Introduction to Organic Geochemistry (second edition)
1
2005
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
Building the biomarker tree of life
2
2005
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
Geomicrobial functional groups: A window on the interaction between life and environments
1
2012
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
地质微生物功能群:生命与环境相互作用的重要突破口
1
2012
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
Application of plant derived n -alkanes and their compound specific hydrogen isotopic composition in paleoenvironment research
1
2009
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
植物正构烷烃及其单体氢同位素在古环境研究中的应用
1
2009
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
An overview on microbial proxies for the reconstruction of past global environmental change
2
2013
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
示踪全球环境变化的微生物代用指标
2
2013
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
Lipid biomarkers for the reconstruction of deep-time environmental conditions
2
2019
... 脂类(lipid)是指由生物合成的、不溶于水而溶于甲醇、氯仿、二氯甲烷等非极性溶剂的生物化学成分[1 ] .脂类来自生物体,主要分布于细胞膜、叶片蜡质层等部位.脂类化合物种类众多,有正构脂类,也有异构脂类和环状脂类.某些特征的脂类专属于特定的生物分类学类群或微生物功能群[2 ,3 ] ,可以充当这些生物类群或代谢途径的标志物.例如,2-甲基藿类主要来自蓝细菌,长链烯酮来源于颗石藻,非常负偏的碳同位素组成(<-50‰)可以指示甲烷氧化过程.由于能够记录生物源、代谢途径、生物生长或早期成岩转化过程中的生态环境信息,脂类已经成为生物—有机地球化学、新生代古气候与古生态研究等领域的重要工具.已有较多的文章综述了脂类在古环境和古生态方面的研究进展[4 ,5 ,6 ] . ...
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
Isotope-ratio-monitoring gas chromatography-mass spectrometry
1
1978
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Isotope-ratio detection for gas chromatography
2
2006
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
... 除了分离纯化,单体碳分析技术还有一个很重要的环节,就是同位素结果的校正[8 ] .通过色谱分离后的脂类在氧化炉中被在线燃烧转化为CO2 气体.这个过程需要CuO或CuO-NiO释放氧气,因此需要定期通氧来保证反应器的供氧能力.由于燃烧过程是在高温条件(900~1 000 ℃)下完成,如此高的温度有可能造成碳同位素分馏,因而需要在样品分析中加入碳同位素值已知的内标化合物来校正燃烧过程对碳同位素的影响.同时,还需要定期运行碳同位素值已知的标样来定期检验仪器的状态.目前,大部分实验室采用的是美国Indiana大学Arndt Schimmelmann博士实验室配置的单个正构烷烃或混合标样.最近Schimmelmann博士联合多家国际实验室,建立了多种新的同位素参考物质,其中包括正构碳十六烷(编号为USGS67, USGS68和USGS69,分别对应于不同负偏的碳同位素值)[17 ] . ...
Pitfalls in compound-specific isotope analysis of environmental samples
1
2008
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Current challenges in compound-specific stable isotope analysis of environmental organic contaminants
1
2012
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Chain-length distribution of n -alkanes from lake sediments in relation to post-glacial environmental change
1
1973
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Comparison of carbon isotopic composition of the saturated hydrocarbons before and after complexation 5? molecular sieve
1
2005
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
饱和烃经5 ?分子筛络合前后单体烃碳同位素分析对比研究
1
2005
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
An improved and rapid 5A molecular sieve method for gas chromatography isotope ratio mass spectrometry of n -alkanes (C8 -C30+ )
1
2008
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Purification of dinosterol for hydrogen isotopic analysis using high-performance liquid chromatography-mass spectrometry
1
2007
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Purification of dinosterol from complex mixtures of sedimentary lipids for hydrogen isotope analysis
1
2012
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Derivatization of organic compounds prior to gas chromatographic-combustion-isotope ratio mass spectrometric analysis: Identification of isotope fractionation processes
1
1994
... 受益于已故美国科学院院士John M. Hayes教授课题组的开创性贡献[7 ] ,以及后续技术改进,单体碳同位素分析技术已经成为古生物地球化学过程研究的重要技术.脂类单体碳同位素分析包括几个重要的环节:化合物分离,在线燃烧转化为CO2 ,CO2 碳同位素比值测定,以及同位素结果的校正[8 ,9 ,10 ] .在这些环节中,化合物的分离纯化非常关键.对于新鲜的植物样品或有机质含量比较高的泥炭或湖相沉积物样品,长链正构烷烃的含量通常比较高,且其他组分的干扰比较弱,因而硅胶柱层析后得到的烷烃组分可以直接上机进行测试.在一些情况下,植物或土壤样品中含有较高含量的长链烯烃,可以在柱层析环节添加硝酸银来去除烯烃[11 ] .对于其他的样品,需要考虑色谱峰之间的干扰,尤其是鼓包(unresolved complex mixture),在很多情况下需要进行分子筛络合或尿素络合[12 ,13 ] ,消除异构烷烃和环烷烃的干扰.对于一些非常关键的化合物,还可以采用制备液相色谱法来进行单个化合物的纯化[14 ,15 ] .极性脂类如脂肪酸和脂肪醇等要进行衍生化后才能上机测试,常规的衍生化方法有硅烷化和甲酯化.这些衍生化过程会加入额外的碳原子,需要对最终的测试结果进行校正[16 ] .一方面,尽量少改变原化合物的碳原子数量;另一方面,需要测定衍生化过程中加入的碳原子的碳同位素组成. ...
Organic reference materials for hydrogen, carbon, and nitrogen stable isotope-ratio measurements: Caffeines, n -alkanes, fatty acid, methyl esters, glycines, L-valines, polyethylenes, and oils
1
2016
... 除了分离纯化,单体碳分析技术还有一个很重要的环节,就是同位素结果的校正[8 ] .通过色谱分离后的脂类在氧化炉中被在线燃烧转化为CO2 气体.这个过程需要CuO或CuO-NiO释放氧气,因此需要定期通氧来保证反应器的供氧能力.由于燃烧过程是在高温条件(900~1 000 ℃)下完成,如此高的温度有可能造成碳同位素分馏,因而需要在样品分析中加入碳同位素值已知的内标化合物来校正燃烧过程对碳同位素的影响.同时,还需要定期运行碳同位素值已知的标样来定期检验仪器的状态.目前,大部分实验室采用的是美国Indiana大学Arndt Schimmelmann博士实验室配置的单个正构烷烃或混合标样.最近Schimmelmann博士联合多家国际实验室,建立了多种新的同位素参考物质,其中包括正构碳十六烷(编号为USGS67, USGS68和USGS69,分别对应于不同负偏的碳同位素值)[17 ] . ...
Preservation and diagenesis of hopanoids in recent lacustrine sediments of Priest Pot, England
1
1997
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
Lipids of symbiotic methane-oxidizing bacteria in peat moss studied using stable carbon isotopic labelling
2
2010
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
δ13 C values and radiocarbon dates of microbial biomarkers as tracers for carbon recycling in peat deposits
2
2000
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
Carbon isotopic composition of branched tetraether membrane lipids in soils suggest a rapid turnover and a heterotrophic lifestyle of their source organism(s)
3
2010
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
... [21 ].在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
Carbon isotopic composition of branched tetraether membrane lipids in a loess-paleosol sequence and its geochemical significance
1
2018
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
Stable carbon isotope ratios of intact GDGTs indicate heterogeneous sources to marine sediments
1
2016
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
Carbon isotopic composition of intact branched GDGT core lipids in Greenland Lake sediments and soils
3
2017
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
A novel method to measure the 13 C composition of intact bacteriohopanepolyols
2
2018
... 在单体碳同位素分析方面,还需要提及常规气相色谱仪难以气化的脂类,主要是藿类和四醚类.传统上,需要将具有多种官能团的藿类经氢碘酸/硼氢化钠反应后,降解成藿醇,然后进行单体碳同位素分析[18 ,19 ] .四醚类化合物也需要断裂醚键,转变为支链烷烃来进行单体碳同位素分析[20 ,21 ,22 ] .最近,Pearson等[23 ] 建立了一种能够直接测试完整四醚膜脂(Glycerol Dialkyl Glycerol Tetraether Lipids,GDGTs)碳同位素组成的新方法.该方法已被用于格陵兰湖泊沉积物中完整GDGTs单体碳同位素组成的研究[24 ] .Hemingway等[25 ] 建立了一种可以直接测试完整的细菌藿多醇(Bacteriohopanepolyols,BHPs)单体碳同位素组成的新方法. ...
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
Leaf epicuticular waxes
1
1967
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
Composition of plant cuticular waxes
1
2006
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
Plant cuticular Waxes: Composition, function, and interactions with microorganisms
1
2018
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
Significance of long chain iso and anteiso monomethyl alkanes in the Lamiaceae (mint family)
1
2011
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
Concentrations and δ2 H values of cuticular n -alkanes vary significantly among plant organs,species and habitats in grasses from an alpine and a temperate European grassland
1
2015
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
Distribution and carbon isotope patterns of diterpenoids and triterpenoids in modern temperate C3 trees and their geochemical significance
3
2012
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
... 三萜类和叶蜡脂类都来自高等植物,在响应气候变化方面,它们的碳同位素组成是表现出相似还是差异的响应是值得关注的科学问题.不过,相关的研究还非常少.一项在非洲东部Lake Challa开展的研究显示,从末次冰盛期以来,湖泊沉积柱中C31 正构烷烃和脱A-环羽扇烷(羽扇烷的一种降解产物)的碳同位素表现出了非常不一样的变化趋势[81 ] .后者的变化非常平缓,而前者则表现出了明显的变化,在末次冰消期C31 正构烷烃的碳同位素组成相对偏正,到了全新世,2种脂类的碳同位素组成的变化则非常相似.对于这种情况,作者解释为脱A-环羽扇烷主要来自C3 植物,而C31 正构烷烃同时来自C3 和C4 植物.基于此,作者提出可以将主要来自C3 植物的脱A-环羽扇烷的δ13 C值作为对应时段的C3 植物δ13 C值的端元值,由此可以更为准确地估算流域内C4 植物的比率[81 ] .这其中需要考虑合成途径差异而造成的正构烷烃和三萜类碳同位素的差值[31 ,82 ] .由此可见,综合多种脂类的碳同位素组成能够更准确地揭示古生态和古环境的变化,还有可能发掘出更多的环境信息. ...
Leaf wax n -alkane distributions in and across modern plants:Implications for paleoecology and chemotaxonomy
1
2013
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
Extracting the most from terrestrial plantderived n -alkyl lipids and their carbon isotopes from the sedimentary record:A review
6
2017
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
... 高等植物脂类中的碳原子来源于植物光合作用固定的CO2 ,它们的碳同位素组成会继承光合作用过程中的碳同位素分馏值(通常被记做Δleaf )[33 ] 和脂类生物合成中的同位素分馏值(εlipid )[33 ] ,因而能够记录相应的温度、干湿等环境条件和植物生理过程的变化[33 ,34 ] . ...
... [33 ],因而能够记录相应的温度、干湿等环境条件和植物生理过程的变化[33 ,34 ] . ...
... [33 ,34 ]. ...
