. Nitrogen cycles: Past, present, and future
2
2004
... 氮素是一种重要的生源要素,对人类生命、生活等具有重要意义.在人类能够合成氨之前,全球活性氮主要通过生物固氮产生.据估算,1860年全球活性氮的年产生总量为262 Tg;人类能够合成氨后,人为活性氮数量逐年增加,在20世纪90年代初期,Haber-Bosch过程每年合成的活性氮总量达到100 Tg,预期到2050年将达到165 Tg[1].人为活性氮的大量投入为提高粮食产量、消除饥饿做出了不可替代的贡献,人类还将继续依赖大量的氮肥投入以满足对粮食的需求.另一方面,人为活性氮的大量投入已经造成了严重的区域环境问题,如水体富营养化和生物多样性破坏等.现在我们面临协调粮食安全、生态环境安全和气候变化的三重挑战[2]. ...
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
Managing nitrogen for sustainable development
1
2015
... 氮素是一种重要的生源要素,对人类生命、生活等具有重要意义.在人类能够合成氨之前,全球活性氮主要通过生物固氮产生.据估算,1860年全球活性氮的年产生总量为262 Tg;人类能够合成氨后,人为活性氮数量逐年增加,在20世纪90年代初期,Haber-Bosch过程每年合成的活性氮总量达到100 Tg,预期到2050年将达到165 Tg[1].人为活性氮的大量投入为提高粮食产量、消除饥饿做出了不可替代的贡献,人类还将继续依赖大量的氮肥投入以满足对粮食的需求.另一方面,人为活性氮的大量投入已经造成了严重的区域环境问题,如水体富营养化和生物多样性破坏等.现在我们面临协调粮食安全、生态环境安全和气候变化的三重挑战[2]. ...
Centennial-scale analysis of the creation and fate of reactive nitrogen in China (1910-2010)
1
2013
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
Nitrogen Soils of China
1
1992
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
中国土壤氮素
1
1992
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
Measurement of anaerobic ammonium oxidation activity in soil systems and identification of Planctomycetes
1
2003
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
Anaerobic ammonium oxidation by anammox bacteria in the Black Sea
1
2003
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
Anaerobic ammonium oxidation measured in sediments along the Thames estuary, United Kingdom
1
2003
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
Nitrogen cycling in forest soils across climate gradients in Eastern China
2
2011
... 绝大部分活性氮(陆地生物固定的和Haber-Bosch过程生产的)首先进入土壤[1,3],除极少部分残留在土壤中外,绝大部分在土壤中发生一系列的转化,以不同的形态进入到生物、大气和水体.进入生物、大气和水体的活性氮的形态和相对比例受诸多因素的影响,如气候、植被、地形、氮肥管理措施等外因和土壤氮转化特性等内因.理论上,土壤氮的各个转化过程及其速率的组合对土壤氮去向具有“调配器”的作用.总体而言,关于土壤氮转化过程,当前的研究主要集中于参与各过程的功能微生物、转化过程发生的环境条件、产物以及转化速率的定量化.现在对于土壤中单个氮转化过程(如矿化、硝化、反硝化、微生物同化、厌氧氨氧化、NO异化还原为铵等)的微生物机理和影响因素已经有了较深的认识[4,5,6,7].但是,土壤氮形态众多,极为复杂,且不同氮形态间的转化过程同时发生、相互影响、相互作用,共同影响氮形态及去向,需要同时定量各个过程的速率才能更好地阐明氮动态机理[8];另一方面,可以直接测定的土壤氮库指标却很少,因此很难同时定量多个过程的转化速率.已有的大部分研究都是假定其他过程速率不变的前提下,一次定量单个或少数几个过程的速率,限制了对氮过程间相互作用的认识.需要采用新的研究方法实现同时定量土壤中主要氮转化过程的速率,基于此深入研究土壤氮转化过程特点及其驱动的土壤氮“调配器”作用,研究结果可以为因土因作物合理施用氮肥、制订氮污染控制对策措施等提供理论依据. ...
