
1
2006
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
C、O、Sr同位素及微量元素组成在生物礁礁体环境研究中的应用
3
2008
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
C、O、Sr同位素及微量元素组成在生物礁礁体环境研究中的应用
3
2008
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
南海生物礁及礁油气藏形成的基本特征
1
2003
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
南海生物礁及礁油气藏形成的基本特征
1
2003
... 国外内有很多学者将碳同位素方法运用于恢复古海洋环境的研究中[39 ,40 ,41 ] .据现代海洋生物礁生长可知,其生长环境需要适宜的温度与盐度,如现代珊瑚的适应温度为18~30 ℃,盐度在28‰左右,这些因素在石牛栏组时期生物礁的生长过程中起了至关重要的作用[42 ] .为了计算生物礁发育期的海水温度和盐度,本次研究选取羊九村骑龙剖面石牛栏组51个样品进行碳氧同位素分析,挑选后期风化较弱及受成岩作用相对影响较小的样品,可以很好地反映原始沉积环境,测试由中国地质大学地质过程与矿产资源国家重点实验室MAT-251质谱仪完成. ...
碳酸盐岩中氧、碳稳定同位素与古盐度、古水温的关系
1
1985
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
碳酸盐岩中氧、碳稳定同位素与古盐度、古水温的关系
1
1985
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
碳酸盐岩氧、碳同位素与古温度等的关系
1
1994
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
碳酸盐岩氧、碳同位素与古温度等的关系
1
1994
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
Isotopic composition and environmental classification of selected limestones and fossils
2
1964
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
Experimental paleotemperature equation for planktonic foraminifera
1
1983
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
Miocene tropical Indian Ocean temperatures: Evidence from three exceptionally preserved foraminiferal assemblages from Tanzania
1
2004
... δ18 O值变化可较好地反映古海洋环境的温度变化.一般情况下,δ18 O值降低,代表温度升高;δ18 O值升高,代表温度降低[41 ] .但利用此值测定古水温有一定局限性,用δ18 O计算中生代以前的碳酸盐岩的古温度要考虑其成岩作用的影响,时代越老,其受成岩作用的影响越大[43 ] .因此,用δ18 O计算中生代以前的碳酸盐岩的古温度必须要进行“年代效应”的校正[44 ] .根据Keith等[45 ] 总结出的δ18 O与地质年代的关系,以第四纪海相碳酸盐岩的δ18 O平均值-1.2‰为标准进行“年代效应”校正,通过分析可知,石牛栏组碳酸盐岩δ18 O平均值为-9.55‰,二者差值为-8.35‰.此处采用的古温度方程式是通过校准浮游型有孔虫Globigerinoides sacculifer 古温度得出[46 ] ,碳氧同位素值均采用PDB(Pee Dee Belemnite)表示[47 ] . ...
微量元素分析在判别沉积环境中的应用——以江汉盆地潜江组为例
1
2007
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
微量元素分析在判别沉积环境中的应用——以江汉盆地潜江组为例
1
2007
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
五峰期上扬子海古盐度分析
1
1993
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
五峰期上扬子海古盐度分析
1
1993
... 古海洋的盐度含量可用m 值(m =100×MgO/Al2 O3 ),Sr/Ba值与Cl的值等指标测定,另外,综合δ18 O和δ13 C的值,也可以定量地获得古盐度的值[41 ,48 ,49 ] .本文选取Keith等[45 ] 综合δ18 O和δ13 C值的Z 值以指示古盐度的变化: ...
华南板块古生代生物礁及其古地理控制因素
2
2014
... 前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] .石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... [50 ],泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
华南板块古生代生物礁及其古地理控制因素
2
2014
... 前人在研究石牛栏组生物礁生长发育时,提出了石牛栏组广泛发育碳酸盐与陆源碎屑的混合沉积作用,制约石牛栏组生物礁生长及演化的观点[19 ,50 ] .石牛栏组宏观剖面上存在3种混积层系:泥质灰岩与灰质泥岩,生物灰岩与灰质泥岩,泥质灰岩与砂质泥岩[19 ] ,镜下可见灰岩中存在大量石英颗粒(图7 a~c).这3种岩系交互出现恰好说明石牛栏组生物礁有外来碎屑物质注入,这些物质可能来源于南东方向雪峰山隆起的陆源碎屑物[19 ] .碎屑物的注入使水体缺氧、浑浊,导致生物发生窒息死亡,光线的射入被阻挡,影响生物礁体内一些藻类的光合作用[34 ] .通过许多剖面观察,泥质条带频繁出现在石牛栏组中(图7 d),有学者认为泥质条带是潮汐沉积产物,是涨潮流带来的一定量的碎屑物及细粒悬浮物质,在潮水稳定时沉积形成[50 ] ,泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
... [50 ],泥质条带在石牛栏组中的出现也从另一方面证实了生物礁受外来物源搅动影响. ...