... 在纯C3 植物或C4 植物占比非常少的环境中,例如森林和淡水沼泽,δ13 Calk 变化通常被解释为相对湿度变化[40 ,41 ] .其中的机制是,大气湿度调控叶片气孔的开闭程度,进而影响到大气CO2 向叶片内部的扩散速率.一般来讲,在湿润条件下,如热带雨林,光合作用过程中碳同位素的分馏系数受CO2 扩散过程的影响较弱,因而Rubisco酶的分馏效应起到主导作用,合成的有机质表现出更偏负的碳同位素值.前期研究显示,叶片光合作用碳同位素分馏值(Δleaf )与降雨量之间存在显著的相关性[42 ] .在一项新近的美国亚热带Lake Tulane末次冰期古生态研究中,Arnold等[43 ] 基于湖泊沉积物中长链烷烃δ13 C值估算了Δleaf 值.结果发现,在Heinrich事件2~4时期,Δleaf 处于高值,而长链烷烃δD值处于低值,揭示出了末次冰期冷事件时的冷湿的气候特征.除了相对湿度,δ13 Calk 值还受CO2 浓度及其碳同位素组成、温度、植物生活型等多种因素的影响[33 ,44 ] . ...
... 在解译湖沼沉积物中δ13 Calk 信号时,还需要考虑从枯枝落叶堆积到被流水搬运等过程中的微生物降解及潜在的微生物输入[33 ,45 ,46 ,47 ] .由于微生物偏向于利用轻碳,有机质在经过微生物降解作用后,某些易被降解的脂类单体碳同位素组成可能会发生偏正.此外,在搬运过程中微生物自身产生的有机质输入也可能会对单体碳同位素的解译产生影响,特别是一些自养微生物产生的脂类可能具有比较偏负的碳同位素组成.气溶胶传输也可能是湖泊沉积物中保存的陆生叶蜡烷烃的一种潜在输入途径[48 ,49 ] . ...
13 C/12 C signatures in plants and algae
3
2014
... 高等植物能够合成种类众多的脂类,包括长链正构烷烃、正构脂肪酸和脂肪醇,非藿五环三萜类(下文中简称为三萜类),以及一些C29 甾醇[26 ,27 ,28 ] .其中长链正构脂类(包括烷烃,脂肪酸和脂肪醇,碳链长度>C21 )主要分布于叶片的表皮蜡层,因而常常被称为叶蜡脂类.实际上,长链正构脂类也少量地分布于其他器官的表皮中[29 ,30 ] .三萜类主要分布于常绿阔叶树的叶片和树皮等中[31 ] .前期已有文章总结了高等植物脂类组成和含量在生物体中的分布特征[32 ,33 ,34 ] . ...
... 高等植物脂类中的碳原子来源于植物光合作用固定的CO2 ,它们的碳同位素组成会继承光合作用过程中的碳同位素分馏值(通常被记做Δleaf )[33 ] 和脂类生物合成中的同位素分馏值(εlipid )[33 ] ,因而能够记录相应的温度、干湿等环境条件和植物生理过程的变化[33 ,34 ] . ...
... 相对于长链正构烷烃,长链正构脂肪酸(δ13 Cfa )和脂肪醇的单体碳同位素组成研究很少,尤其是脂肪醇单体碳同位素组成.从叶蜡中脂类组成来看,叶片中往往含有更高含量的长链正构烷烃[64 ] .Chikaraishi[34 ] 汇总了目前已经发表的陆生C3 植物中各种叶蜡脂类的δ13 C值分布范围.在沉积序列研究中,一些研究组专注于δ13 Cfa ,而另一些研究组侧重于δ13 Calk .在同一套沉积样品中,δ13 Cfa 和δ13 Calk 的响应是否一致,这是值得关注的问题,目前这方面的对比研究还十分欠缺.这涉及到不同脂类碳同位素序列的对比问题,也可能从不同脂类碳同位素的差异上挖掘出更多的环境或生态信息. ...
Biogeochemical evidence of Holocene East Asian summer and winter monsoon variability from a tropical maar lake in southern China
1
2015
... 长链正构烷烃单体碳同位素组成(δ13 Calk )已经得到了广泛的研究和应用.在大多数情况下,δ13 Calk 信号被解释为C3 与C4 植物相对比率的变化[35 ,36 ,37 ] .在光合作用过程中,C3 与C4 植物采用不同的固碳途径,并表现出有显著差异的碳同位素分馏系数.相对于C4 植物,C3 植物合成的有机物具有更偏负的碳同位素值(典型的C3 植物总有机碳同位素值为-25‰~-27‰,而C4 植物典型值为-11‰~-12‰)[38 ] .从光合作用产物到叶蜡正构烷烃,会发生进一步的碳同位素分馏,使得陆生C3 植物叶蜡烷烃的δ13 C均值为-33.1‰,而陆生C4 植物叶蜡烷烃的δ13 C均值为-21.7‰[39 ] . ...
Spatiotemporal evolution of C3 /C4 vegetation and its controlling factors in southern China since the last glacial maximum
1
2018
... 长链正构烷烃单体碳同位素组成(δ13 Calk )已经得到了广泛的研究和应用.在大多数情况下,δ13 Calk 信号被解释为C3 与C4 植物相对比率的变化[35 ,36 ,37 ] .在光合作用过程中,C3 与C4 植物采用不同的固碳途径,并表现出有显著差异的碳同位素分馏系数.相对于C4 植物,C3 植物合成的有机物具有更偏负的碳同位素值(典型的C3 植物总有机碳同位素值为-25‰~-27‰,而C4 植物典型值为-11‰~-12‰)[38 ] .从光合作用产物到叶蜡正构烷烃,会发生进一步的碳同位素分馏,使得陆生C3 植物叶蜡烷烃的δ13 C均值为-33.1‰,而陆生C4 植物叶蜡烷烃的δ13 C均值为-21.7‰[39 ] . ...
Biomarkers for terrestrial plants and climate
1
2014
... 长链正构烷烃单体碳同位素组成(δ13 Calk )已经得到了广泛的研究和应用.在大多数情况下,δ13 Calk 信号被解释为C3 与C4 植物相对比率的变化[35 ,36 ,37 ] .在光合作用过程中,C3 与C4 植物采用不同的固碳途径,并表现出有显著差异的碳同位素分馏系数.相对于C4 植物,C3 植物合成的有机物具有更偏负的碳同位素值(典型的C3 植物总有机碳同位素值为-25‰~-27‰,而C4 植物典型值为-11‰~-12‰)[38 ] .从光合作用产物到叶蜡正构烷烃,会发生进一步的碳同位素分馏,使得陆生C3 植物叶蜡烷烃的δ13 C均值为-33.1‰,而陆生C4 植物叶蜡烷烃的δ13 C均值为-21.7‰[39 ] . ...
The early origins of terrestrial C4 photosynthesis
1
2007
... 长链正构烷烃单体碳同位素组成(δ13 Calk )已经得到了广泛的研究和应用.在大多数情况下,δ13 Calk 信号被解释为C3 与C4 植物相对比率的变化[35 ,36 ,37 ] .在光合作用过程中,C3 与C4 植物采用不同的固碳途径,并表现出有显著差异的碳同位素分馏系数.相对于C4 植物,C3 植物合成的有机物具有更偏负的碳同位素值(典型的C3 植物总有机碳同位素值为-25‰~-27‰,而C4 植物典型值为-11‰~-12‰)[38 ] .从光合作用产物到叶蜡正构烷烃,会发生进一步的碳同位素分馏,使得陆生C3 植物叶蜡烷烃的δ13 C均值为-33.1‰,而陆生C4 植物叶蜡烷烃的δ13 C均值为-21.7‰[39 ] . ...
A 35 Myr North American leaf-wax compound-specific carbon and hydrogen isotope record:Implications for C4 grasslands and hydrologic cycle dynamics
1
2010
... 长链正构烷烃单体碳同位素组成(δ13 Calk )已经得到了广泛的研究和应用.在大多数情况下,δ13 Calk 信号被解释为C3 与C4 植物相对比率的变化[35 ,36 ,37 ] .在光合作用过程中,C3 与C4 植物采用不同的固碳途径,并表现出有显著差异的碳同位素分馏系数.相对于C4 植物,C3 植物合成的有机物具有更偏负的碳同位素值(典型的C3 植物总有机碳同位素值为-25‰~-27‰,而C4 植物典型值为-11‰~-12‰)[38 ] .从光合作用产物到叶蜡正构烷烃,会发生进一步的碳同位素分馏,使得陆生C3 植物叶蜡烷烃的δ13 C均值为-33.1‰,而陆生C4 植物叶蜡烷烃的δ13 C均值为-21.7‰[39 ] . ...
Alkanes, compound-specific carbon isotope measures and climate variation during the last millennium from varved sediments of Lake Xiaolongwan, Northeast China
2
2013
... 在纯C3 植物或C4 植物占比非常少的环境中,例如森林和淡水沼泽,δ13 Calk 变化通常被解释为相对湿度变化[40 ,41 ] .其中的机制是,大气湿度调控叶片气孔的开闭程度,进而影响到大气CO2 向叶片内部的扩散速率.一般来讲,在湿润条件下,如热带雨林,光合作用过程中碳同位素的分馏系数受CO2 扩散过程的影响较弱,因而Rubisco酶的分馏效应起到主导作用,合成的有机质表现出更偏负的碳同位素值.前期研究显示,叶片光合作用碳同位素分馏值(Δleaf )与降雨量之间存在显著的相关性[42 ] .在一项新近的美国亚热带Lake Tulane末次冰期古生态研究中,Arnold等[43 ] 基于湖泊沉积物中长链烷烃δ13 C值估算了Δleaf 值.结果发现,在Heinrich事件2~4时期,Δleaf 处于高值,而长链烷烃δD值处于低值,揭示出了末次冰期冷事件时的冷湿的气候特征.除了相对湿度,δ13 Calk 值还受CO2 浓度及其碳同位素组成、温度、植物生活型等多种因素的影响[33 ,44 ] . ...
... 对于湖泊等沉积场所中长链正构脂类的研究,还需要注意原地生物贡献的影响.通过对青海湖水生植物和表层沉积物中长链脂类的研究,Liu等[50 ] 提出水生的藻类和沉水植物也能贡献长链正构脂类,这些水生植物产生的长链正构烷烃的δ13 Calk 值非常接近陆地C4 植物的特征,因而会对评估流域内C4 植物的比率产生干扰.值得关注的是,湖泊内源贡献的长链脂类主要是C29 及以下正构烷烃、C30 及以下正构脂肪酸,而C31 及以上碳链的正构烷烃主要是流域内陆生植物的贡献,因此,作者提出C31 正构烷烃的δ13 Calk 值可以更准确地反映陆生植物对气候环境变化的响应[50 ] .此外,在一些湖泊中,水生生物还有可能利用来自甲烷的碳,使得合成的正构烷烃δ13 Calk 值显著偏负[40 ] .因此,对于湖沼沉积序列,在解释古气候和古环境变化时,要充分考虑沉积环境的多样性,具体问题具体分析. ...
An n -alkane and carbon isotope record during the last deglaciation from annually laminated sediment in Lake Xiaolongwan, northeastern China
1
2016
... 在纯C3 植物或C4 植物占比非常少的环境中,例如森林和淡水沼泽,δ13 Calk 变化通常被解释为相对湿度变化[40 ,41 ] .其中的机制是,大气湿度调控叶片气孔的开闭程度,进而影响到大气CO2 向叶片内部的扩散速率.一般来讲,在湿润条件下,如热带雨林,光合作用过程中碳同位素的分馏系数受CO2 扩散过程的影响较弱,因而Rubisco酶的分馏效应起到主导作用,合成的有机质表现出更偏负的碳同位素值.前期研究显示,叶片光合作用碳同位素分馏值(Δleaf )与降雨量之间存在显著的相关性[42 ] .在一项新近的美国亚热带Lake Tulane末次冰期古生态研究中,Arnold等[43 ] 基于湖泊沉积物中长链烷烃δ13 C值估算了Δleaf 值.结果发现,在Heinrich事件2~4时期,Δleaf 处于高值,而长链烷烃δD值处于低值,揭示出了末次冰期冷事件时的冷湿的气候特征.除了相对湿度,δ13 Calk 值还受CO2 浓度及其碳同位素组成、温度、植物生活型等多种因素的影响[33 ,44 ] . ...