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
On progress in gross nitrogen transformation using 15N isotopic pool dilution
4
2009
... 依据存在形态,土壤氮库可分为有机氮库和无机氮库,有机氮是土壤中最大的氮库,其组成极为复杂,可利用性很低;无机氮组成相对简单,主要为铵态氮(NH)和硝态氮(NO),其仅占土壤全氮的极小部分,但却是土壤中主要的可利用氮形态.土壤中各种形态的氮可以相互转化,而且这些转化过程每时每刻都在发生.每种形态氮的含量往往受多个转化过程控制.例如,即使在没有外源氮输入、植物吸收和淋溶、径流损失的实验室培养条件下,硝化作用产生NO,NO会同时通过微生物同化、反硝化作用而被消耗,NO的含量则为NO产生与消耗的各个过程速率的综合结果.在自然生态系统中,影响土壤有机氮、NH和NO含量变化的过程更多[9],定量各个氮过程转化速率是认识土壤氮动态的基础. ...
... 依据测定方法,土壤氮的转化速率可区分为初级转化速率(Gross rate of nitrogen transformation)和净转化速率(Net rate of nitrogen transformation).初级转化速率是指单位时间内一种形态氮转化为另一种形态氮的实际速率,如NH硝化成为NO的速率,称为初级硝化速率,有机氮矿化成NH的速率称为初级氮矿化速率;净转化速率则是指单位时间内某一种形态氮含量的变化速率,如单位时间内NO含量变化为净硝化速率,单位时间内无机氮(NH与NO总和)含量变化为净矿化速率[9,10].净转化速率是某一种形态氮含量的净变化量,但并不能定量引起该种形态氮含量变化的各个氮转化过程的实际速率.相比之下,测定土壤氮初级转化速率,不仅可以确定土壤中同时发生的各个氮转化过程的实际速率,而且可以计算出各种形态氮的含量变化量(即净转化速率)[9]. ...
... [9]. ...
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
程谊,蔡祖聪,张金波,15N同位素稀释法测定土壤氮素总转化速率研究进展
4
2009
... 依据存在形态,土壤氮库可分为有机氮库和无机氮库,有机氮是土壤中最大的氮库,其组成极为复杂,可利用性很低;无机氮组成相对简单,主要为铵态氮(NH)和硝态氮(NO),其仅占土壤全氮的极小部分,但却是土壤中主要的可利用氮形态.土壤中各种形态的氮可以相互转化,而且这些转化过程每时每刻都在发生.每种形态氮的含量往往受多个转化过程控制.例如,即使在没有外源氮输入、植物吸收和淋溶、径流损失的实验室培养条件下,硝化作用产生NO,NO会同时通过微生物同化、反硝化作用而被消耗,NO的含量则为NO产生与消耗的各个过程速率的综合结果.在自然生态系统中,影响土壤有机氮、NH和NO含量变化的过程更多[9],定量各个氮过程转化速率是认识土壤氮动态的基础. ...
... 依据测定方法,土壤氮的转化速率可区分为初级转化速率(Gross rate of nitrogen transformation)和净转化速率(Net rate of nitrogen transformation).初级转化速率是指单位时间内一种形态氮转化为另一种形态氮的实际速率,如NH硝化成为NO的速率,称为初级硝化速率,有机氮矿化成NH的速率称为初级氮矿化速率;净转化速率则是指单位时间内某一种形态氮含量的变化速率,如单位时间内NO含量变化为净硝化速率,单位时间内无机氮(NH与NO总和)含量变化为净矿化速率[9,10].净转化速率是某一种形态氮含量的净变化量,但并不能定量引起该种形态氮含量变化的各个氮转化过程的实际速率.相比之下,测定土壤氮初级转化速率,不仅可以确定土壤中同时发生的各个氮转化过程的实际速率,而且可以计算出各种形态氮的含量变化量(即净转化速率)[9]. ...