Eustatic sea-level patterns from the Lower Silurian (Llandovery Series) of southern Norway and Estonia
1
1991
... 前人研究表明,早志留世鲁丹期(龙马溪组)上扬子地台为较深水体环境,在埃隆晚期,研究区所处古大陆上升,古陆北缘逐渐向北扩展,海水的升降受其影响而变化,影响着石牛栏组生物礁的生长与演化.伴随着华夏古陆剥蚀量的增加,细碎屑物快速堆积使以往相对平静、以灰泥沉积为主的海底环境(石牛栏组上部)彻底丧失,广泛发育泥质海底,变为时而富氧、时而贫氧的沉积环境[11 ] .在这种环境下,生物群落生长发育受到影响,生物间歇性大量死亡,整个石牛栏组生物礁系统遭受破坏.从全球范围看,埃隆晚期被大多数学者认为是全球海平面的上升期[52 ,53 ,54 ] ,若对应中国南方生物层,大致埃隆晚期海水达到最高位.如此看来,上扬子地区埃隆晚期海平面也应处于上升状态中,但实际情况却与之相反,该时期整个区域海水呈现大幅度变浅的现象.这可能有4个方面原因[11 ] :①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
Tracking silurian eustasy: Alignment of empirical evidence or pursuit of deductive reasoning?
1
2010
... 前人研究表明,早志留世鲁丹期(龙马溪组)上扬子地台为较深水体环境,在埃隆晚期,研究区所处古大陆上升,古陆北缘逐渐向北扩展,海水的升降受其影响而变化,影响着石牛栏组生物礁的生长与演化.伴随着华夏古陆剥蚀量的增加,细碎屑物快速堆积使以往相对平静、以灰泥沉积为主的海底环境(石牛栏组上部)彻底丧失,广泛发育泥质海底,变为时而富氧、时而贫氧的沉积环境[11 ] .在这种环境下,生物群落生长发育受到影响,生物间歇性大量死亡,整个石牛栏组生物礁系统遭受破坏.从全球范围看,埃隆晚期被大多数学者认为是全球海平面的上升期[52 ,53 ,54 ] ,若对应中国南方生物层,大致埃隆晚期海水达到最高位.如此看来,上扬子地区埃隆晚期海平面也应处于上升状态中,但实际情况却与之相反,该时期整个区域海水呈现大幅度变浅的现象.这可能有4个方面原因[11 ] :①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
A chronology of Paleozoic sea-level changes
1
2008
... 前人研究表明,早志留世鲁丹期(龙马溪组)上扬子地台为较深水体环境,在埃隆晚期,研究区所处古大陆上升,古陆北缘逐渐向北扩展,海水的升降受其影响而变化,影响着石牛栏组生物礁的生长与演化.伴随着华夏古陆剥蚀量的增加,细碎屑物快速堆积使以往相对平静、以灰泥沉积为主的海底环境(石牛栏组上部)彻底丧失,广泛发育泥质海底,变为时而富氧、时而贫氧的沉积环境[11 ] .在这种环境下,生物群落生长发育受到影响,生物间歇性大量死亡,整个石牛栏组生物礁系统遭受破坏.从全球范围看,埃隆晚期被大多数学者认为是全球海平面的上升期[52 ,53 ,54 ] ,若对应中国南方生物层,大致埃隆晚期海水达到最高位.如此看来,上扬子地区埃隆晚期海平面也应处于上升状态中,但实际情况却与之相反,该时期整个区域海水呈现大幅度变浅的现象.这可能有4个方面原因[11 ] :①东侧的华夏古陆由东南向西北扩大并逐渐隆起上升;②南方黔北古陆自奥陶纪晚期逐渐隆起,并逐渐向北部扩移;③黔中古陆的隆起上升,在志留纪早期由南向北、与华夏块体一起朝扬子块体挤压;④志留纪早期的加里东运动加速了上扬子地台整体的隆起抬升.正是在这样的大地构造和古地理背景下,导致上扬子地台海平面下降,海水变浅导致生物暴露于水面大量死亡.实际上,构造运动还影响着陆源碎屑的注入、海平面的变化、海底地形、海水深度、温度及盐度等的变化,这些变化追根究底就是生物礁生长与消亡的因素.因此,志留纪兰多维列世埃隆晚期的加里东构造运动所促使的上扬子古陆的上升是生物礁生长与消亡最根本的原因. ...