Global patterns in leaf 13 C discrimination and implications for studies of past and future climate
1
2010
... 在纯C3 植物或C4 植物占比非常少的环境中,例如森林和淡水沼泽,δ13 Calk 变化通常被解释为相对湿度变化[40 ,41 ] .其中的机制是,大气湿度调控叶片气孔的开闭程度,进而影响到大气CO2 向叶片内部的扩散速率.一般来讲,在湿润条件下,如热带雨林,光合作用过程中碳同位素的分馏系数受CO2 扩散过程的影响较弱,因而Rubisco酶的分馏效应起到主导作用,合成的有机质表现出更偏负的碳同位素值.前期研究显示,叶片光合作用碳同位素分馏值(Δleaf )与降雨量之间存在显著的相关性[42 ] .在一项新近的美国亚热带Lake Tulane末次冰期古生态研究中,Arnold等[43 ] 基于湖泊沉积物中长链烷烃δ13 C值估算了Δleaf 值.结果发现,在Heinrich事件2~4时期,Δleaf 处于高值,而长链烷烃δD值处于低值,揭示出了末次冰期冷事件时的冷湿的气候特征.除了相对湿度,δ13 Calk 值还受CO2 浓度及其碳同位素组成、温度、植物生活型等多种因素的影响[33 ,44 ] . ...
Climate response of the Florida Peninsula to Heinrich events in the North Atlantic
1
2018
... 在纯C3 植物或C4 植物占比非常少的环境中,例如森林和淡水沼泽,δ13 Calk 变化通常被解释为相对湿度变化[40 ,41 ] .其中的机制是,大气湿度调控叶片气孔的开闭程度,进而影响到大气CO2 向叶片内部的扩散速率.一般来讲,在湿润条件下,如热带雨林,光合作用过程中碳同位素的分馏系数受CO2 扩散过程的影响较弱,因而Rubisco酶的分馏效应起到主导作用,合成的有机质表现出更偏负的碳同位素值.前期研究显示,叶片光合作用碳同位素分馏值(Δleaf )与降雨量之间存在显著的相关性[42 ] .在一项新近的美国亚热带Lake Tulane末次冰期古生态研究中,Arnold等[43 ] 基于湖泊沉积物中长链烷烃δ13 C值估算了Δleaf 值.结果发现,在Heinrich事件2~4时期,Δleaf 处于高值,而长链烷烃δD值处于低值,揭示出了末次冰期冷事件时的冷湿的气候特征.除了相对湿度,δ13 Calk 值还受CO2 浓度及其碳同位素组成、温度、植物生活型等多种因素的影响[33 ,44 ] . ...
Disentangling temperature effects on leaf wax n -alkane traits and carbon isotopic composition from phylogeny and precipitation
1
2018
... 在纯C3 植物或C4 植物占比非常少的环境中,例如森林和淡水沼泽,δ13 Calk 变化通常被解释为相对湿度变化[40 ,41 ] .其中的机制是,大气湿度调控叶片气孔的开闭程度,进而影响到大气CO2 向叶片内部的扩散速率.一般来讲,在湿润条件下,如热带雨林,光合作用过程中碳同位素的分馏系数受CO2 扩散过程的影响较弱,因而Rubisco酶的分馏效应起到主导作用,合成的有机质表现出更偏负的碳同位素值.前期研究显示,叶片光合作用碳同位素分馏值(Δleaf )与降雨量之间存在显著的相关性[42 ] .在一项新近的美国亚热带Lake Tulane末次冰期古生态研究中,Arnold等[43 ] 基于湖泊沉积物中长链烷烃δ13 C值估算了Δleaf 值.结果发现,在Heinrich事件2~4时期,Δleaf 处于高值,而长链烷烃δD值处于低值,揭示出了末次冰期冷事件时的冷湿的气候特征.除了相对湿度,δ13 Calk 值还受CO2 浓度及其碳同位素组成、温度、植物生活型等多种因素的影响[33 ,44 ] . ...
Early degradation of plant alkanes in soils: A litterbag experiment using 13 C-labelled leaves
1
2011
... 在解译湖沼沉积物中δ13 Calk 信号时,还需要考虑从枯枝落叶堆积到被流水搬运等过程中的微生物降解及潜在的微生物输入[33 ,45 ,46 ,47 ] .由于微生物偏向于利用轻碳,有机质在经过微生物降解作用后,某些易被降解的脂类单体碳同位素组成可能会发生偏正.此外,在搬运过程中微生物自身产生的有机质输入也可能会对单体碳同位素的解译产生影响,特别是一些自养微生物产生的脂类可能具有比较偏负的碳同位素组成.气溶胶传输也可能是湖泊沉积物中保存的陆生叶蜡烷烃的一种潜在输入途径[48 ,49 ] . ...
Microbial production of long-chain n -alkanes: Implication for interpreting sedimentary leaf wax signals
1
2018
... 在解译湖沼沉积物中δ13 Calk 信号时,还需要考虑从枯枝落叶堆积到被流水搬运等过程中的微生物降解及潜在的微生物输入[33 ,45 ,46 ,47 ] .由于微生物偏向于利用轻碳,有机质在经过微生物降解作用后,某些易被降解的脂类单体碳同位素组成可能会发生偏正.此外,在搬运过程中微生物自身产生的有机质输入也可能会对单体碳同位素的解译产生影响,特别是一些自养微生物产生的脂类可能具有比较偏负的碳同位素组成.气溶胶传输也可能是湖泊沉积物中保存的陆生叶蜡烷烃的一种潜在输入途径[48 ,49 ] . ...
Chemical and carbon isotopic dynamics of grass organic matter during litter decompositions: A litterbag experiment
1
2014
... 在解译湖沼沉积物中δ13 Calk 信号时,还需要考虑从枯枝落叶堆积到被流水搬运等过程中的微生物降解及潜在的微生物输入[33 ,45 ,46 ,47 ] .由于微生物偏向于利用轻碳,有机质在经过微生物降解作用后,某些易被降解的脂类单体碳同位素组成可能会发生偏正.此外,在搬运过程中微生物自身产生的有机质输入也可能会对单体碳同位素的解译产生影响,特别是一些自养微生物产生的脂类可能具有比较偏负的碳同位素组成.气溶胶传输也可能是湖泊沉积物中保存的陆生叶蜡烷烃的一种潜在输入途径[48 ,49 ] . ...
Sources and abundances of leaf waxes in aerosols in central Europe
1
2017
... 在解译湖沼沉积物中δ13 Calk 信号时,还需要考虑从枯枝落叶堆积到被流水搬运等过程中的微生物降解及潜在的微生物输入[33 ,45 ,46 ,47 ] .由于微生物偏向于利用轻碳,有机质在经过微生物降解作用后,某些易被降解的脂类单体碳同位素组成可能会发生偏正.此外,在搬运过程中微生物自身产生的有机质输入也可能会对单体碳同位素的解译产生影响,特别是一些自养微生物产生的脂类可能具有比较偏负的碳同位素组成.气溶胶传输也可能是湖泊沉积物中保存的陆生叶蜡烷烃的一种潜在输入途径[48 ,49 ] . ...
Rapid atmospheric transport and large-scale deposition of recently synthesized plant waxes
1
2018
... 在解译湖沼沉积物中δ13 Calk 信号时,还需要考虑从枯枝落叶堆积到被流水搬运等过程中的微生物降解及潜在的微生物输入[33 ,45 ,46 ,47 ] .由于微生物偏向于利用轻碳,有机质在经过微生物降解作用后,某些易被降解的脂类单体碳同位素组成可能会发生偏正.此外,在搬运过程中微生物自身产生的有机质输入也可能会对单体碳同位素的解译产生影响,特别是一些自养微生物产生的脂类可能具有比较偏负的碳同位素组成.气溶胶传输也可能是湖泊沉积物中保存的陆生叶蜡烷烃的一种潜在输入途径[48 ,49 ] . ...
Carbon isotope composition of long chain leaf wax n -alkanes in lake sediments: A dual indicator of paleoenvironment in the Qinghai-Tibet Plateau
2
2015
... 对于湖泊等沉积场所中长链正构脂类的研究,还需要注意原地生物贡献的影响.通过对青海湖水生植物和表层沉积物中长链脂类的研究,Liu等[50 ] 提出水生的藻类和沉水植物也能贡献长链正构脂类,这些水生植物产生的长链正构烷烃的δ13 Calk 值非常接近陆地C4 植物的特征,因而会对评估流域内C4 植物的比率产生干扰.值得关注的是,湖泊内源贡献的长链脂类主要是C29 及以下正构烷烃、C30 及以下正构脂肪酸,而C31 及以上碳链的正构烷烃主要是流域内陆生植物的贡献,因此,作者提出C31 正构烷烃的δ13 Calk 值可以更准确地反映陆生植物对气候环境变化的响应[50 ] .此外,在一些湖泊中,水生生物还有可能利用来自甲烷的碳,使得合成的正构烷烃δ13 Calk 值显著偏负[40 ] .因此,对于湖沼沉积序列,在解释古气候和古环境变化时,要充分考虑沉积环境的多样性,具体问题具体分析. ...
... [50 ].此外,在一些湖泊中,水生生物还有可能利用来自甲烷的碳,使得合成的正构烷烃δ13 Calk 值显著偏负[40 ] .因此,对于湖沼沉积序列,在解释古气候和古环境变化时,要充分考虑沉积环境的多样性,具体问题具体分析. ...
n -Alkane distribution in ombrotrophic mires as indicators of vegetation change related to climatic variation
1
2000
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
Paleohydrologic reconstruction based on n -alkane distributions in ombrotrophic peat
1
2006
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
Methanotrophic symbionts provide carbon for photosynthesis in peat bogs
2
2005
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
Quantitative assessment of precipitation seasonality and summer surface wetness using ombrotrophic sediments from an Arctic Norwegian peatland
2
2009
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
... [54 ,55 ]. ...
A novel framework for quantifying past methane recycling by Sphagnum -methanotroph symbiosis using carbon and hydrogen isotope ratios of leaf wax biomarkers
2
2014
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
... ,55 ]. ...
Effect of different wetness conditions on Sphagnum lipid composition in the Erxianyan peatland,central China
1
2012
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
Hydrologic influence on the δ13 C variation in long chain n -alkanes in the Dajiuhu peatland,central China
1
2014
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
Global prevalence of methane oxidation by symbiotic bacteria in peat-moss ecosystems
1
2010
... 在泥炭地中,来自泥炭藓的脂类的碳同位素组成可以成为研究泥炭藓与微生物共生活动的有力助手.泥炭藓是主要的成炭植物,大多数泥炭藓种是以C23 正构烷烃为主峰,其次是C25 正构烷烃[51 ,52 ] .泥炭藓体内共生的嗜甲烷细菌能将泥炭与大气界面附近的甲烷氧化成CO2 ,可供泥炭藓进行光合作用[53 ] .由于泥炭地中生物成因的甲烷具有显著偏负的碳同位素组成(<-50‰),如果泥炭藓光合作用利用一定比率的来自甲烷的碳,将会在其脂类的碳同位素组成上留下印迹[54 ,55 ,56 ,57 ] .一些研究估计,泥炭藓光合作用固定的CO2 约为10%,甚至高达30%,来自甲烷氧化[58 ] .这种认识也被用于重建全新世泥炭藓泥炭地中甲烷氧化活动的变化[54 ,55 ] . ...
