...
| VPD | [8] | b1=-0.008 ...
Modelling surface conductance of pine forest
2
1988
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... Jarvis冠层导度模型有不同的形式,其中有两种比较简单的方法,一是不考虑冠层尺度通量对冠层结构(如LAI和叶倾角等)的依赖关系,这种方法会使冠层结构的效应平均到模型的率定参数中(如)[13].二是假设每个叶片对冠层的水和二氧化碳通量贡献相等,即冠层通量是叶片水平通量乘以LAI[9,33].本文采用的是第二种方法,这种简单的比例关系特别适合在大叶模型中使用,然而模型模拟结果存在的高估问题应采用修正,本文在前人研究的0.5<<1[24]的基础上发现,对于西北干旱区灌溉绿洲农田玉米而言,在农田郁闭度较高,LAI较大时0.5≤≤0.6,且本文得到的与LAI具体函数表达形式可以为模型构建中的准确取值提供必要的方法和参考. ...
Comparing the Penman-Monteith equation and a modified Jarvis-Stewart model with an artificial neural network to estimate stand-scale transpiration and canopy conductance
1
2009
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
On the scaling up leaf stomatal resistance to canopy resistance using photosynthetic photon flux density
4
2008
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... 在基于叶片气孔导度向冠层导度转换的研究中,直接用阳叶的气孔导度乘以叶面积指数(Leaf Area Index,LAI)的模型会导致对蒸散的严重高估[24],因此应将气孔导度乘以一个遮荫系数()[25].遮荫系数的意义为:通常郁闭度高的作物冠层的上半部分吸收了大部分的净辐射,冠层下方叶子被阻挡了直射光和风,因此水热传输速率比较低[26].与LAI密切相关,数值一般为0.5~1.0,随LAI的增大而减小,前人的研究中多粗略地认为为0.5,并称之为有效叶面积指数[11,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
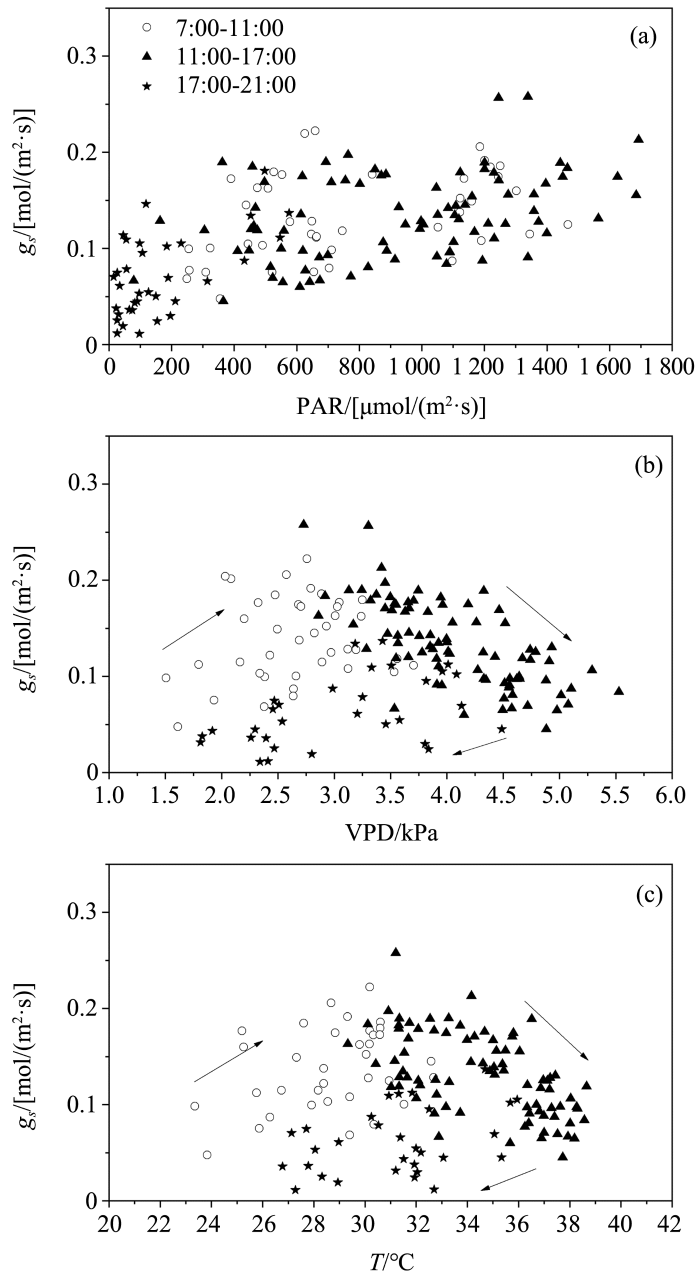
... 植物气孔导度对环境变化很敏感,在全天的不同时间段影响气孔导度的主导因子不同,其对各因素的响应关系也不同.在7:00-11:00和17:00-21:00,即气孔张开和关闭的时段,气孔导度变化主要受PAR的影响,有很多研究认为在模型化的过程中对气孔导度最重要的影响因子是PAR,Irmak等[11]提出了只考虑PAR将叶片气孔导度升尺度转换到冠层导度的方法.然而,本文发现在11:00-17:00,VPD成为气孔导度的主要限制因子,由于该时段大气湿度低,饱和差大,而水分的的运输不足以补充其蒸腾引起的水分消耗,植物会自律性地降低气孔开度[17].此外,对于相同的环境要素值,气孔导度上午和下午的值却表现出显著差别,说明气孔导度变化不只受到单一环境因素的影响,其变化是周围环境和植物自身生理调节的综合结果,因此在建模时综合考虑多个环境因子对气孔导度的影响是很有必要的. ...