Geographical, environmental and intrinsic biotic controls on Phanerozoic marine diversification
1
2010
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
Silurian
1
1997
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
Faunal and sedimentological analysis of a latest Silurian stromatoporoid biostrome from the central Appalachian Basin
1
2015
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
First report of (Porifera) from the Wenlock of Gotland, Sweden
1
2016
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
Late Silurian reconstruction indicated by migration of reef biota between Alaska, Baltica (Urals), and Siberia (Salair)
1
2006
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
A new, major Silurian reef tract and overview of regional Silurian reef development, Canadian Arctic and north Greenland
1
1998
... 由于地形和气候等各种因素的影响,全球志留纪早、中、晚期的生物礁有明显差异.生物礁在志留纪初期复苏相对缓慢,全球发现志留纪最早大型生物礁在加拿大东部的安蒂科斯蒂岛地层中[4 ] ,距离奥陶纪末大规模生物灭绝事件已有约400万年的时间[55 ] ,这也造成了生物礁在志留纪初期多样性较低.志留纪早期,全球造礁生物大多以珊瑚和层孔虫为主,其他类型生物礁鲜有发育.珊瑚—层孔虫点礁最早出现在波罗的海的兰多维列世鲁丹阶中,这是目前已知最早的珊瑚—层孔虫礁[56 ] ,但全球真正意义上形成较大规模的珊瑚—层孔虫点礁则是在早志留世埃隆期,这样的成礁模式在全球下志留统地层已得到了响应[4 ,5 ,6 ,57 ] .中—晚志留世,全球生物礁生物的多样性开始增加,除珊瑚—层孔虫礁在全球地层中的发育,其他类型的生物礁也相继出现.Rhebergen等[58 ] 在瑞典的哥特兰岛中志留世文洛克地层中发现了以海绵为主要造礁生物的点礁,海绵为造礁生物第一次出现于志留纪的地层中.晚志留世勒德洛期,在阿拉斯加东南部和乌拉尔山脉附近发现了相似度极高的以微生物—海绵—水螅为主要造礁生物的生物礁[59 ] ,以及在加拿大北极圈内发现的同时代的珊瑚—微生物黏结灰岩和微生物岩层[60 ] ,均证实志留纪生物礁的多样性增加.晚志留世勒德洛期,在瑞典哥特兰岛东南部地层中已形成大规模以层孔虫为造礁生物的生物礁,各类层孔虫组合形成一个密集的生物礁群落[7 ] ,单一造礁生物的成礁模式在晚志留世海洋中出现.总的来说,志留纪从早期到晚期各类型生物礁的大量出现,标志着志留纪生物礁多样性的增加,加之单一生物形成大型的生物礁群落,说明随着时间的推移志留纪生物礁的造礁模式由简入繁、由少至多,并逐渐成熟. ...