Boreal bog Sphagnum refixes soil-produced and respired 14 CO2
1
1999
... 在大多数环境中,高等植物都利用大气中的CO2 进行光合作用.在一些富碳环境中,例如,泥炭地,水位降低会造成大量的有机碳暴露在大气中,暴露的有机质会被分解为CO2 ,进而排放到大气中.这些分解产生的CO2 在土—气界面传输过程中,有可能被地表植物利用来进行光合作用.这种老碳利用的方式并未得到重视[59 ,60 ] .在神农架大九湖开展的一项研究显示,该泥炭地在中全新世遭遇了显著的干旱事件[61 ] .对应于干旱事件,来自高等植物的长链正构烷烃的碳同位素组成并没有表现出相应的正偏,而是出现了显著的负偏.负偏的幅度随着干旱次数的增加而变大(图1 )[62 ] .这种严重干旱时发生的δ13 Calk 值显著负偏的现象,有可能是因为植物利用了再循环的CO2 ,其δ13 C值比大气CO2 更偏负.这种作用机制在一定程度上减缓了因泥炭有机质分解向大气排放的CO2 量.同时,这项研究透露出了叶蜡脂类单体碳同位素在揭示生物地球化学过程对气候变化响应方面的研究潜力. ...
Substrate-derived CO2 is important in the development of Sphagnum spp
1
2001
... 在大多数环境中,高等植物都利用大气中的CO2 进行光合作用.在一些富碳环境中,例如,泥炭地,水位降低会造成大量的有机碳暴露在大气中,暴露的有机质会被分解为CO2 ,进而排放到大气中.这些分解产生的CO2 在土—气界面传输过程中,有可能被地表植物利用来进行光合作用.这种老碳利用的方式并未得到重视[59 ,60 ] .在神农架大九湖开展的一项研究显示,该泥炭地在中全新世遭遇了显著的干旱事件[61 ] .对应于干旱事件,来自高等植物的长链正构烷烃的碳同位素组成并没有表现出相应的正偏,而是出现了显著的负偏.负偏的幅度随着干旱次数的增加而变大(图1 )[62 ] .这种严重干旱时发生的δ13 Calk 值显著负偏的现象,有可能是因为植物利用了再循环的CO2 ,其δ13 C值比大气CO2 更偏负.这种作用机制在一定程度上减缓了因泥炭有机质分解向大气排放的CO2 量.同时,这项研究透露出了叶蜡脂类单体碳同位素在揭示生物地球化学过程对气候变化响应方面的研究潜力. ...
Concordant monsoon-driven postglacial hydrological changes in peat and stalagmite records and their impacts on prehistoric cultures in central China
4
2013
... 在大多数环境中,高等植物都利用大气中的CO2 进行光合作用.在一些富碳环境中,例如,泥炭地,水位降低会造成大量的有机碳暴露在大气中,暴露的有机质会被分解为CO2 ,进而排放到大气中.这些分解产生的CO2 在土—气界面传输过程中,有可能被地表植物利用来进行光合作用.这种老碳利用的方式并未得到重视[59 ,60 ] .在神农架大九湖开展的一项研究显示,该泥炭地在中全新世遭遇了显著的干旱事件[61 ] .对应于干旱事件,来自高等植物的长链正构烷烃的碳同位素组成并没有表现出相应的正偏,而是出现了显著的负偏.负偏的幅度随着干旱次数的增加而变大(图1 )[62 ] .这种严重干旱时发生的δ13 Calk 值显著负偏的现象,有可能是因为植物利用了再循环的CO2 ,其δ13 C值比大气CO2 更偏负.这种作用机制在一定程度上减缓了因泥炭有机质分解向大气排放的CO2 量.同时,这项研究透露出了叶蜡脂类单体碳同位素在揭示生物地球化学过程对气候变化响应方面的研究潜力. ...
... (a)大九湖ZK-5泥炭柱C29 正构烷烃δ2 H[62 ] ; (b) 大九湖ZK-3泥炭柱藿类通量[61 ] ; (c)和尚洞HS-4石笋磁学参数[63 ] ; (d) 大九湖ZK-5泥炭柱C29 正构烷烃δ13 C [62 ] ...
... (a) δ2 H of n -C29 alkane in ZK-5 peat core from Dajiuhu peatland[62 ] ; (b) Hopanoid flux in ZK-3 peat core from Dajiuhu peatland[61 ] ; ...
... 藿类广泛地分布于泥炭和湖泊沉积物中.在酸性泥炭地中,C31 αβ构型藿烷是含量最高的藿烷化合物,在一些样品中,其含量超过了正构烷烃主峰碳的含量[110 ] .基于对大九湖泥炭地藿烷环化酶基因[111 ] 以及现代环境中藿类总量与水位关系的认识,Xie等[61 ] 提出泥炭沉积中藿类主要由好氧细菌合成,藿类化合物的沉积通量可以用来反演泥炭地水位的变化.即在高水位条件下,泥炭表层处于淹水状态,好氧细菌活动受到抑制,产生的藿类总量非常低;相反,在低水位条件下,泥炭表层处于通氧状态,适合好氧细菌的繁殖,能够产生较高含量的藿类. ...
Response of carbon cycle to drier conditions in the Mid-Holocene in central China
10
2018
... 在大多数环境中,高等植物都利用大气中的CO2 进行光合作用.在一些富碳环境中,例如,泥炭地,水位降低会造成大量的有机碳暴露在大气中,暴露的有机质会被分解为CO2 ,进而排放到大气中.这些分解产生的CO2 在土—气界面传输过程中,有可能被地表植物利用来进行光合作用.这种老碳利用的方式并未得到重视[59 ,60 ] .在神农架大九湖开展的一项研究显示,该泥炭地在中全新世遭遇了显著的干旱事件[61 ] .对应于干旱事件,来自高等植物的长链正构烷烃的碳同位素组成并没有表现出相应的正偏,而是出现了显著的负偏.负偏的幅度随着干旱次数的增加而变大(图1 )[62 ] .这种严重干旱时发生的δ13 Calk 值显著负偏的现象,有可能是因为植物利用了再循环的CO2 ,其δ13 C值比大气CO2 更偏负.这种作用机制在一定程度上减缓了因泥炭有机质分解向大气排放的CO2 量.同时,这项研究透露出了叶蜡脂类单体碳同位素在揭示生物地球化学过程对气候变化响应方面的研究潜力. ...
... (a)大九湖ZK-5泥炭柱C29 正构烷烃δ2 H[62 ] ; (b) 大九湖ZK-3泥炭柱藿类通量[61 ] ; (c)和尚洞HS-4石笋磁学参数[63 ] ; (d) 大九湖ZK-5泥炭柱C29 正构烷烃δ13 C [62 ] ...
... [62 ] ...
... (a) δ2 H of n -C29 alkane in ZK-5 peat core from Dajiuhu peatland[62 ] ; (b) Hopanoid flux in ZK-3 peat core from Dajiuhu peatland[61 ] ; ...
... (c)Magnetic parameter in HS-4 stalagmite from Heshang cave[63 ] ; (d) δ13 C of n -C29 alkane in ZK-5 peat core from Dajiuhu peatland[62 ] ...
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
... [62 ].不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
... [62 ].在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ
13 C值普遍显著偏负
[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ
13 C值最负可达-62.7‰
[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联.
图2 泥炭沉积中生物体、糖类和脂类<strong>δ<sup>13</sup>C</strong>分布示意图(据参考文献[<xref ref-type="bibr" rid="R88">88</xref>]修改) <strong>Schematic diagram showing the relationship between organic carbon constituents of plants and heterotrophic bacteria in peat deposits </strong>(<strong>modified after reference</strong>[<xref ref-type="bibr" rid="R88">88</xref>]) Fig. 2 ![]()
图3 神农架大九湖<strong>ZK-5</strong>泥炭柱中藿烷和<strong>C<sub>29</sub></strong>正构烷烃<strong>δ<sup>13</sup>C</strong>序列对比<sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup> <strong>Comparisons of δ<sup>13</sup>C records in the ZK-5 peat core retrieved from the Dajiuhu peatland</strong>,<strong> Shennongjia</strong><sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup> (a) C29 ββ藿烷; (b)C31 ββ藿烷; (c)C31 αβ藿烷; (d) C29 正构烷烃 ...
... [
62 ]
(a) C29 ββ藿烷; (b)C31 ββ藿烷; (c)C31 αβ藿烷; (d) C29 正构烷烃 ...
Holocene ENSO-related cyclic storms recorded by magnetic minerals in speleothems of central China
2
2017
... (a)大九湖ZK-5泥炭柱C29 正构烷烃δ2 H[62 ] ; (b) 大九湖ZK-3泥炭柱藿类通量[61 ] ; (c)和尚洞HS-4石笋磁学参数[63 ] ; (d) 大九湖ZK-5泥炭柱C29 正构烷烃δ13 C [62 ] ...
... (c)Magnetic parameter in HS-4 stalagmite from Heshang cave[63 ] ; (d) δ13 C of n -C29 alkane in ZK-5 peat core from Dajiuhu peatland[62 ] ...
Hydrogen isotopes of n -alkanes and n -alkanoic acids as tracers of precipitation in a temperate forest and implications for paleorecords
1
2017
... 相对于长链正构烷烃,长链正构脂肪酸(δ13 Cfa )和脂肪醇的单体碳同位素组成研究很少,尤其是脂肪醇单体碳同位素组成.从叶蜡中脂类组成来看,叶片中往往含有更高含量的长链正构烷烃[64 ] .Chikaraishi[34 ] 汇总了目前已经发表的陆生C3 植物中各种叶蜡脂类的δ13 C值分布范围.在沉积序列研究中,一些研究组专注于δ13 Cfa ,而另一些研究组侧重于δ13 Calk .在同一套沉积样品中,δ13 Cfa 和δ13 Calk 的响应是否一致,这是值得关注的问题,目前这方面的对比研究还十分欠缺.这涉及到不同脂类碳同位素序列的对比问题,也可能从不同脂类碳同位素的差异上挖掘出更多的环境或生态信息. ...