Canopy reflectance, photosynthesis, and transpiration. III. A reanalysis using improved leaf models and a new canopy integration scheme
1
1992
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
A canopy conductance and photosynthesis model for use in a GCM land surface scheme
2
1998
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... Jarvis冠层导度模型有不同的形式,其中有两种比较简单的方法,一是不考虑冠层尺度通量对冠层结构(如LAI和叶倾角等)的依赖关系,这种方法会使冠层结构的效应平均到模型的率定参数中(如)[13].二是假设每个叶片对冠层的水和二氧化碳通量贡献相等,即冠层通量是叶片水平通量乘以LAI[9,33].本文采用的是第二种方法,这种简单的比例关系特别适合在大叶模型中使用,然而模型模拟结果存在的高估问题应采用修正,本文在前人研究的0.5<<1[24]的基础上发现,对于西北干旱区灌溉绿洲农田玉米而言,在农田郁闭度较高,LAI较大时0.5≤≤0.6,且本文得到的与LAI具体函数表达形式可以为模型构建中的准确取值提供必要的方法和参考. ...
Modelling the soil-plant-atmosphere continuum in a Quercus-Acer stand at Harvard Forest: The regulation of stomatal conductance by light, nitrogen and soil/plant hydraulic properties
1
1996
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
Crop water relations under different CO2 and irrigation: Testing of ecosys with the Free Air CO2 Enrichment (FACE) experiment
1
1999
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
A coupled model of stomatal conductance, photosynthesis and transpiration
1
2003
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
4
2010
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... 图1为7个典型晴天的叶片气孔导度和环境因子的平均日变化过程,表现为单峰型,上午6:30日出之后7:00-11:00气孔导度迅速增加,11:00左右到达峰值,之后缓慢下降,在17:00往后快速回落,20:30日落后在21:00降至最小.因此将日变化进程分为3个阶段(7:00-11:00、11:00-17:00和17:00-21:00).玉米叶片气孔导度对各环境变量的响应关系如图2所示,对VPD和T的响应表现出顺时针“非对称性响应”关系,即在相同环境条件下上午的气孔导度大于下午的气孔导度,气孔张开和闭合过程对环境变化的响应是非对称的,表现出滞后现象,而对PAR的响应几乎没有出现滞后现象.曲线上升阶段和下降阶段的路径不同,这对模拟日尺度的气孔行为是重要问题[17],为了提高模型的预测精度,本文进一步考虑对于叶片气孔导度日响应过程的各阶段是否应当采用不同函数形式拟合. ...
... 植物气孔导度对环境变化很敏感,在全天的不同时间段影响气孔导度的主导因子不同,其对各因素的响应关系也不同.在7:00-11:00和17:00-21:00,即气孔张开和关闭的时段,气孔导度变化主要受PAR的影响,有很多研究认为在模型化的过程中对气孔导度最重要的影响因子是PAR,Irmak等[11]提出了只考虑PAR将叶片气孔导度升尺度转换到冠层导度的方法.然而,本文发现在11:00-17:00,VPD成为气孔导度的主要限制因子,由于该时段大气湿度低,饱和差大,而水分的的运输不足以补充其蒸腾引起的水分消耗,植物会自律性地降低气孔开度[17].此外,对于相同的环境要素值,气孔导度上午和下午的值却表现出显著差别,说明气孔导度变化不只受到单一环境因素的影响,其变化是周围环境和植物自身生理调节的综合结果,因此在建模时综合考虑多个环境因子对气孔导度的影响是很有必要的. ...
... 尽管本文得到的Jarvis冠层导度模型能比较好地模拟干旱区灌溉绿洲农田玉米冠层导度的响应变化,但本质上是经验模型,对于新的植物种类和环境条件需重新对模型进行参数化,而且目前提出的气孔导度模拟模型大多为统计模型[49],还需更多理论总结进行机理的推求.本文将冠层视为“大叶”,但作物冠层结构复杂,冠层内气孔导度随叶倾角、叶位等的不同而变化,而且当冠层发育完全,叶片开始老化和变皱也会影响对冠层导度的模拟和计算[45].冠层导度是植被总体气孔导度的集合[35],随着环境因子的变化,有的气孔导度增加,有的气孔导度下降,冠层导度表示的是其整体效应[17],与气孔导度的响应变化并不完全一致,而冠层导度模型是基于叶片尺度气孔导度的提升,这也是Jarvis模型的局限性之一.然而,简化冠层结构是模型应用的必要条件[21],尽管如此,冠层导度的估算模型仍能充分反映其随主要环境影响要素的变化规律.本研究基于“单源模型”的假设,只分析了农田郁闭度较高时模型的应用效果,若在LAI较低时应用该冠层导度模型,用考虑土壤蒸发的“双源模型”模拟蒸散可能更准确,这在今后的研究需要考虑. ...