Organic buildups and reefs on the Palaeozoic carbonate platform margin, Pechora Urals, Russia
2
1998
... 中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段.与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] .在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
... [61 ];在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
Plate-tectonic maps of the Phanerozoic
0
2013
Kazakhstanskaya skladcataya oblast
2
1990
... 中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段.与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] .在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
... [63 ].事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
Paleoecology of framebuilders in Early Silurian reefs (Brassfield Formation, Southwestern Ohio)
1
2002
... 中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段.与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] .在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
Lower Silurian sequence stratigraphy and sea-level history of the Hudson Bay Platform
1
1992
... 中国上扬子地台,在经历了奥陶纪末期生物大灭绝后,很长时间都无生物礁发育,直至在志留纪埃隆晚期的石牛栏组中发现了生物礁,才标志着上扬子地台志留纪生物礁的生长与演化进入复苏阶段.与上扬子地台石牛栏组同时代的生物礁在全球地层均有分布,主要分布在劳伦古陆、西伯利亚古陆和哈萨克斯坦板块的地层中,这表明志留纪礁在早志留世相应地层的恢复已是一个全球性的现象[4 ,5 ,61 ~63 ] .在劳伦古陆,志留纪埃隆期生物礁分布广泛,主要分布在现今加拿大东部安蒂科斯蒂岛、安大略省北部和俄亥俄周中西部地区[4 ,64 ,65 ] ;在西伯利亚古陆,埃隆期生物礁主要分布于现今的俄罗斯乌拉尔山脉伯朝拉河附近[61 ] ;在哈萨克斯坦板块,埃隆期生物礁现今主要分布于哈萨克斯坦及乌克兰国内志留纪地层中[63 ] .事实上,全球各地志留纪埃隆期生物礁在形态、规模和演化上都存在很大差异. ...
New biostratigraphic and chemostratigraphic data from the Chicotte Formation (Llandovery, Anticosti Island, Laurentia) compared with the Viki core (Estonia, Baltica)
2
2009
... 加拿大东部安蒂科斯蒂岛的Menier组(埃隆晚期,兰多维列世)地层发现了生物礁,其造礁生物主要是四射珊瑚与横板珊瑚,以及较少的层孔虫,附礁生物包括海百合、腕足类、绿藻、蓝藻等[4 ] ,因其造礁生物与地层岩性等都与石牛栏组生物礁保持高度的相似性,我们认为其相当于同期的石牛栏组生物礁.通过分析可知,两地的生物礁存在几个方面的共同点:①生物礁均生长在低纬度热带大陆架的边缘地带,拥有良好的气候和适宜的海水深度;②造礁生物均为珊瑚和层孔虫,附礁生物为藻类、腕足类、海百合等,生物类型体现出高度相似性.但实际上,Menier组与石牛栏组生物礁在特征上存在很大差异性,主要体现在(图8 ):①Menier组生物礁的珊瑚、层孔虫、钙化藻呈现最大的多样性,但腕足动物、头足类和海百合的多样性却很低;石牛栏组珊瑚—层孔虫虽然大量繁殖,但整体生物多样性却较低,两地生物的种属也大相径庭.②Menier组大多数生物礁露头呈现不规则圆形,个体之间相对密集,生物礁个体或聚集的群体在露头上展布方向都是随机的,规模较大;而石牛栏组生物礁的生长模式以点礁为主,含少量生物丘和生物层的类型,规模较小.③Menier组生物礁的生物外壳、生物骨骼及层间沉积物等主要是由粉砂质灰岩至砂质灰岩构成,泥岩成分几乎没有,生物骨骼几乎完整,只在一些较浅的水域受风暴影响出现生物碎屑.而石牛栏组生物礁生物碎屑在下段地层中显示很多,生物外壳、生物骨骼及层间沉积物除一些砂质灰岩之外,还会有泥质成分.④安蒂科斯蒂岛特里奇期延续Menier组珊瑚—层孔虫礁生长趋势,在上部地层Chicotte组发育了更大规模且生物多样性更高的生物礁[66 ,67 ] ;而上扬子地台石牛栏组发育一定规模的生物礁后,特里奇期没有相应规模的生物礁发育或仅发育规模数米的小型点礁[68 ] . ...