Isotopic composition and probable origin of individual fatty acids in modern sediments from Ruoergai Marsh and Nansha Sea,China
2
1997
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
... [65 ,67 ].同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
Carbon isotopic studies of individual lipids in organisms from the Nansha sea area,China
2
2004
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
... [66 ].与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
沼泽沉积环境中植物和沉积脂类单体碳同位素组成特征及其成因关系研究
2
2004
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
... [66 ].与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
Compound-specific isotopic analyses:anovel tool for reconstruction of ancient biogeochemical processes
1
1990
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
Carbon isotopic variability in monosaccharides and lipids of aquatic algae and terrestrial plants
1
2002
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
The disparity of chain length distribution patterns and carbon isotopic compositions between different fatty acid purification procedures
1
2013
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
两种纯化方法获得脂肪酸的链长及碳同位素分布特征对比
1
2013
... 在湖沼相沉积环境中,脂肪酸单体碳同位素组成可以在一定程度上反映有机质的来源.例如,在若尔盖沼泽的几项研究中,发现泥炭沉积物中C16 -C28 脂肪酸的δ13 C平均值(-33.7‰)与草本和木本植物中脂肪酸的δ13 C平均值十分接近,表明泥炭中的这些脂肪酸主要来源于高等植物[65 ,66 ] ;而泥炭沉积物中的C14 和C15 脂肪酸具有明显偏负的碳同位素组成,分别为-38.7‰和-37.2‰,表明这些短链脂肪酸可能来源于化能自养细菌[65 ,67 ] .同时,还发现若尔盖沼泽中草本植物(-33.4‰)的脂肪酸δ13 C较木本植物(-32.5‰)略微偏负,与之相似的是草本植物的正构烷烃δ13 C较木本植物也偏负,这可能是由于不同植物在生长过程中碳同位素分馏的差异所致,也可能是由于草本植物和木本植物的生长高度不同,草本植物紧贴地面生长导致吸收更多偏负的CO2 [66 ] .与之相似,von Dongen等[68 ] 发现荷兰Bargerveen泥炭地中的狭叶泥炭藓Sphagnum cuspidatum 的C16 脂肪酸具有较为偏负的碳同位素组成(-35.0‰),而同样生长在泥炭地中的杜鹃花科植物Erica tetralix 根中的C16 脂肪酸δ13 C略微偏正(-28.8‰).在一项采用2种不同纯化方法对比植物和泥炭脂肪酸δ13 C的研究中,发现C4 植物狗尾草(Setaira viridis )的C16 脂肪酸δ13 C比C3 植物三叶草(Trifolium repens )明显偏正[69 ] ,这与C3 和C4 植物叶蜡正构烷烃δ13 C的差异类似.总体上,在沼泽环境中不同植被来源的脂肪酸δ13 C的大致变化规律为:泥炭藓<C3 草本植物<C3 木本植物<C4 植物,而一些来源于自养细菌的脂肪酸δ13 C可能更加偏负. ...
Variability in the distribution of lipid biomarkers and their molecular isotopic composition in Altamaha estuarine sediments: Implications for the relative contribution of organic matter from various sources
1
2001
... 相对于沼泽环境,湖泊等沉积环境中脂肪酸δ13 C在反映有机质来源上可能更加复杂.例如,在美国佐治亚州Altamaha河口沉积物中脂肪酸分布及其δ13 C的研究中,发现长链脂肪酸δ13 C比较偏负,被认为主要来源于陆源输入,而短链脂肪酸δ13 C比较偏正,被认为主要来源于海洋浮游生物[70 ] .王丽芳等[71 ] 对巢湖沉积钻孔中结合态脂肪酸的组成及其单体碳同位素进行了研究,发现湖泊内源的藻类和细菌是C16 脂肪酸的主要贡献者,并且沉积柱从下到上C16 脂肪酸δ13 C值的逐渐偏正反映了湖泊的富营养化.陶舒琴等[72 ] 分析了黄河悬浮颗粒物中脂肪酸的δ13 C特征,发现长链脂肪酸(C28 ,C30 )的δ13 C变化范围为-31.1‰ ~ -32.2‰,表明其主要来源于以C3 植被覆盖为主的黄土土壤有机质,而δ13 C比较偏正的短链脂肪酸(C16 ,C18 )可能主要来源于水生藻类和异养微生物. ...
Sedimentary records of the process of eutrophication in Chaohu Lake:Evidence from bound fatty acid and stable special carbon isotope data
1
2010
... 相对于沼泽环境,湖泊等沉积环境中脂肪酸δ13 C在反映有机质来源上可能更加复杂.例如,在美国佐治亚州Altamaha河口沉积物中脂肪酸分布及其δ13 C的研究中,发现长链脂肪酸δ13 C比较偏负,被认为主要来源于陆源输入,而短链脂肪酸δ13 C比较偏正,被认为主要来源于海洋浮游生物[70 ] .王丽芳等[71 ] 对巢湖沉积钻孔中结合态脂肪酸的组成及其单体碳同位素进行了研究,发现湖泊内源的藻类和细菌是C16 脂肪酸的主要贡献者,并且沉积柱从下到上C16 脂肪酸δ13 C值的逐渐偏正反映了湖泊的富营养化.陶舒琴等[72 ] 分析了黄河悬浮颗粒物中脂肪酸的δ13 C特征,发现长链脂肪酸(C28 ,C30 )的δ13 C变化范围为-31.1‰ ~ -32.2‰,表明其主要来源于以C3 植被覆盖为主的黄土土壤有机质,而δ13 C比较偏正的短链脂肪酸(C16 ,C18 )可能主要来源于水生藻类和异养微生物. ...
巢湖富营养化的沉积记录:结合态脂肪酸及其单体碳同位素特征
1
2010
... 相对于沼泽环境,湖泊等沉积环境中脂肪酸δ13 C在反映有机质来源上可能更加复杂.例如,在美国佐治亚州Altamaha河口沉积物中脂肪酸分布及其δ13 C的研究中,发现长链脂肪酸δ13 C比较偏负,被认为主要来源于陆源输入,而短链脂肪酸δ13 C比较偏正,被认为主要来源于海洋浮游生物[70 ] .王丽芳等[71 ] 对巢湖沉积钻孔中结合态脂肪酸的组成及其单体碳同位素进行了研究,发现湖泊内源的藻类和细菌是C16 脂肪酸的主要贡献者,并且沉积柱从下到上C16 脂肪酸δ13 C值的逐渐偏正反映了湖泊的富营养化.陶舒琴等[72 ] 分析了黄河悬浮颗粒物中脂肪酸的δ13 C特征,发现长链脂肪酸(C28 ,C30 )的δ13 C变化范围为-31.1‰ ~ -32.2‰,表明其主要来源于以C3 植被覆盖为主的黄土土壤有机质,而δ13 C比较偏正的短链脂肪酸(C16 ,C18 )可能主要来源于水生藻类和异养微生物. ...
The abundance and 13 C characteristics of lipid biomarkers in surface suspended particulates from the Yellow River and their source implication
1
2015
... 相对于沼泽环境,湖泊等沉积环境中脂肪酸δ13 C在反映有机质来源上可能更加复杂.例如,在美国佐治亚州Altamaha河口沉积物中脂肪酸分布及其δ13 C的研究中,发现长链脂肪酸δ13 C比较偏负,被认为主要来源于陆源输入,而短链脂肪酸δ13 C比较偏正,被认为主要来源于海洋浮游生物[70 ] .王丽芳等[71 ] 对巢湖沉积钻孔中结合态脂肪酸的组成及其单体碳同位素进行了研究,发现湖泊内源的藻类和细菌是C16 脂肪酸的主要贡献者,并且沉积柱从下到上C16 脂肪酸δ13 C值的逐渐偏正反映了湖泊的富营养化.陶舒琴等[72 ] 分析了黄河悬浮颗粒物中脂肪酸的δ13 C特征,发现长链脂肪酸(C28 ,C30 )的δ13 C变化范围为-31.1‰ ~ -32.2‰,表明其主要来源于以C3 植被覆盖为主的黄土土壤有机质,而δ13 C比较偏正的短链脂肪酸(C16 ,C18 )可能主要来源于水生藻类和异养微生物. ...
黄河悬浮颗粒物中类脂生物标志物的组成和稳定碳同位素分布特征及来源研究
1
2015
... 相对于沼泽环境,湖泊等沉积环境中脂肪酸δ13 C在反映有机质来源上可能更加复杂.例如,在美国佐治亚州Altamaha河口沉积物中脂肪酸分布及其δ13 C的研究中,发现长链脂肪酸δ13 C比较偏负,被认为主要来源于陆源输入,而短链脂肪酸δ13 C比较偏正,被认为主要来源于海洋浮游生物[70 ] .王丽芳等[71 ] 对巢湖沉积钻孔中结合态脂肪酸的组成及其单体碳同位素进行了研究,发现湖泊内源的藻类和细菌是C16 脂肪酸的主要贡献者,并且沉积柱从下到上C16 脂肪酸δ13 C值的逐渐偏正反映了湖泊的富营养化.陶舒琴等[72 ] 分析了黄河悬浮颗粒物中脂肪酸的δ13 C特征,发现长链脂肪酸(C28 ,C30 )的δ13 C变化范围为-31.1‰ ~ -32.2‰,表明其主要来源于以C3 植被覆盖为主的黄土土壤有机质,而δ13 C比较偏正的短链脂肪酸(C16 ,C18 )可能主要来源于水生藻类和异养微生物. ...
Abrupt tropical vegetation response to rapid climate changes
1
2004
... 除了反映有机质来源,脂肪酸单体碳同位素组成在反演古植被与古气候变化中也具有一定的潜力.例如,Hughen等[73 ] 利用南美洲北部Cariaco盆地沉积物中的长链脂肪酸δ13 C和平均碳链长度(Average Chain Length,ACL),重建了该地区末次冰消期的古植被变化,发现北大西洋热带地区和高纬度地区的气候变化是同步的,且热带植被变化响应气候变化具有几十年的滞后.在最近的一项关于日本富士山东北部Yamanaka湖泊沉积物叶蜡长链脂肪酸分布和单体碳同位素组成的研究中,Yamamoto等[74 ] 发现15 000年以来C30 脂肪酸δ13 C的偏负对应于脂肪酸ACL的减小,表明植物在湿润条件下对可利用水增多时的生理反应,这与湿润条件下植物叶蜡正构烷烃δ13 C的偏负类似;同时全新世的C30 脂肪酸δ13 C显示出与董哥洞石笋δ18 O类似的波动趋势,指示了这一地区的水文气候的变化与季风活动有关. ...
Volcanic and environmental influences of Mt. Fuji on the δ13 C of terrestrially-derived n -alkanoic acids in sediment from Lake Yamanaka,central Japan
1
2018
... 除了反映有机质来源,脂肪酸单体碳同位素组成在反演古植被与古气候变化中也具有一定的潜力.例如,Hughen等[73 ] 利用南美洲北部Cariaco盆地沉积物中的长链脂肪酸δ13 C和平均碳链长度(Average Chain Length,ACL),重建了该地区末次冰消期的古植被变化,发现北大西洋热带地区和高纬度地区的气候变化是同步的,且热带植被变化响应气候变化具有几十年的滞后.在最近的一项关于日本富士山东北部Yamanaka湖泊沉积物叶蜡长链脂肪酸分布和单体碳同位素组成的研究中,Yamamoto等[74 ] 发现15 000年以来C30 脂肪酸δ13 C的偏负对应于脂肪酸ACL的减小,表明植物在湿润条件下对可利用水增多时的生理反应,这与湿润条件下植物叶蜡正构烷烃δ13 C的偏负类似;同时全新世的C30 脂肪酸δ13 C显示出与董哥洞石笋δ18 O类似的波动趋势,指示了这一地区的水文气候的变化与季风活动有关. ...
Sesqui-,di-,and triterpenoids as chemosystematic markers in Extant conifers—A review
1
2001
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
Higher plant terpenoids: A phytocentric overview of their ecological roles
1
1994
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
A phylogenetic analysis of conifer diterpenoids and their carbon isotopes for chemotaxonomic applications
1
2019
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
Contrasted distributions of triterpene derivatives in the sediments of Lake Ca?ó reflect paleoenvironmental changes during the last 20,000yrs in NE Brazil
1
2007
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
Distribution of aliphatic des-A-triterpenoids in the Dajiuhu peat deposit,Southern China
1
2008
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
Paleoclimate influence on early diagenesis of plant triterpenes in the Dajiuhu peatland,central China
1
2013
... 除了长链正构脂类,被子植物中通常含有较高含量的三萜类化合物,主要有奥利烷、乌散烷和羽扇烷等3个系列[75 ] .这些三萜类化合物被认为是植物体抵抗病虫害的一种方式[76 ] .在沉积物中,三萜类化合物可以被用来指示被子植物的输入,以区别来自裸子植物的二萜类化合物[31 ,77 ] .在成岩作用过程中,三萜类化合物会受到微生物改造,转变为各种三萜烯.沉积环境的相关信息,特别是与水文相关的氧化还原状况,会被三萜烯的组成记录下来,成为古环境重建的潜在指标[78 ,79 ,80 ] . ...