4
2010
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... 图1为7个典型晴天的叶片气孔导度和环境因子的平均日变化过程,表现为单峰型,上午6:30日出之后7:00-11:00气孔导度迅速增加,11:00左右到达峰值,之后缓慢下降,在17:00往后快速回落,20:30日落后在21:00降至最小.因此将日变化进程分为3个阶段(7:00-11:00、11:00-17:00和17:00-21:00).玉米叶片气孔导度对各环境变量的响应关系如图2所示,对VPD和T的响应表现出顺时针“非对称性响应”关系,即在相同环境条件下上午的气孔导度大于下午的气孔导度,气孔张开和闭合过程对环境变化的响应是非对称的,表现出滞后现象,而对PAR的响应几乎没有出现滞后现象.曲线上升阶段和下降阶段的路径不同,这对模拟日尺度的气孔行为是重要问题[17],为了提高模型的预测精度,本文进一步考虑对于叶片气孔导度日响应过程的各阶段是否应当采用不同函数形式拟合. ...
... 植物气孔导度对环境变化很敏感,在全天的不同时间段影响气孔导度的主导因子不同,其对各因素的响应关系也不同.在7:00-11:00和17:00-21:00,即气孔张开和关闭的时段,气孔导度变化主要受PAR的影响,有很多研究认为在模型化的过程中对气孔导度最重要的影响因子是PAR,Irmak等[11]提出了只考虑PAR将叶片气孔导度升尺度转换到冠层导度的方法.然而,本文发现在11:00-17:00,VPD成为气孔导度的主要限制因子,由于该时段大气湿度低,饱和差大,而水分的的运输不足以补充其蒸腾引起的水分消耗,植物会自律性地降低气孔开度[17].此外,对于相同的环境要素值,气孔导度上午和下午的值却表现出显著差别,说明气孔导度变化不只受到单一环境因素的影响,其变化是周围环境和植物自身生理调节的综合结果,因此在建模时综合考虑多个环境因子对气孔导度的影响是很有必要的. ...
... 尽管本文得到的Jarvis冠层导度模型能比较好地模拟干旱区灌溉绿洲农田玉米冠层导度的响应变化,但本质上是经验模型,对于新的植物种类和环境条件需重新对模型进行参数化,而且目前提出的气孔导度模拟模型大多为统计模型[49],还需更多理论总结进行机理的推求.本文将冠层视为“大叶”,但作物冠层结构复杂,冠层内气孔导度随叶倾角、叶位等的不同而变化,而且当冠层发育完全,叶片开始老化和变皱也会影响对冠层导度的模拟和计算[45].冠层导度是植被总体气孔导度的集合[35],随着环境因子的变化,有的气孔导度增加,有的气孔导度下降,冠层导度表示的是其整体效应[17],与气孔导度的响应变化并不完全一致,而冠层导度模型是基于叶片尺度气孔导度的提升,这也是Jarvis模型的局限性之一.然而,简化冠层结构是模型应用的必要条件[21],尽管如此,冠层导度的估算模型仍能充分反映其随主要环境影响要素的变化规律.本研究基于“单源模型”的假设,只分析了农田郁闭度较高时模型的应用效果,若在LAI较低时应用该冠层导度模型,用考虑土壤蒸发的“双源模型”模拟蒸散可能更准确,这在今后的研究需要考虑. ...
Responses and modeling of canopy stomatal conductance of Platycladus orientalis to environmental factors in Hedong sandy land,Ningxia
3
2018
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
... [18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
宁夏河东沙区侧柏冠层气孔导度对环境因子的响应及其模拟
3
2018
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
... [18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
Simulations of Larix principis-rupprechtii stand mean canopy stomatal conductance and its responses to environmental factors
1
2011
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
华北落叶松冠层平均气孔导度模拟及其对环境因子的响应
1
2011
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
Simulation of stomatal behaviour and transpiration of single leaf of spring wheat
1
1992
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
春小麦单叶气孔行为及蒸腾作用的模拟
1
1992
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
Estimating sensible and latent heat fluxes from a temperate broad-leaved forest using the Simple Biosphere (SiB) model
4
1997
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
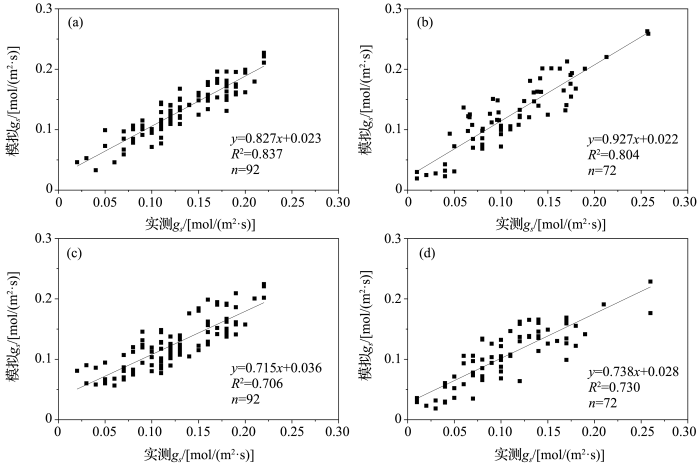
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... b2=0.208 | 0.092 |
| [21] | b1=-0.395 ...