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
Paleoecologic, temporal, and spatial analysis of early Silurian Reefs of the Chicotte Formation, Anticosti Island, Quebec, Canada
1
1994
... 加拿大东部安蒂科斯蒂岛的Menier组(埃隆晚期,兰多维列世)地层发现了生物礁,其造礁生物主要是四射珊瑚与横板珊瑚,以及较少的层孔虫,附礁生物包括海百合、腕足类、绿藻、蓝藻等[4 ] ,因其造礁生物与地层岩性等都与石牛栏组生物礁保持高度的相似性,我们认为其相当于同期的石牛栏组生物礁.通过分析可知,两地的生物礁存在几个方面的共同点:①生物礁均生长在低纬度热带大陆架的边缘地带,拥有良好的气候和适宜的海水深度;②造礁生物均为珊瑚和层孔虫,附礁生物为藻类、腕足类、海百合等,生物类型体现出高度相似性.但实际上,Menier组与石牛栏组生物礁在特征上存在很大差异性,主要体现在(图8 ):①Menier组生物礁的珊瑚、层孔虫、钙化藻呈现最大的多样性,但腕足动物、头足类和海百合的多样性却很低;石牛栏组珊瑚—层孔虫虽然大量繁殖,但整体生物多样性却较低,两地生物的种属也大相径庭.②Menier组大多数生物礁露头呈现不规则圆形,个体之间相对密集,生物礁个体或聚集的群体在露头上展布方向都是随机的,规模较大;而石牛栏组生物礁的生长模式以点礁为主,含少量生物丘和生物层的类型,规模较小.③Menier组生物礁的生物外壳、生物骨骼及层间沉积物等主要是由粉砂质灰岩至砂质灰岩构成,泥岩成分几乎没有,生物骨骼几乎完整,只在一些较浅的水域受风暴影响出现生物碎屑.而石牛栏组生物礁生物碎屑在下段地层中显示很多,生物外壳、生物骨骼及层间沉积物除一些砂质灰岩之外,还会有泥质成分.④安蒂科斯蒂岛特里奇期延续Menier组珊瑚—层孔虫礁生长趋势,在上部地层Chicotte组发育了更大规模且生物多样性更高的生物礁[66 ,67 ] ;而上扬子地台石牛栏组发育一定规模的生物礁后,特里奇期没有相应规模的生物礁发育或仅发育规模数米的小型点礁[68 ] . ...
泥质对志留系礁滩生长的抑制作用:黔北桐梓韩家店组的例证
1
2012
... 加拿大东部安蒂科斯蒂岛的Menier组(埃隆晚期,兰多维列世)地层发现了生物礁,其造礁生物主要是四射珊瑚与横板珊瑚,以及较少的层孔虫,附礁生物包括海百合、腕足类、绿藻、蓝藻等[4 ] ,因其造礁生物与地层岩性等都与石牛栏组生物礁保持高度的相似性,我们认为其相当于同期的石牛栏组生物礁.通过分析可知,两地的生物礁存在几个方面的共同点:①生物礁均生长在低纬度热带大陆架的边缘地带,拥有良好的气候和适宜的海水深度;②造礁生物均为珊瑚和层孔虫,附礁生物为藻类、腕足类、海百合等,生物类型体现出高度相似性.但实际上,Menier组与石牛栏组生物礁在特征上存在很大差异性,主要体现在(图8 ):①Menier组生物礁的珊瑚、层孔虫、钙化藻呈现最大的多样性,但腕足动物、头足类和海百合的多样性却很低;石牛栏组珊瑚—层孔虫虽然大量繁殖,但整体生物多样性却较低,两地生物的种属也大相径庭.②Menier组大多数生物礁露头呈现不规则圆形,个体之间相对密集,生物礁个体或聚集的群体在露头上展布方向都是随机的,规模较大;而石牛栏组生物礁的生长模式以点礁为主,含少量生物丘和生物层的类型,规模较小.③Menier组生物礁的生物外壳、生物骨骼及层间沉积物等主要是由粉砂质灰岩至砂质灰岩构成,泥岩成分几乎没有,生物骨骼几乎完整,只在一些较浅的水域受风暴影响出现生物碎屑.而石牛栏组生物礁生物碎屑在下段地层中显示很多,生物外壳、生物骨骼及层间沉积物除一些砂质灰岩之外,还会有泥质成分.④安蒂科斯蒂岛特里奇期延续Menier组珊瑚—层孔虫礁生长趋势,在上部地层Chicotte组发育了更大规模且生物多样性更高的生物礁[66 ,67 ] ;而上扬子地台石牛栏组发育一定规模的生物礁后,特里奇期没有相应规模的生物礁发育或仅发育规模数米的小型点礁[68 ] . ...