Des -A-lupane in an East African lake sedimentaryrecord as a new proxy for the stable carbon isotopic composition of C3 plants
2
2016
... 三萜类和叶蜡脂类都来自高等植物,在响应气候变化方面,它们的碳同位素组成是表现出相似还是差异的响应是值得关注的科学问题.不过,相关的研究还非常少.一项在非洲东部Lake Challa开展的研究显示,从末次冰盛期以来,湖泊沉积柱中C31 正构烷烃和脱A-环羽扇烷(羽扇烷的一种降解产物)的碳同位素表现出了非常不一样的变化趋势[81 ] .后者的变化非常平缓,而前者则表现出了明显的变化,在末次冰消期C31 正构烷烃的碳同位素组成相对偏正,到了全新世,2种脂类的碳同位素组成的变化则非常相似.对于这种情况,作者解释为脱A-环羽扇烷主要来自C3 植物,而C31 正构烷烃同时来自C3 和C4 植物.基于此,作者提出可以将主要来自C3 植物的脱A-环羽扇烷的δ13 C值作为对应时段的C3 植物δ13 C值的端元值,由此可以更为准确地估算流域内C4 植物的比率[81 ] .这其中需要考虑合成途径差异而造成的正构烷烃和三萜类碳同位素的差值[31 ,82 ] .由此可见,综合多种脂类的碳同位素组成能够更准确地揭示古生态和古环境的变化,还有可能发掘出更多的环境信息. ...
... [81 ].这其中需要考虑合成途径差异而造成的正构烷烃和三萜类碳同位素的差值[31 ,82 ] .由此可见,综合多种脂类的碳同位素组成能够更准确地揭示古生态和古环境的变化,还有可能发掘出更多的环境信息. ...
Fractionation of carbon and hydrogen isotopes in biosynthetic processes
2
2001
... 三萜类和叶蜡脂类都来自高等植物,在响应气候变化方面,它们的碳同位素组成是表现出相似还是差异的响应是值得关注的科学问题.不过,相关的研究还非常少.一项在非洲东部Lake Challa开展的研究显示,从末次冰盛期以来,湖泊沉积柱中C31 正构烷烃和脱A-环羽扇烷(羽扇烷的一种降解产物)的碳同位素表现出了非常不一样的变化趋势[81 ] .后者的变化非常平缓,而前者则表现出了明显的变化,在末次冰消期C31 正构烷烃的碳同位素组成相对偏正,到了全新世,2种脂类的碳同位素组成的变化则非常相似.对于这种情况,作者解释为脱A-环羽扇烷主要来自C3 植物,而C31 正构烷烃同时来自C3 和C4 植物.基于此,作者提出可以将主要来自C3 植物的脱A-环羽扇烷的δ13 C值作为对应时段的C3 植物δ13 C值的端元值,由此可以更为准确地估算流域内C4 植物的比率[81 ] .这其中需要考虑合成途径差异而造成的正构烷烃和三萜类碳同位素的差值[31 ,82 ] .由此可见,综合多种脂类的碳同位素组成能够更准确地揭示古生态和古环境的变化,还有可能发掘出更多的环境信息. ...
... 对于自养微生物,其脂类δ13 C值主要受控于无机碳源的δ13 C值与供应量、固碳途径和微生物的生长阶段[88 ] .异养微生物主要利用已有的有机物进行自身的生长.传统上认为,异养微生物生物量的δ13 C值非常接近于其摄取的有机物的δ13 C值[82 ] . ...
The Paleocene-Eocene carbon isotope excursion in higher plant organic matter:Differential fractionation of angiosperms and conifers in the Arctic
2
2007
... 结合三萜类和长链正构烷烃的碳同位素组成,还被用来更好地认识古新世和始新世之间极热事件(Paleocene-Eocene Thermal Maximum,PETM)不同碳组分碳同位素负偏移程度差异的原因[83 ] .三萜类主要来自被子植物,而长链正构烷烃有被子植物和裸子植物的贡献.研究者还利用三萜类的碳同位素组成来探讨植物光合作用的碳同位素分馏系数是否在PETM时期表现出显著的特异性[84 ] . ...
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
Paleogene plants fractionated carbon isotopes similar to modern plants
1
2015
... 结合三萜类和长链正构烷烃的碳同位素组成,还被用来更好地认识古新世和始新世之间极热事件(Paleocene-Eocene Thermal Maximum,PETM)不同碳组分碳同位素负偏移程度差异的原因[83 ] .三萜类主要来自被子植物,而长链正构烷烃有被子植物和裸子植物的贡献.研究者还利用三萜类的碳同位素组成来探讨植物光合作用的碳同位素分馏系数是否在PETM时期表现出显著的特异性[84 ] . ...
Principles,methodology and application of molecular stratigraphy
1
2007
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
分子地层学的原理、方法及应用实例
1
2007
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
A review of molecular organic proxies for examining modern and ancient lacustrine environments
1
2011
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
The organic geochemistry of glycerol dialkyl glycerol tetraether lipids:A review
1
2013
... 微生物是陆地生态系统的重要组成部分.在改造植物有机质的同时,微生物输入自身合成的脂类.在微生物体中,脂类主要分布在细胞膜中,包括磷脂、GDGTs和藿类等.一方面,某些脂类只来源于特定的微生物类别或代谢途径,因此可以直接用来指示物源或代谢过程[2 ,6 ,85 ] .另一方面,得益于微生物活动对环境变化的灵敏响应,基于微生物的脂类指标已经成为湖沼沉积古环境和古生态重建的重要工具[5 ,86 ,87 ] . ...
Carbon isotopic compositions of prokaryotic lipids as tracers of carbon cycling in diverse settings
4
2003
... 对于自养微生物,其脂类δ13 C值主要受控于无机碳源的δ13 C值与供应量、固碳途径和微生物的生长阶段[88 ] .异养微生物主要利用已有的有机物进行自身的生长.传统上认为,异养微生物生物量的δ13 C值非常接近于其摄取的有机物的δ13 C值[82 ] . ...
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ
13 C值普遍显著偏负
[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ
13 C值最负可达-62.7‰
[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联.
图2 泥炭沉积中生物体、糖类和脂类<strong>δ<sup>13</sup>C</strong>分布示意图(据参考文献[<xref ref-type="bibr" rid="R88">88</xref>]修改) <strong>Schematic diagram showing the relationship between organic carbon constituents of plants and heterotrophic bacteria in peat deposits </strong>(<strong>modified after reference</strong>[<xref ref-type="bibr" rid="R88">88</xref>]) Fig. 2 ![]()
图3 神农架大九湖<strong>ZK-5</strong>泥炭柱中藿烷和<strong>C<sub>29</sub></strong>正构烷烃<strong>δ<sup>13</sup>C</strong>序列对比<sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup> <strong>Comparisons of δ<sup>13</sup>C records in the ZK-5 peat core retrieved from the Dajiuhu peatland</strong>,<strong> Shennongjia</strong><sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup> (a) C29 ββ藿烷; (b)C31 ββ藿烷; (c)C31 αβ藿烷; (d) C29 正构烷烃 ...
... [
88 ])
Fig. 2 ![]()
图3 神农架大九湖<strong>ZK-5</strong>泥炭柱中藿烷和<strong>C<sub>29</sub></strong>正构烷烃<strong>δ<sup>13</sup>C</strong>序列对比<sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup> <strong>Comparisons of δ<sup>13</sup>C records in the ZK-5 peat core retrieved from the Dajiuhu peatland</strong>,<strong> Shennongjia</strong><sup>[<xref ref-type="bibr" rid="R62">62</xref>]</sup> (a) C29 ββ藿烷; (b)C31 ββ藿烷; (c)C31 αβ藿烷; (d) C29 正构烷烃 ...
... (2)在关注单个化合物的碳同位素组成的基础上,可以考虑联合多种脂类的碳同位素值,利用它们指示的不同生态系统成员的优势,从更多的侧面去认识生态系统对气候变化的响应过程和机制.这方面已有一些探索性的研究,例如非洲湖泊中长链正构烷烃和脱A-环羽扇烷单体碳的对比研究.在泥炭沉积中,此方面尤其值得重视.泥炭有机质含量非常高,包含有来自高等植物的各种脂类和来自微生物的多种脂类,可以从碳同位素角度深入地刻画泥炭地生态系统对气候变化的生态响应[88 ] . ...
The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil
1
1996
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Lipid analysis in microbial ecology
1
1989
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Methods for assessing the composition and diversity of soil microbial communities
1
2000
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques
1
2003
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Microbial carbon cycling in oligotrophic regional aquifers near the Tono Uranium Mine,Japan as inferred from δ13 C and Δ14 C values of in situ phospholipid fatty acids and carbon sources
2
2010
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
... [93 ]研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Stable isotope techniques of PLFAs and its application in soil microbiology
1
2008
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
PLFAs稳定同位素技术及其在土壤微生物学中的应用
1
2008
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Application of 13 C-labeled PLFA analysis in soil microbial ecology studies
1
2016
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
13 C标记磷脂脂肪酸分析在土壤微生物生态研究中的应用
1
2016
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Microbial phospholipid biomarkers and stable isotope methods help reveal soil functions
1
2015
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Carbon isotope fractionation in phospholipid fatty acid biomarkers of bacteria and fungi native to an acid mine drainage lake
1
2009
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Autotrophic and heterotrophic associated biosignatures in modern freshwater microbialites over seasonal and spatial gradients
1
2014
... PLFAs技术已经被广泛应用于各种环境中活体微生物生物量的定量测量,反映微生物群落结构的变化等[89 ,90 ,91 ,92 ] .在此基础上,PLFAs单体碳同位素(δ13 CPLFA )可以反映在原位生长的微生物碳同位素组成的细节,提供微生物利用的碳和能源的信息[93 ] ,深入揭示微生物参与的有机质代谢过程,将特定的微生物种群与土壤过程联系起来[94 ,95 ,96 ] .Cowie等[97 ] 对加拿大安大略省北部酸性矿坑水中的微生物进行培养,利用PLFA单体碳同位素揭示了自养和异养微生物在碳源利用过程中的同位素分馏差异.Mills等[93 ] 研究了日本一处矿井沉积物和地下水中PLFAs的单体碳同位素特征,发现指示II型甲烷氧化菌的PLFA18∶ 1 ω8c的δ13 C值在沉积物和地下水中明显不同,表明甲烷氧化菌在2种环境中可能采用不同的碳同化途径,并且地下水中δ13 CPLFA 值更加偏负,指示了可能由化能自养作用主导的微生物群落.Brady等[98 ] 研究了加拿大西部Pavilion湖泊中淡水微生物的PLFA丰度和单体碳同位素组成,发现不同种类的PLFA的δ13 C与总有机碳δ13 Corg 之间的差值Δδ13 CPLFA-org 有明显差别,可以指示自养或者异养微生物过程,并且利用PLFA单体碳同位素揭示了湖泊中不同位置微生物自养和异养过程的差异. ...