... 尽管本文得到的Jarvis冠层导度模型能比较好地模拟干旱区灌溉绿洲农田玉米冠层导度的响应变化,但本质上是经验模型,对于新的植物种类和环境条件需重新对模型进行参数化,而且目前提出的气孔导度模拟模型大多为统计模型[49],还需更多理论总结进行机理的推求.本文将冠层视为“大叶”,但作物冠层结构复杂,冠层内气孔导度随叶倾角、叶位等的不同而变化,而且当冠层发育完全,叶片开始老化和变皱也会影响对冠层导度的模拟和计算[45].冠层导度是植被总体气孔导度的集合[35],随着环境因子的变化,有的气孔导度增加,有的气孔导度下降,冠层导度表示的是其整体效应[17],与气孔导度的响应变化并不完全一致,而冠层导度模型是基于叶片尺度气孔导度的提升,这也是Jarvis模型的局限性之一.然而,简化冠层结构是模型应用的必要条件[21],尽管如此,冠层导度的估算模型仍能充分反映其随主要环境影响要素的变化规律.本研究基于“单源模型”的假设,只分析了农田郁闭度较高时模型的应用效果,若在LAI较低时应用该冠层导度模型,用考虑土壤蒸发的“双源模型”模拟蒸散可能更准确,这在今后的研究需要考虑. ...
Sap flow, canopy conductance and microclimate in a banana screenhouse
2
2015
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
Spatio-temporal variation in transpiration responses of maize plants to vapor pressure deficit under an arid climatic condition
1
2016
... 冠层导度的模拟大多基于叶片气孔导度,再通过尺度提升方法进行转换,常用的估算方法可分为3类[5]:多元线性回归模型(Multiple correlation models)、现象模型(Phenomenological models)和气孔行为机制模型(Mechanical models),3种模型都属于稳态模型.多元线性回归模型为气孔导度关于气象因子的简单线性模型,由于没有考虑冠层内部叶片接收的光合有效辐射(PhotosyntheticallyActive Radiation,PAR)和温度(T)等环境要素差异,相比于现象模型预测精度较低[6],且无法解释气孔响应环境变化的调控行为;气孔行为机制模型为气孔导度关于净光合速率和环境因子的线性相关模型[7],净光合速率在普通气象站难以获取,而估算模型多为生化模型,计算过程复杂,应用不便.现象模型由环境因子的响应函数和最大气孔导度组成[8,9,10],模型形式简单、灵活,且已被证明能够较为准确地估算陆地不同生态系统的冠层导度,被广泛应用于水文模型[11]、陆面过程模型[12,13]和生态模型[14,15,16]中.然而,气孔导度对主要环境影响要素的响应依赖于环境条件和植物本身,对具体求取某一特定环境条件下的特定植被而言,需要通过观测试验,才能够获取较为准确的模拟冠层导度模型[17].但是以往多数气孔导度模型拟合都只采用了正午时段的观测数据,上午和下午的数据因数值较小多被剔除[18],如孙林等[19]在模拟冠层平均气孔导度时剔除了饱和水汽压差(Vapor Pressure Deficit,VPD)小于0.6 kPa的数据.气孔导度对环境变量响应的研究也出现不同看法,如对于气孔导度和VPD的关系,多数研究认为是负相关,且选择了不同的函数表现形式模拟,如郑海雷[20]认为气孔导度对VPD的响应符合变形倒数函数,Schelde等[21]认为符合线性函数.而Liu等[22]却发现冠层导度与VPD的关系表现为正相关,Zhao等[23]发现气孔导度对VPD的响应存在转折点,当VPD<3.5 kPa时表现为正相关,当VPD≥3.5 kPa时表现为负相关.目前,气孔导度对环境因子变化的全天响应研究仍然较少,值得关注和进一步研究. ...
Modeling stomatal resistance: An overview of the 1989 workshop at the Pennsylvania State University
2
1991
... 在基于叶片气孔导度向冠层导度转换的研究中,直接用阳叶的气孔导度乘以叶面积指数(Leaf Area Index,LAI)的模型会导致对蒸散的严重高估[24],因此应将气孔导度乘以一个遮荫系数()[25].遮荫系数的意义为:通常郁闭度高的作物冠层的上半部分吸收了大部分的净辐射,冠层下方叶子被阻挡了直射光和风,因此水热传输速率比较低[26].与LAI密切相关,数值一般为0.5~1.0,随LAI的增大而减小,前人的研究中多粗略地认为为0.5,并称之为有效叶面积指数[11,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
... Jarvis冠层导度模型有不同的形式,其中有两种比较简单的方法,一是不考虑冠层尺度通量对冠层结构(如LAI和叶倾角等)的依赖关系,这种方法会使冠层结构的效应平均到模型的率定参数中(如)[13].二是假设每个叶片对冠层的水和二氧化碳通量贡献相等,即冠层通量是叶片水平通量乘以LAI[9,33].本文采用的是第二种方法,这种简单的比例关系特别适合在大叶模型中使用,然而模型模拟结果存在的高估问题应采用修正,本文在前人研究的0.5<<1[24]的基础上发现,对于西北干旱区灌溉绿洲农田玉米而言,在农田郁闭度较高,LAI较大时0.5≤≤0.6,且本文得到的与LAI具体函数表达形式可以为模型构建中的准确取值提供必要的方法和参考. ...