泥质对志留系礁滩生长的抑制作用:黔北桐梓韩家店组的例证
1
2012
... 加拿大东部安蒂科斯蒂岛的Menier组(埃隆晚期,兰多维列世)地层发现了生物礁,其造礁生物主要是四射珊瑚与横板珊瑚,以及较少的层孔虫,附礁生物包括海百合、腕足类、绿藻、蓝藻等[4 ] ,因其造礁生物与地层岩性等都与石牛栏组生物礁保持高度的相似性,我们认为其相当于同期的石牛栏组生物礁.通过分析可知,两地的生物礁存在几个方面的共同点:①生物礁均生长在低纬度热带大陆架的边缘地带,拥有良好的气候和适宜的海水深度;②造礁生物均为珊瑚和层孔虫,附礁生物为藻类、腕足类、海百合等,生物类型体现出高度相似性.但实际上,Menier组与石牛栏组生物礁在特征上存在很大差异性,主要体现在(图8 ):①Menier组生物礁的珊瑚、层孔虫、钙化藻呈现最大的多样性,但腕足动物、头足类和海百合的多样性却很低;石牛栏组珊瑚—层孔虫虽然大量繁殖,但整体生物多样性却较低,两地生物的种属也大相径庭.②Menier组大多数生物礁露头呈现不规则圆形,个体之间相对密集,生物礁个体或聚集的群体在露头上展布方向都是随机的,规模较大;而石牛栏组生物礁的生长模式以点礁为主,含少量生物丘和生物层的类型,规模较小.③Menier组生物礁的生物外壳、生物骨骼及层间沉积物等主要是由粉砂质灰岩至砂质灰岩构成,泥岩成分几乎没有,生物骨骼几乎完整,只在一些较浅的水域受风暴影响出现生物碎屑.而石牛栏组生物礁生物碎屑在下段地层中显示很多,生物外壳、生物骨骼及层间沉积物除一些砂质灰岩之外,还会有泥质成分.④安蒂科斯蒂岛特里奇期延续Menier组珊瑚—层孔虫礁生长趋势,在上部地层Chicotte组发育了更大规模且生物多样性更高的生物礁[66 ,67 ] ;而上扬子地台石牛栏组发育一定规模的生物礁后,特里奇期没有相应规模的生物礁发育或仅发育规模数米的小型点礁[68 ] . ...
华南奥陶—志留纪之交的碳酸盐岩对古气候的诠释
1
2008
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
华南奥陶—志留纪之交的碳酸盐岩对古气候的诠释
1
2008
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...
New biostratigraphic and chemostratigraphic data from the Chicotte Formation (Llandovery, Anticosti Island, Laurentia) compared with the Viki core (Estonia, Baltica)
1
2009
... 造成这种差异的原因是多方面的,但主要为以下4个方面:①气候变化影响.奥陶纪末期处于全球冰期,这对喜温生物的生长发育造成了一定影响.早志留世龙马溪沉积期冰川开始消融,藻类大量繁衍,并通过光合作用改善了该海区低温、缺氧的环境,成为海洋生态系统修复的主要动力.鲁丹阶早期龙马溪组灰岩较薄,生物种类少.上扬子地台直至早志留世气温才逐渐回暖,各生物门类开始复苏,受温度影响的石牛栏组生物礁是非常典型的暖水标志物.但在这期间,高纬度区的冷水团影响着扬子地台的远岸区,即使生物在局部近岸带温暖条件下幸存,但也避免不了大灭绝事件背景下生物种类,尤其是造礁动物在多样性和生物群落规模上的萧条[69 ] .②陆源碎屑的注入.Menier组生物礁在发育过程未受风暴及陆源碎屑注入的影响,但石牛栏组生物礁在发育过程中频繁受到陆源碎屑注入的影响,混合沉积的方式造成石牛栏组岩性错综复杂;动荡混浊的水体环境还对生物礁的生物多样性及生长演化造成一定程度上的限制[20 ] .③海平面的升降.Menier组生物礁受海平面波动影响较小,而埃隆期上扬子区海平面的不断变化,位于浅水区石牛栏组生物礁生长因此受到了限制[19 ] .埃隆晚期海平面持续下降,最终导致生物礁群落灭亡,生物礁在石牛栏上部志留纪地层缺失,而Menier组生物礁没有受到海平面升降的影响,在志留纪中、晚期都有很好的继承与发展[66 ,70 ] .④构造运动与海水深度.志留纪早期加里东构造运动在上扬子地区尤为强烈,从台地到盆地上扬子地台早奥陶世之后经历漫长的较深水、海底时而缺氧的环境.地形地貌的改变促使石牛栏组生物礁生长方向、规模与密度都产生了相应的变化;而上扬子区自南向北海水深度的变化同样影响着生物礁群落生物的生长与分布,从而造成上扬子地区礁的组合模式存在较大差异[11 ] .Menier组和石牛栏组生物礁不同的发育特征、演化、规模等,也恰好从侧面证实了石牛栏组其生长频受各类因素制约,导致其生长演化与Menier组生物礁呈现了较大差异性. ...