Production rates of bacterial tetraether lipids and fatty acids in peatland under varying oxygen concentrations
2
2017
... 除了探究天然环境中PLFAs的单体碳同位素特征,许多研究还采用13 C标记配合微生物培养的手段研究各种碳源底物在微生物中的代谢方式.Huguet等[99 ] 对泥炭进行有氧与厌氧条件下的培养,对比了泥炭中细菌来源的脂肪酸和支链甘油二烷基甘油四醚(Branched Glycerol Dialkyl Glycerol Tetraether,bGDGT)脂类的生产率,并通过13 C和氘(2 H)同位素标记的方法,识别出2种支链PLFA与bGDGT具有相似的生产模式,为解决沉积环境中bGDGT的生物来源提供了重要信息.Veuger等[100 ] 向潮汐沉积物中加入13 C标记的葡萄糖和15 N标记的铵,探究有机质降解过程中的可水解氨基酸,单糖和脂肪酸的变化,发现磷脂脂肪酸和总脂肪酸具有相似的变化:在前期标记阶段没有13 C的损失,而在后期阶段13 C损失比较快,表明这2种来源的脂肪酸的不稳定性非常相似.同时根据13 C的富集程度识别出18∶ 1 ω7c,a15∶ 0 和i15∶ 0 3种相对抗降解的脂肪酸,而在沉积物中藻类物质的降解过程中也观察到这3种脂肪酸的产生. ...
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
Fate of microbial nitrogen,carbon,hydrolysable amino acids,monosaccharides,and fatty acids in sediment
1
2012
... 除了探究天然环境中PLFAs的单体碳同位素特征,许多研究还采用13 C标记配合微生物培养的手段研究各种碳源底物在微生物中的代谢方式.Huguet等[99 ] 对泥炭进行有氧与厌氧条件下的培养,对比了泥炭中细菌来源的脂肪酸和支链甘油二烷基甘油四醚(Branched Glycerol Dialkyl Glycerol Tetraether,bGDGT)脂类的生产率,并通过13 C和氘(2 H)同位素标记的方法,识别出2种支链PLFA与bGDGT具有相似的生产模式,为解决沉积环境中bGDGT的生物来源提供了重要信息.Veuger等[100 ] 向潮汐沉积物中加入13 C标记的葡萄糖和15 N标记的铵,探究有机质降解过程中的可水解氨基酸,单糖和脂肪酸的变化,发现磷脂脂肪酸和总脂肪酸具有相似的变化:在前期标记阶段没有13 C的损失,而在后期阶段13 C损失比较快,表明这2种来源的脂肪酸的不稳定性非常相似.同时根据13 C的富集程度识别出18∶ 1 ω7c,a15∶ 0 和i15∶ 0 3种相对抗降解的脂肪酸,而在沉积物中藻类物质的降解过程中也观察到这3种脂肪酸的产生. ...
Hopanoids
1
2010
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Hopanoids play a role in membrane integrity and pH homeostasis in Rhodopseudomonas palustris TIE-1
1
2009
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Draft genome sequence of Rhodomicrobium udaipurense JA643T with special reference to hopanoid biosynthesis
1
2014
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
2-Methylhopanoids are maximally produced in akinetes of Nostoc punctiforme :Geobiological implications
1
2009
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Phylogenetic analysis of HpnP reveals the origin of 2-methylhopanoid production in Alphaproteobacteria
1
2015
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Bacterial populations recorded in diverse sedimentary biohopanoid distributions
1
2007
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
fiy063
1
2018
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Novel hopanoid cyclases from the environment
1
2007
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Hopanoid lipids:From membranes to plant-bacteria interactions
1
2018
... 藿类主要分布在细菌的细胞膜中,充当着类似于真核生物细胞膜中甾醇的功能[101 ] .藿类化合物种类众多,前驱物主要有C35 的细菌藿四醇及衍生物、C30 的里白醇.多种类型的细菌可以合成藿类化合物,包括甲烷氧化菌、蓝细菌、固氮菌和紫色非硫细菌等[102 ,103 ,104 ,105 ,106 ,107 ] .Pearson等[108 ] 汇总了藿烷环化酶(控制藿类合成的一种关键酶)的基因数据,发现只有10%的细菌包含藿烷环化酶基因.另外还有少量的真核生物中发现了藿类化合物,如高等植物、蕨类植物、苔藓、地衣和真菌,这些真核生物中的藿类化合物大多是二级代谢产物,由共生或者寄生的微生物所产生[109 ] . ...
Environmental factors affecting the low temperature isomerization of homohopanes in acidic peat deposits,central China
1
2015
... 藿类广泛地分布于泥炭和湖泊沉积物中.在酸性泥炭地中,C31 αβ构型藿烷是含量最高的藿烷化合物,在一些样品中,其含量超过了正构烷烃主峰碳的含量[110 ] .基于对大九湖泥炭地藿烷环化酶基因[111 ] 以及现代环境中藿类总量与水位关系的认识,Xie等[61 ] 提出泥炭沉积中藿类主要由好氧细菌合成,藿类化合物的沉积通量可以用来反演泥炭地水位的变化.即在高水位条件下,泥炭表层处于淹水状态,好氧细菌活动受到抑制,产生的藿类总量非常低;相反,在低水位条件下,泥炭表层处于通氧状态,适合好氧细菌的繁殖,能够产生较高含量的藿类. ...
pH shaping the composition of sqhC -containing bacterial communities
1
2015
... 藿类广泛地分布于泥炭和湖泊沉积物中.在酸性泥炭地中,C31 αβ构型藿烷是含量最高的藿烷化合物,在一些样品中,其含量超过了正构烷烃主峰碳的含量[110 ] .基于对大九湖泥炭地藿烷环化酶基因[111 ] 以及现代环境中藿类总量与水位关系的认识,Xie等[61 ] 提出泥炭沉积中藿类主要由好氧细菌合成,藿类化合物的沉积通量可以用来反演泥炭地水位的变化.即在高水位条件下,泥炭表层处于淹水状态,好氧细菌活动受到抑制,产生的藿类总量非常低;相反,在低水位条件下,泥炭表层处于通氧状态,适合好氧细菌的繁殖,能够产生较高含量的藿类. ...
Molecular and isotopic stratigraphy in an ombrotrophic mire for paleoclimate reconstruction
1
2004
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
Increased terrestrial methane cycling at the Palaeocene-Eocene thermal maximum
1
2007
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
Holocene variations in peatland methane cycling associated with the Asian summer monsoon system
1
2014
... 研究者对泥炭沉积中藿烷的碳同位素进行了研究.结果发现,处于显著地理隔离的不同泥炭地,C31 αβ构型藿烷的δ13 C值在表层泥炭中非常相似(约-26‰),相对总有机碳同位素组成显著偏正,被认为是利用了相对富集13 C的糖类进行异养生长(图2 )[20 ,62 ,112 ] .在大九湖的ZK-5泥炭柱中,C31 αβ藿烷的δ13 C值在末次冰消期相对偏负(约2‰)[62 ] .不同于C31 αβ藿烷,C29 ββ藿烷的δ13 C值变化幅度更大,最负时接近-40‰,指示出可能有来自嗜甲烷细菌的贡献(图3 )[62 ] .在PETM时期的褐煤中,研究者观察到C29 ββ藿烷的δ13 C值发生了显著的负偏,最低值接近-80‰,负偏的幅度大于C31 ββ藿烷的δ13 C值[113 ] .泥炭藓提取的脂类中,裂解产生的C32 ββ藿酸的δ13 C值约为-39.8‰,比泥炭藓自身的有机质的碳同位素组成偏负,很可能来自内共生的嗜甲烷细菌[53 ] .从泥炭藓中提取的藿类的δ13 C值表现出随变干而增大的趋势[19 ] .在红原泥炭地全新世泥炭沉积中,里白烯δ13 C值在中全新世干旱阶段显著负偏(<-50‰)[114 ] .已有的研究充分体现出了利用藿烷单体碳同位素来研究湿地生态响应的优越性. ...
Diploptene:An indicator of terrigenous organic carbon in Washington coastal sediments
1
1992
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Stable carbon isotopic correlation of individual biolipids in aquatic organisms and a lake bottom sediment
1
1994
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Molecular and isotopic biogeochemistry of the Miocene Clarkia Formation: Hydrocarbons and alcohols
1
1995
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Identification of unusual 17 β (H)-moret-22(29)-ene in lake sediments
1
1995
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Isotopic biogeochemistry of the lipids in recent sediments of Lake Bled (Slovenia) and Baldeggersee (Switzerland)
1
2002
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Biomarker and stable carbon isotope analyses of sedimentary organic matter from Lake Tswaing:evidence for deglacial wetness and early Holocene drought from South Africa
2
2010
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
... 一项在非洲湖泊开展的研究,探讨了末次间冰期湖泊沉积物中来自葡萄球藻的葡萄球藻烯的δ13 C变化[128 ] .在湖泊沉积物中,短链异构烷烃(主要是C17 和C18 )主要来自蓝细菌[129 ] .来自蓝细菌的这些短链烷烃的δ13 C值通常表现出较大的变化范围[120 ,130 ,131 ] ,例如,在CO2 供应受限的碱性淡水水体中,来自蓝细菌的n -C17 烷的δ13 C值会显著偏正[130 ] . ...
Diploptene δ13 C values from contemporary thermokarst lake sediments show complex spatial variation
1
2016
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Biomarker and compound-specific δ13 C evidence for changing environmental conditions and carbon limitation at Lake Koucha,eastern Tibetan Plateau
1
2010
... 湖泊沉积中的藿类单体碳同位素组成也得到了一定程度的关注,不过主要是集中于表层样品中,缺乏连续的时间序列研究.在湖泊环境中,尤其是青藏高原湖泊和高纬湖泊中,里白烯δ13 C值普遍显著偏负[115 ,116 ,117 ,118 ,119 ,120 ,121 ] .例如在青藏高原寇查湖末次冰消期以来的沉积物中,里白烯δ13 C值最负可达-62.7‰[122 ] ,被认为与湖泊环境中的嗜甲烷过程有关联. ...
Coevolution between terrestrial ecosystem and Earth environment
1
2016
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
陆地生态系统与地球环境的协同演化
1
2016
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
Redox-dependent niche differentiation provides evidence for multiple bacterial sources of glycerol tetraether lipids in lakes
2
2018
... GDGTs是近些年来得到广泛关注的生物标志物,基于GDGTs建立了多种古气候指标[83 ] ,可以用来探讨陆地生态系统与地球环境的协同演化[123 ] .根据结构的差异,可分为类异戊二烯类(isoprenoid Glycerol Dialkyl Glycerol Tetraether,iGDGT)和支链GDGT(branched Glycerol Dialkyl Glycerol Tetraether,bGDGT).前者来自古菌,而后者由生源尚未确定的一类细菌产生.单体碳同位素的研究为探索bGDGT的生物来源提供了有用的线索[21 ,99 ,124 ] .根据相对偏正的δ13 C值以及在土壤剖面中的分布,bGDGT的生物源被推测为生活在厌氧条件下的异养细菌[21 ] .在一些湖泊环境中,研究者观察到了非常负偏的bGDGT δ13 C值,被认为摄取了来自甲烷氧化细菌的有机质[24 ,124 ] . ...
... ,124 ]. ...
Biomarker-based inferences of past climate:The alkenone p CO2 Proxy
1
2014
... 除了上述类别的微生物脂类,来自颗石藻的长链烯酮化合物的碳同位素也得到了广泛研究.主要集中在海相沉积物中,被用来重建新生代大气CO2 浓度的变化[125 ] .在少数的几项陆地湖泊烯酮单体碳研究中,烯酮δ13 C值表现出较为偏负的特征.例如在南极Ace Lake中,烯酮δ13 C值范围为-31.7‰~-36.1‰[126 ] ;而在格陵兰湖泊中,烯酮δ13 C值约为-40‰[127 ] .这种较负的碳同位素特征,被解释为较低的湖水温度下碳同位素分馏系数增大,或者是利用了来自有机质分解产生的CO2 . ...