Below-canopy fluxes in a simplified one-dimensional theoretical description of the vegetation-atmosphere interaction
1
1979
... 在基于叶片气孔导度向冠层导度转换的研究中,直接用阳叶的气孔导度乘以叶面积指数(Leaf Area Index,LAI)的模型会导致对蒸散的严重高估[24],因此应将气孔导度乘以一个遮荫系数()[25].遮荫系数的意义为:通常郁闭度高的作物冠层的上半部分吸收了大部分的净辐射,冠层下方叶子被阻挡了直射光和风,因此水热传输速率比较低[26].与LAI密切相关,数值一般为0.5~1.0,随LAI的增大而减小,前人的研究中多粗略地认为为0.5,并称之为有效叶面积指数[11,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
Operational estimates of reference evapotranspiration
2
1989
... 在基于叶片气孔导度向冠层导度转换的研究中,直接用阳叶的气孔导度乘以叶面积指数(Leaf Area Index,LAI)的模型会导致对蒸散的严重高估[24],因此应将气孔导度乘以一个遮荫系数()[25].遮荫系数的意义为:通常郁闭度高的作物冠层的上半部分吸收了大部分的净辐射,冠层下方叶子被阻挡了直射光和风,因此水热传输速率比较低[26].与LAI密切相关,数值一般为0.5~1.0,随LAI的增大而减小,前人的研究中多粗略地认为为0.5,并称之为有效叶面积指数[11,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
... ,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
Surface resistance of crop canopies
1
1969
... 在基于叶片气孔导度向冠层导度转换的研究中,直接用阳叶的气孔导度乘以叶面积指数(Leaf Area Index,LAI)的模型会导致对蒸散的严重高估[24],因此应将气孔导度乘以一个遮荫系数()[25].遮荫系数的意义为:通常郁闭度高的作物冠层的上半部分吸收了大部分的净辐射,冠层下方叶子被阻挡了直射光和风,因此水热传输速率比较低[26].与LAI密切相关,数值一般为0.5~1.0,随LAI的增大而减小,前人的研究中多粗略地认为为0.5,并称之为有效叶面积指数[11,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
Turbulence in waving wheat. Part II: Structure of momentum transfer
1
1979
... 在基于叶片气孔导度向冠层导度转换的研究中,直接用阳叶的气孔导度乘以叶面积指数(Leaf Area Index,LAI)的模型会导致对蒸散的严重高估[24],因此应将气孔导度乘以一个遮荫系数()[25].遮荫系数的意义为:通常郁闭度高的作物冠层的上半部分吸收了大部分的净辐射,冠层下方叶子被阻挡了直射光和风,因此水热传输速率比较低[26].与LAI密切相关,数值一般为0.5~1.0,随LAI的增大而减小,前人的研究中多粗略地认为为0.5,并称之为有效叶面积指数[11,26,27].而Rochette等[3]用不同升尺度方法得到冠层导度模型,发现为0.5~0.8,却没有得到的计算方法.目前,关于冠层导度模型中准确取值的研究仍较缺乏,Finnigan[28]认为是冠层上方空气动力学导度与气孔导度比值的函数,然而得到的误差比较大,因此需深入了解的对应关系来模拟其准确取值,以提高冠层导度模型预测精度. ...
Simulation Analysis and Optimal Regulation for Agro-Hydrological Processes and Water Use Efficiency on Multiple Scales of the Middle Heihe River Basin
1
2017
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
黑河中游绿洲多尺度农业水文过程及用水效率的模拟分析与优化调控研究
1
2017
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
Integrated water systems model for terrestrial water cycle simulation
1
2019
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
陆地水循环过程的综合集成与模拟
1
2019
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
Research on climate feedback of human water use and its impact on terrestrial water cycles—Advances and challenges
1
2019
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
. 人类用水活动的气候反馈及其对陆地水循环的影响研究——进展与挑战
1
2019
... 黑河流域地处我国西北干旱区,其特殊的地理环境形成了许多灌溉绿洲,尤其在中游地区,主要作物为玉米,由于缺乏有效的水资源统筹规划与管理措施,集中的农业灌溉消耗了约83%的黑河出山径流量[29],而蒸散是干旱区水分循环的重要组成部分,较为准确估算农田蒸散量可为区域水资源管理和水资源合理决策提供关键科学依据.此外,大多数估算冠层导度的研究对象都是草地和森林,对于行作物研究较少[11].因此,本文利用河西走廊中段临泽绿洲灌溉农田微气象、涡度相关、叶片气体交换等观测试验数据,基于Jarvis模型原理,得出了适用于干旱区灌溉绿洲农田玉米冠层导度的估算模型,这将为干旱区灌溉绿洲农田生态水文过程、水循环和植被—大气相互作用等研究中有关蒸散和水热传输的深入研究提供重要参考[30,31]. ...