 , 谭先锋
, 谭先锋

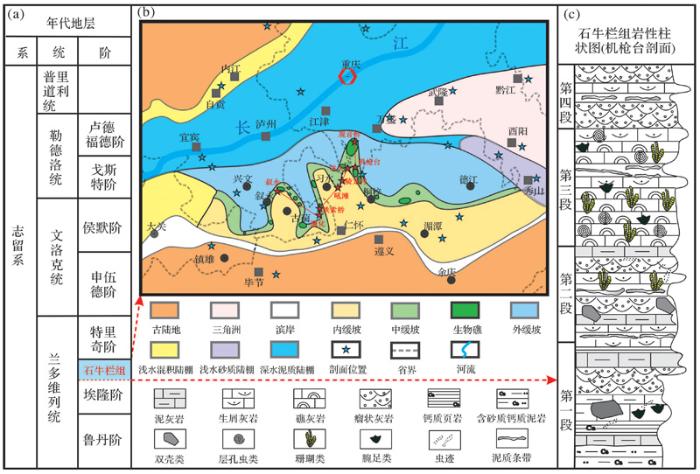
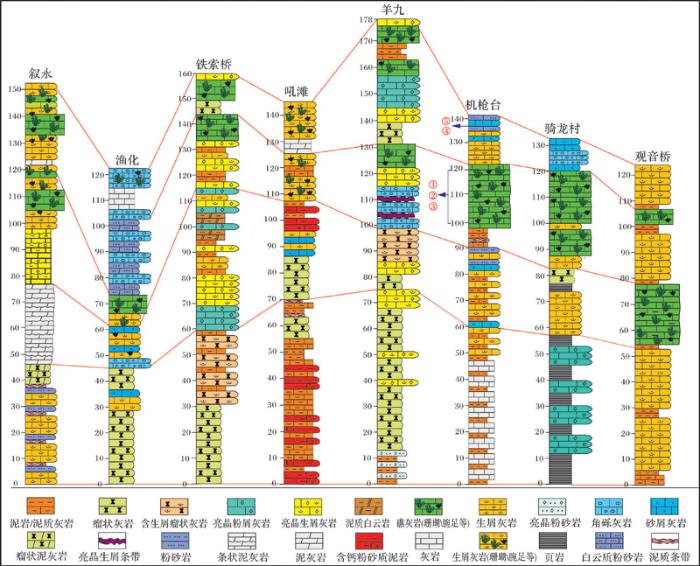

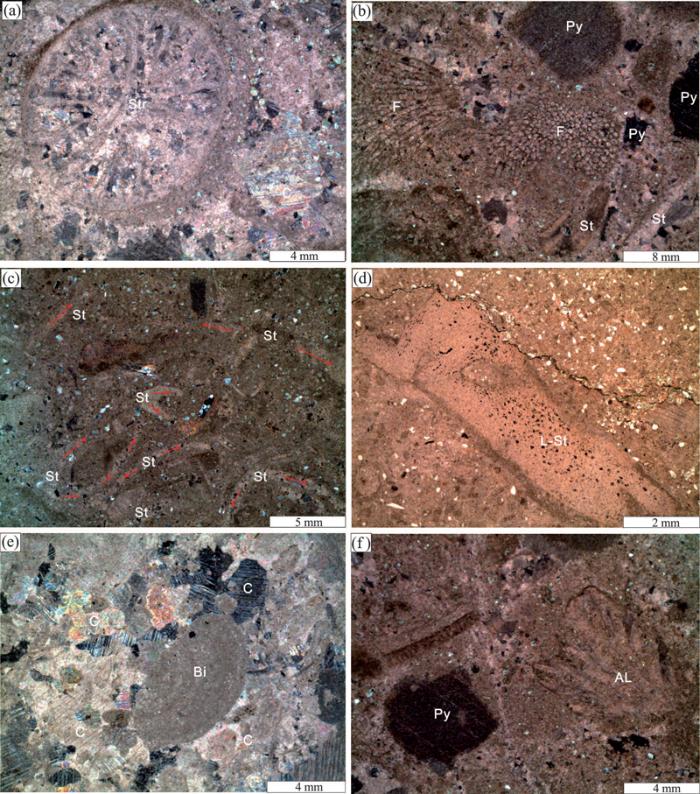