Molecular organic tracers of biogeochemical processes in a saline meromictic lake (Ace Lake)
1
2001
... 除了上述类别的微生物脂类,来自颗石藻的长链烯酮化合物的碳同位素也得到了广泛研究.主要集中在海相沉积物中,被用来重建新生代大气CO2 浓度的变化[125 ] .在少数的几项陆地湖泊烯酮单体碳研究中,烯酮δ13 C值表现出较为偏负的特征.例如在南极Ace Lake中,烯酮δ13 C值范围为-31.7‰~-36.1‰[126 ] ;而在格陵兰湖泊中,烯酮δ13 C值约为-40‰[127 ] .这种较负的碳同位素特征,被解释为较低的湖水温度下碳同位素分馏系数增大,或者是利用了来自有机质分解产生的CO2 . ...
Long-chain alkenones in Greenland lake sediments:Low δ13 C values and exceptional abundance
1
2005
... 除了上述类别的微生物脂类,来自颗石藻的长链烯酮化合物的碳同位素也得到了广泛研究.主要集中在海相沉积物中,被用来重建新生代大气CO2 浓度的变化[125 ] .在少数的几项陆地湖泊烯酮单体碳研究中,烯酮δ13 C值表现出较为偏负的特征.例如在南极Ace Lake中,烯酮δ13 C值范围为-31.7‰~-36.1‰[126 ] ;而在格陵兰湖泊中,烯酮δ13 C值约为-40‰[127 ] .这种较负的碳同位素特征,被解释为较低的湖水温度下碳同位素分馏系数增大,或者是利用了来自有机质分解产生的CO2 . ...
Contrasting variations in the structure and stable carbon isotopic composition of botryococcenes through the last Glacial-Interglacial transition in Lake Masoko (southern Tanzania)
1
2012
... 一项在非洲湖泊开展的研究,探讨了末次间冰期湖泊沉积物中来自葡萄球藻的葡萄球藻烯的δ13 C变化[128 ] .在湖泊沉积物中,短链异构烷烃(主要是C17 和C18 )主要来自蓝细菌[129 ] .来自蓝细菌的这些短链烷烃的δ13 C值通常表现出较大的变化范围[120 ,130 ,131 ] ,例如,在CO2 供应受限的碱性淡水水体中,来自蓝细菌的n -C17 烷的δ13 C值会显著偏正[130 ] . ...
Lipid biomarkers and their stable carbon isotopes in oxic and anoxic sediments of Lake Bled (NW Slovenia)
1
2017
... 一项在非洲湖泊开展的研究,探讨了末次间冰期湖泊沉积物中来自葡萄球藻的葡萄球藻烯的δ13 C变化[128 ] .在湖泊沉积物中,短链异构烷烃(主要是C17 和C18 )主要来自蓝细菌[129 ] .来自蓝细菌的这些短链烷烃的δ13 C值通常表现出较大的变化范围[120 ,130 ,131 ] ,例如,在CO2 供应受限的碱性淡水水体中,来自蓝细菌的n -C17 烷的δ13 C值会显著偏正[130 ] . ...
An isotopic biogeochemical assessment of shifts in organic matter input to Holocene sediments from Mud Lake, Florida
2
2001
... 一项在非洲湖泊开展的研究,探讨了末次间冰期湖泊沉积物中来自葡萄球藻的葡萄球藻烯的δ13 C变化[128 ] .在湖泊沉积物中,短链异构烷烃(主要是C17 和C18 )主要来自蓝细菌[129 ] .来自蓝细菌的这些短链烷烃的δ13 C值通常表现出较大的变化范围[120 ,130 ,131 ] ,例如,在CO2 供应受限的碱性淡水水体中,来自蓝细菌的n -C17 烷的δ13 C值会显著偏正[130 ] . ...
... [130 ]. ...
Source characterization of sedimentary organic matter using molecular and stable isotopic composition of n -alkanes and fatty acids in sediment core from Lake Dianchi, China
1
2014
... 一项在非洲湖泊开展的研究,探讨了末次间冰期湖泊沉积物中来自葡萄球藻的葡萄球藻烯的δ13 C变化[128 ] .在湖泊沉积物中,短链异构烷烃(主要是C17 和C18 )主要来自蓝细菌[129 ] .来自蓝细菌的这些短链烷烃的δ13 C值通常表现出较大的变化范围[120 ,130 ,131 ] ,例如,在CO2 供应受限的碱性淡水水体中,来自蓝细菌的n -C17 烷的δ13 C值会显著偏正[130 ] . ...
Encyclopedia of Ocean Sciences(Third Edition)
1
2018
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
A review on techniques and applications of biomarker compound-specific radiocarbon analysis
1
2017
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
生物标志物单体放射性碳同位素分析技术的发展
1
2017
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
Pre-aged soil organic carbon as a major component of the Yellow River suspended load:Regional significance and global relevance
1
2015
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
Methods and future directions for paleoclimatology in the Maya Lowlands
1
2016
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
Hydrologic control of carbon cycling and aged carbon discharge in the Congo River basin
1
2016
... (1)重视实验技术.目前主流的单体碳测试仪器要求化合物的量约数十微克,这接近常规色谱柱的容量上限,容易造成色谱峰不规则,影响峰分离,并降低测试精度.另一方面,较高的浓度要求使得很多样品量较小的样品无法进行准确地单体碳分析.因此,需要建立小样品量的单体碳测试方法.对于化合物组成复杂的样品,需要建立目标化合物分离纯化的方法,除了常规的分子筛、尿素络合方法,可以考虑制备气相色谱和制备液相色谱法.对于分子量相对较大的脂类(如GDGTs,BHPs),需要开发并推广能够直接测试其碳同位素组成的新技术[24 ,25 ] .单体放射性碳同位素也在得到日益广泛地重视[132 ,133 ] ,在年代学、陆地有机质搬运等领域取得了重要的进展[134 ,135 ,136 ] . ...
Dual isotope evidence for sedimentary integration of plant wax biomarkers across an Andes-Amazon elevation transect
1
2018
... (3)同一脂类碳同位素和氢同位素的联合也是值得考虑的方向.叶蜡脂类的单体碳和氢同位素组成会受到类似环境条件的控制,因此联合2种同位素比值可以更好地识别环境控制条件.Feakins等[137 ] 在南美洲亚马逊流域开展的一项研究中,利用正构烷烃和正构脂肪酸碳同位素和氢同位素的联合,能够更好地识别河流颗粒物中有机质的来源. ...




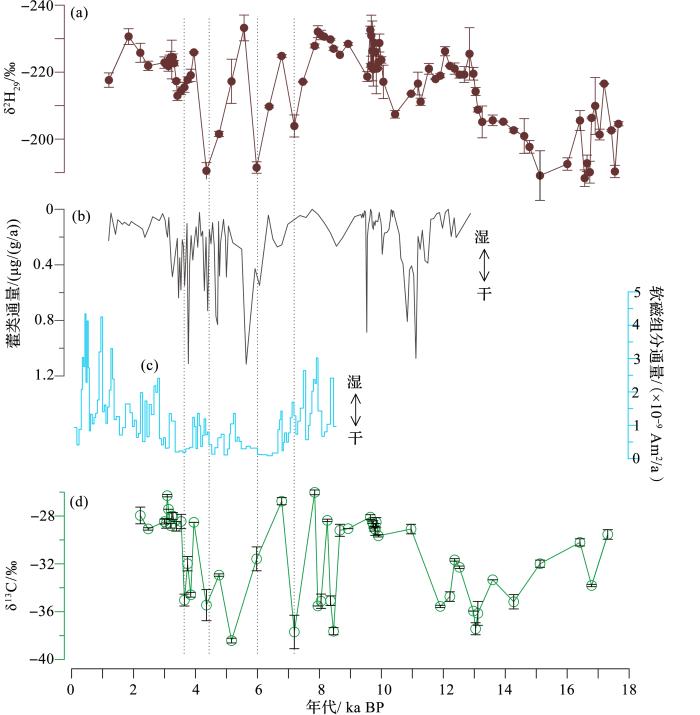
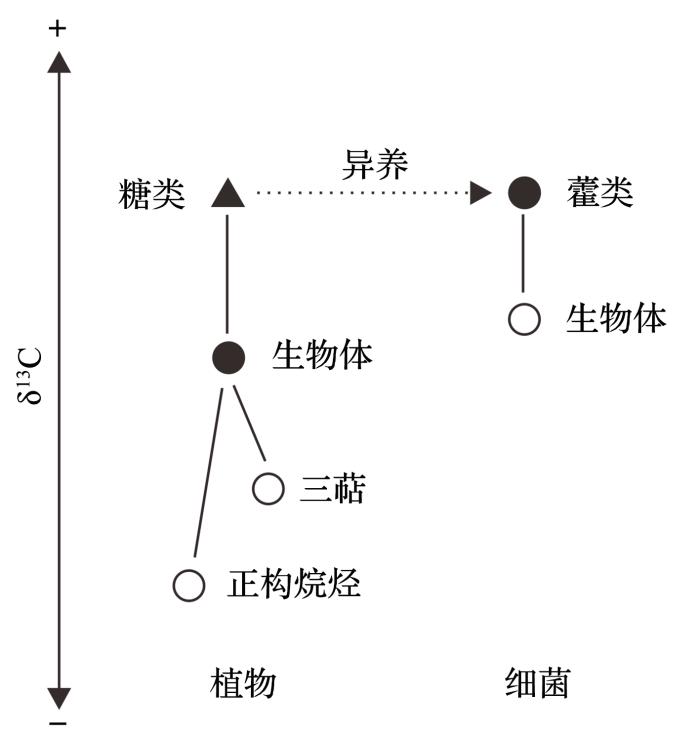
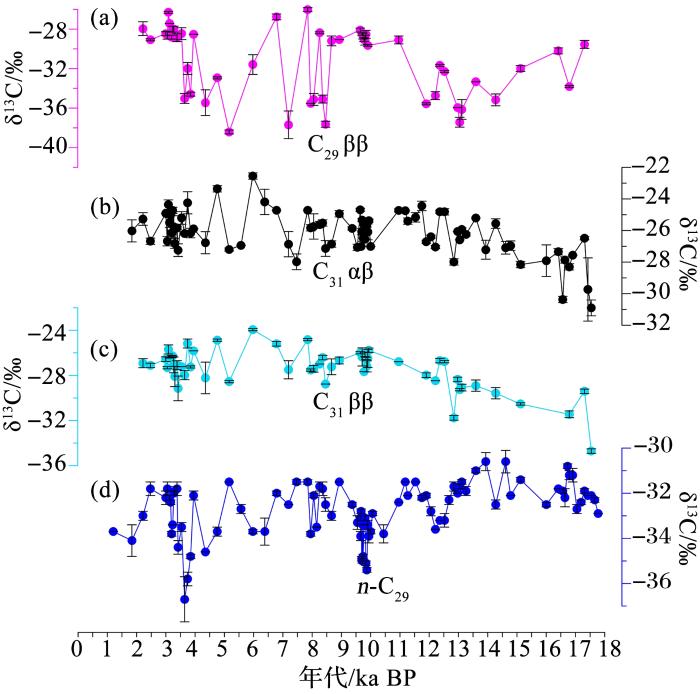

 甘公网安备62010202000687
甘公网安备62010202000687{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}