Carbon dioxide exchange in an irrigated agricultural field within an oasis, Northwest China
1
2011
... 农田蒸散数据来源于试验地涡度相关系统测算结果,主要由三维超声风速仪(HS50, Gill Solent Instruments,英国)和快速响应红外CO2/H2O气体分析仪(Li-7500,LI-COR Inc.,美国)组成,具体见Ji等[32]对农田通量的研究.气象数据来自试验区的绿洲农田小气候观测系统,本文所用风速(LISA,Siggelkow,德国)、温度和湿度(HMP45D,Vaisala,芬兰)均为2 m处数据;净辐射仪(Kipp & Zonen,Delft,芬兰)位于地面以上4 m处;土壤热通量(HFP01,Hukseflux,荷兰)测量在地面以下5 cm处. ...
Regulation of stomatal conductance and transpiration in forest canopies
2
1998
... 在大叶模型中,冠层通常被认为是叶子随机排列的结果,冠层导度代表了水汽和CO2在冠层内外扩散的难易程度,为尽可能减小土壤蒸发对蒸散的影响,本文选取了LAI大于3的数据进行分析和模型验证.基于Jarvis模型原理,冠层导度估算模型表达式如下[33]: ...
... Jarvis冠层导度模型有不同的形式,其中有两种比较简单的方法,一是不考虑冠层尺度通量对冠层结构(如LAI和叶倾角等)的依赖关系,这种方法会使冠层结构的效应平均到模型的率定参数中(如)[13].二是假设每个叶片对冠层的水和二氧化碳通量贡献相等,即冠层通量是叶片水平通量乘以LAI[9,33].本文采用的是第二种方法,这种简单的比例关系特别适合在大叶模型中使用,然而模型模拟结果存在的高估问题应采用修正,本文在前人研究的0.5<<1[24]的基础上发现,对于西北干旱区灌溉绿洲农田玉米而言,在农田郁闭度较高,LAI较大时0.5≤≤0.6,且本文得到的与LAI具体函数表达形式可以为模型构建中的准确取值提供必要的方法和参考. ...
The environmental responses and simulation of canopy conductance in a winter wheat field of North China Plain
1
2007
... 基于单源模型的假设,将植被冠层视作单层封闭型冠层,忽略土壤蒸发[34],根据“大叶”模型原理,将通过Penman-Monteith公式[35]反推得到的表面导度近似为冠层导度来验证本文提出的冠层导度估算模型. ...
华北平原冬小麦冠层导度的环境响应及模拟
1
2007
... 基于单源模型的假设,将植被冠层视作单层封闭型冠层,忽略土壤蒸发[34],根据“大叶”模型原理,将通过Penman-Monteith公式[35]反推得到的表面导度近似为冠层导度来验证本文提出的冠层导度估算模型. ...
A recommendation on standardized surface resistance for hourly calculation of reference ETo by the FAO56 Penman-Monteith method
2
2006
... 基于单源模型的假设,将植被冠层视作单层封闭型冠层,忽略土壤蒸发[34],根据“大叶”模型原理,将通过Penman-Monteith公式[35]反推得到的表面导度近似为冠层导度来验证本文提出的冠层导度估算模型. ...
... 尽管本文得到的Jarvis冠层导度模型能比较好地模拟干旱区灌溉绿洲农田玉米冠层导度的响应变化,但本质上是经验模型,对于新的植物种类和环境条件需重新对模型进行参数化,而且目前提出的气孔导度模拟模型大多为统计模型[49],还需更多理论总结进行机理的推求.本文将冠层视为“大叶”,但作物冠层结构复杂,冠层内气孔导度随叶倾角、叶位等的不同而变化,而且当冠层发育完全,叶片开始老化和变皱也会影响对冠层导度的模拟和计算[45].冠层导度是植被总体气孔导度的集合[35],随着环境因子的变化,有的气孔导度增加,有的气孔导度下降,冠层导度表示的是其整体效应[17],与气孔导度的响应变化并不完全一致,而冠层导度模型是基于叶片尺度气孔导度的提升,这也是Jarvis模型的局限性之一.然而,简化冠层结构是模型应用的必要条件[21],尽管如此,冠层导度的估算模型仍能充分反映其随主要环境影响要素的变化规律.本研究基于“单源模型”的假设,只分析了农田郁闭度较高时模型的应用效果,若在LAI较低时应用该冠层导度模型,用考虑土壤蒸发的“双源模型”模拟蒸散可能更准确,这在今后的研究需要考虑. ...
Field studies of the conductance of wheat leaves and transpiration
2
1976
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... Fitting functions and its coefficient of determination Table 1| 环境变量 | 响应函数 | 7:00-11:00 | 11:00-17:00 | 17:00-21:00 |
|---|
| 参数取值 | 决定系数 | 参数取值 | 决定系数 | 参数取值 | 决定系数 |
|---|
| PAR | [36] | a1=0.051 ...
Stomatal factor in the water use and dry matter production by sorghum
2
1973
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... a2=0.0002 | 0.351 | | [37] | a1=0.066 ...
Canopy resistance formulation and its effect in mesoscale models: A HAPEX perspective
2
1991
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... a2=0.002 | 0.359 | | [38] | a1=0.067 ...
Two-dimensional model of water transport in the root zone and plant for container-grown chrysanthemum
2
1992
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... b2=0.017 | 0.106 | | [39] | b1=0.051 ...
Mathematical simulation of responses of wheat stomata to environment factors in the field
3
1988
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... b2=-8.765 | 0.081 | | T | [40] | c1=-0.177 ...