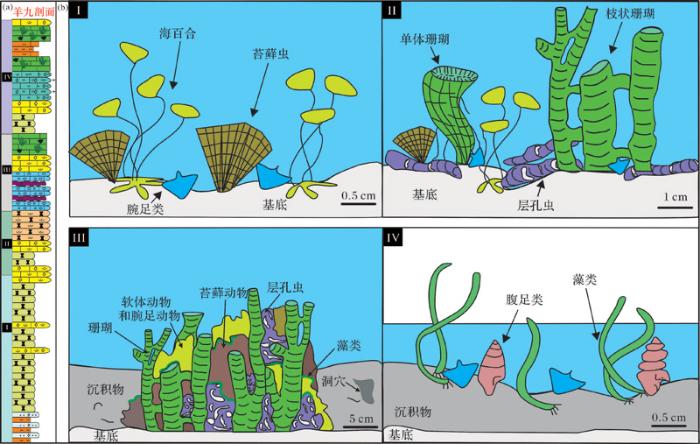
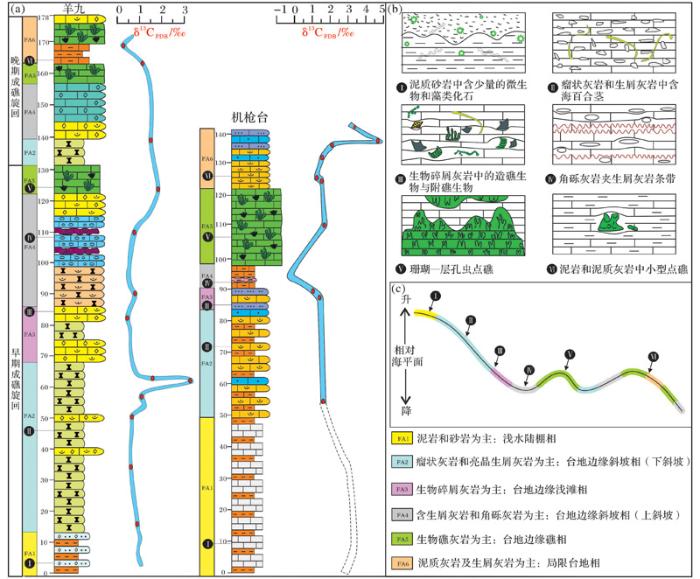

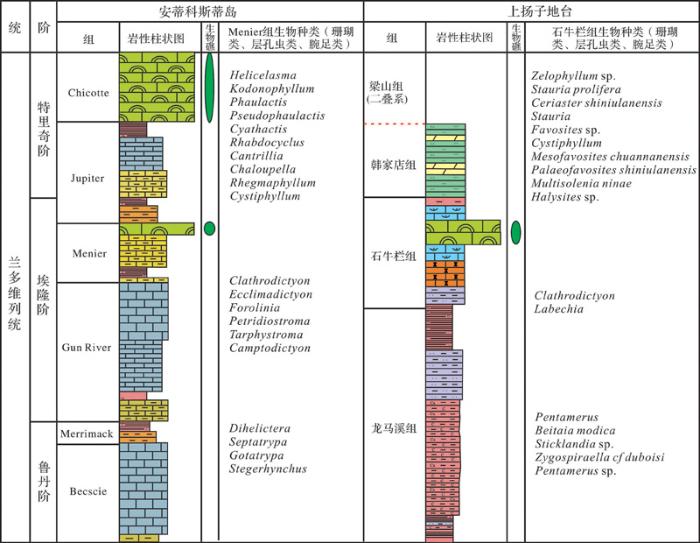
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}