... [9]. ...
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Isotopic dilution methods to determine the gross transformation rates of nitrogen, phosphorus, and sulfur in soil: A review of the theory, methodologies, and limitations
2
2000
... 依据测定方法,土壤氮的转化速率可区分为初级转化速率(Gross rate of nitrogen transformation)和净转化速率(Net rate of nitrogen transformation).初级转化速率是指单位时间内一种形态氮转化为另一种形态氮的实际速率,如NH硝化成为NO的速率,称为初级硝化速率,有机氮矿化成NH的速率称为初级氮矿化速率;净转化速率则是指单位时间内某一种形态氮含量的变化速率,如单位时间内NO含量变化为净硝化速率,单位时间内无机氮(NH与NO总和)含量变化为净矿化速率[9,10].净转化速率是某一种形态氮含量的净变化量,但并不能定量引起该种形态氮含量变化的各个氮转化过程的实际速率.相比之下,测定土壤氮初级转化速率,不仅可以确定土壤中同时发生的各个氮转化过程的实际速率,而且可以计算出各种形态氮的含量变化量(即净转化速率)[9]. ...
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Equations for following nutrient transformations in soil utilizing tracer data
1
1954
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Estimation of parameters in complex 15N tracing models by Monte Carlo sampling
4
2007
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
... [12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
... [12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
... [12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Simultaneous estimation of several nitrogen cycle rates using 15N: Theory and application
1
1986
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Calculating gross N transformation rates of 15N pool dilution experiments with acid forest litter: Analytical and numerical approaches
1
1992
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Estimating nitrogen transformations rates in surface aerobic soil of a paddy field
1
1994
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
A 15N tracing model to analyse N transformations in old grassland soil
1
2004
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
A model for calculating nitrogen fluxes in soil using 15N tracing
3
1998
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
... [17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
... [17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
15N tracing models with a Monte Carlo optimization procedure provide new insights on gross N transformations in soils
1
2007
... 15N稳定同位素稀释方法是测定土壤氮初级转化速率的唯一方法[10].20世纪50年代,Kirkham 等[11]提出的同位素稀释法基本原理为利用15N示踪技术测定氮素初级转化速率奠定了基础,由此提出了定量氮初级转化速率的算术分析方法,为研究土壤氮初级转化速率提供了有效的方法[9].但是该方法存在一些缺点:不能评价参数的不确定性;仅计算被标记氮库的产生速率和消耗速率,不能准确量化具体的转化过程;只能定量少数几个氮过程的速率,而土壤中各种形态氮之间的转化过程是同时发生的,且能够测定的氮库数经常少于需要计算的过程数,因此,仅采用算术分析方法难以计算各过程的初级转化速率.随着计算机技术的快速发展,数值分析方法在氮转化过程研究中的应用越来越受到重视[12].学者们先后建立了多种数值分析模型,基于15N成对标记示踪技术和数值分析模型相结合,实现同时定量土壤中多个氮转化过程的初级转化速率[12,13,14,15,16,17].现在应用较为广泛的是FLUAZ 模型[17]和MCMC模型[12].应用 FLUAZ 模型通常可以计算出土壤中6个主要氮过程的初级转化速率,即有机氮矿化速率、NH生物同化速率、NO生物同化速率、硝化作用速率、氨挥发速率或反硝化作用速率和标记后生物同化氮的再矿化速率[17].为了提高计算结果的准确性,Müller 等[12]把马尔柯夫链蒙特卡洛随机采样方法(Markov Chain Monte Carlo, MCMC)引入到数值分析模型中,从而有效地避免了模型运算过程中的局域寻优问题,确保找到真正的全局最优,结果更能反映土壤氮素转化的真实情形[18].这些研究方法已经成为深入研究土壤氮转化过程的有力工具. ...