... c2=0.004 | 0.063 | | [40] | c1=-0.984 ...
田间小麦叶片气孔对环境因子响应的模拟及叶片水分平衡的计算
3
1988
... 由于Jarvis模型为经验模型,模型假定植物气孔导度与环境变量间存在函数关系并且各环境变量独立地对植物气孔导度产生影响[8],因此采用常用函数对各环境变量分别拟合(表1).应用较普遍的函数中,气孔导度对PAR的响应有线性函数、指数函数和直角双曲线函数[36,37,38]等;对VPD的响应有线性函数、负指数函数和变形倒数函数[8,21,39]等;对T的响应主要有线性函数和二次函数[40].从表1给出的结果可以看出,7:00-11:00叶片气孔导度对各环境变量响应的敏感度依次为:PAR、T、VPD,即气孔张开过程主要受光照的影响;11:00-17:00敏感度顺序变为:VPD、T、PAR,这个时段气孔主要受VPD限制,以防止蒸腾过快;17:00-21:00相比于前两个时段的敏感度较差,顺序为PAR、VPD、T,随着光照强度减弱气孔逐渐关闭.第一和第三时段气孔导度对各环境变量的响应均表现为正相关,第二时段气孔导度对VPD和T的响应变为负相关,对PAR的响应无明显改变.得到各环境变量的最优响应函数:PAR为变形倒数函数,T为二次函数,在第一时段和第三时段VPD为线性函数,第二时段为指数函数.因此对于叶片气孔导度日响应过程各时段的模拟应当选用不同函数形式,模型具体表达式如表2所列. ...
... b2=-8.765 | 0.081 | | T | [40] | c1=-0.177 ...
... c2=0.004 | 0.063 | | [40] | c1=-0.984 ...
Net ecosystemexchange of CO2 on sunny and cloudy days over a reed wetland in the Yellow River Delta, China
1
2015
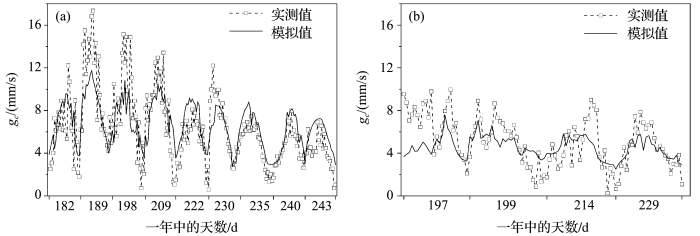
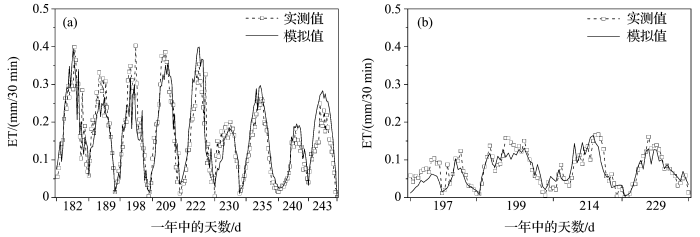
... 在玉米拔节期之后7月和8月,LAI一般为3~4,拟合得到在该范围内0.5≤≤0.6,随LAI增大而减小.以叶片尺度气孔导度模型为基础,应用推导的计算公式和实测LAI值升尺度转换得到基于Jarvis模型原理的各时段冠层导度模型,用2010年玉米生长季晴天的数据模拟冠层导度,并在半小时尺度上,用涡度相关数据结合Penman-Monteith公式反推得到的冠层导度实测值对模型进行验证(图4a),决定系数为0.665,实测冠层导度平均值为6.205 mm/s.试验发现气孔导度对辐射变化很敏感,为了验证冠层导度模型在太阳辐射变化较小情况下的效果,依据初小静等[41]的天气划分方法,我们选择了阴天和多云天气各2天的观测数据,对干旱区灌溉绿洲农田生态系统冠层导度进行模拟(图4b),模型决定系数为0.606,实测冠层导平均值为4.836 mm/s.表明基于Jarvis模型原理得到的冠层导度估算模型适合当地不同天气条件. ...
阴天和晴天对黄河三角洲芦苇湿地净生态系统CO2交换的影响
1
2015
... 在玉米拔节期之后7月和8月,LAI一般为3~4,拟合得到在该范围内0.5≤≤0.6,随LAI增大而减小.以叶片尺度气孔导度模型为基础,应用推导的计算公式和实测LAI值升尺度转换得到基于Jarvis模型原理的各时段冠层导度模型,用2010年玉米生长季晴天的数据模拟冠层导度,并在半小时尺度上,用涡度相关数据结合Penman-Monteith公式反推得到的冠层导度实测值对模型进行验证(图4a),决定系数为0.665,实测冠层导度平均值为6.205 mm/s.试验发现气孔导度对辐射变化很敏感,为了验证冠层导度模型在太阳辐射变化较小情况下的效果,依据初小静等[41]的天气划分方法,我们选择了阴天和多云天气各2天的观测数据,对干旱区灌溉绿洲农田生态系统冠层导度进行模拟(图4b),模型决定系数为0.606,实测冠层导平均值为4.836 mm/s.表明基于Jarvis模型原理得到的冠层导度估算模型适合当地不同天气条件. ...
(
2
2015
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
... [42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
Water flux and canopy conductance of natural versus planted forests in Patagonia, South America
1
2009
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
A generic model of forest canopy conductance dependent on climate, soil water availability and leaf area index
1
2000
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
Effects of stand age and tree species on canopy transpiration and average stomatal conductance of boreal forests
2
2005
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
... 尽管本文得到的Jarvis冠层导度模型能比较好地模拟干旱区灌溉绿洲农田玉米冠层导度的响应变化,但本质上是经验模型,对于新的植物种类和环境条件需重新对模型进行参数化,而且目前提出的气孔导度模拟模型大多为统计模型[49],还需更多理论总结进行机理的推求.本文将冠层视为“大叶”,但作物冠层结构复杂,冠层内气孔导度随叶倾角、叶位等的不同而变化,而且当冠层发育完全,叶片开始老化和变皱也会影响对冠层导度的模拟和计算[45].冠层导度是植被总体气孔导度的集合[35],随着环境因子的变化,有的气孔导度增加,有的气孔导度下降,冠层导度表示的是其整体效应[17],与气孔导度的响应变化并不完全一致,而冠层导度模型是基于叶片尺度气孔导度的提升,这也是Jarvis模型的局限性之一.然而,简化冠层结构是模型应用的必要条件[21],尽管如此,冠层导度的估算模型仍能充分反映其随主要环境影响要素的变化规律.本研究基于“单源模型”的假设,只分析了农田郁闭度较高时模型的应用效果,若在LAI较低时应用该冠层导度模型,用考虑土壤蒸发的“双源模型”模拟蒸散可能更准确,这在今后的研究需要考虑. ...
Characterizing the diurnal patterns of errors in the prediction of evapotranspiration by several land-surface models: An NACP analysis
1
2014
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
The hysteretic evapotranspiration-Vapor pressure deficit relation
1
2014
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
An overview of researches on land-atmosphere interaction over semi-arid region under global changes
1
2018
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
全球变化背景下半干旱区陆气机制研究综述
1
2018
... 气孔导度与VPD和T的关系呈现顺时针“非对称性响应”,这一变化规律与其他植物如绿洲生态系统中的葡萄[42]和沙区的侧柏[18]一致.目前,已有很多研究发现气孔导度和蒸腾对环境因子的滞后响应[22,43],Granier等[44]的研究还表明考虑时间滞后的蒸腾和环境因子的决定系数比不考虑时滞提高近1倍;Ewers等[45]发现蒸腾对PAR的滞后值较VPD大,且对PAR的“非对称性响应”表现为逆时针而对VPD表现为“顺时针”,而本文认为气孔导度对PAR并没有表现出明显的滞后响应.国内外学者对于滞后产生的原因有着不同的看法,Matheny等[46]认为这些误差很大程度上是由模型缺乏对植物水动力学的描述造成的,叶片需水量与土壤供水量之间的不平衡对气孔导度造成了水动力限制;Zhang等[47]认为随着土壤含水量下降,植物和土壤水势是调节滞后关系的关键因素.植物气孔对环境因子的响应过程十分复杂,因此将全天冠层导度和蒸腾与环境因子一起进行统计分析的方法并不能揭示植物生理过程和环境变化过程的调控机理[18],分析白天不同时段气孔导度对环境变量的响应差异也有助于更好地理解气孔的整体行为[42],将生产实际中的逐日资料分时段统计建模可以获得更准确的冠层导度和蒸腾全日变化预测值,为当地农业管理提供理论依据,而且蒸散是干旱区绿洲农田生态系统能量平衡的主要部分[48],也是天气预报和气候建模的依据之一. ...
A combination model for estimating stomatal conductance of maize (Zea mays L.) leaves over a long term
1
1998
... 尽管本文得到的Jarvis冠层导度模型能比较好地模拟干旱区灌溉绿洲农田玉米冠层导度的响应变化,但本质上是经验模型,对于新的植物种类和环境条件需重新对模型进行参数化,而且目前提出的气孔导度模拟模型大多为统计模型[49],还需更多理论总结进行机理的推求.本文将冠层视为“大叶”,但作物冠层结构复杂,冠层内气孔导度随叶倾角、叶位等的不同而变化,而且当冠层发育完全,叶片开始老化和变皱也会影响对冠层导度的模拟和计算[45].冠层导度是植被总体气孔导度的集合[35],随着环境因子的变化,有的气孔导度增加,有的气孔导度下降,冠层导度表示的是其整体效应[17],与气孔导度的响应变化并不完全一致,而冠层导度模型是基于叶片尺度气孔导度的提升,这也是Jarvis模型的局限性之一.然而,简化冠层结构是模型应用的必要条件[21],尽管如此,冠层导度的估算模型仍能充分反映其随主要环境影响要素的变化规律.本研究基于“单源模型”的假设,只分析了农田郁闭度较高时模型的应用效果,若在LAI较低时应用该冠层导度模型,用考虑土壤蒸发的“双源模型”模拟蒸散可能更准确,这在今后的研究需要考虑. ...
/
|
〈
|
![]()
|
〉
|
|
|






 甘公网安备62010202000687
甘公网安备62010202000687{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}