Spatial and seasonal variations of gross nitrogen transformations and microbial biomass in a Northeastern US grassland
1
2002
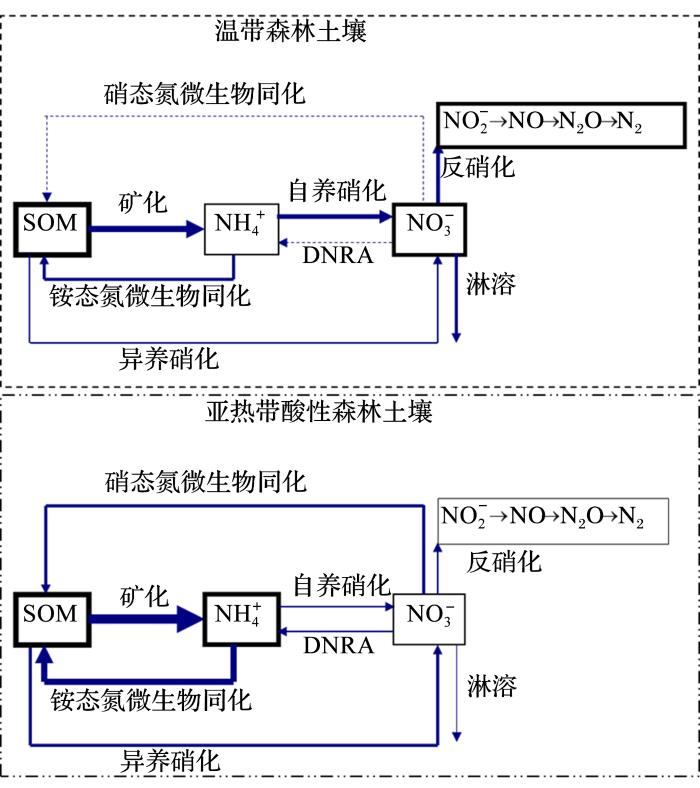
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Gross nitrogen transformations in adjacent native and plantation forests of subtropical Australia
1
2007
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Mechanisms of soil N dynamics following long-term application of organic fertilizers to subtropical rain-fed purple soil in China
2
2015
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
... 基于河南封丘农田生态系统国家野外研究站和四川盐亭农田生态系统国家野外研究站的长期肥料试验的研究结果表明,不同的肥料管理措施对土壤氮转化过程的初级转化速率有明显的影响,虽然不同地区的响应也存在明显的差异,但是都有一个一致的结论,在等氮量施肥的条件下,土壤氮淋溶损失量或土壤N2O排放均随土壤初级硝化速率的降低呈指数降低,而作物产量随土壤有机氮初级矿化速率增长呈线性增加[21,27],说明土壤氮转化特点与氮去向有密切的关系,土壤氮的淋溶损失、活性氮向大气的扩散和作物产量均受到氮转化过程的控制,不同农业管理方式对氮利用率、氮损失、作物产量等的影响主要是通过调控土壤氮转化过程特点实现的.因此,深入认识自然和人为因素对土壤氮转化特性的影响及其作用机理,理论上将有助于根据土壤与作物特性,采取针对性的措施,通过发挥土壤氮形态“调配器”的作用,提高氮肥利用率,减少土壤活性氮向环境的扩散. ...
Soil nitrogen conservation mechanisms in a pristine south Chilean Nothofagus forest ecosystem
1
2007
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Microbial immobilization of ammonium and nitrate in relation to ammonification and nitrification rates in organic and conventional cropping systems
1
2003
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Estimation of gross nitrogen transformations and nitrogen retention in grassland soils using FLUAZ
1
2005
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Mechanisms for the retention of inorganic N in acidic forest soils of southern China
1
2013
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Agricultural land use affects nitrate production and conservation in humid subtropical soils in China
1
2013
... 近年来,土壤氮初级转化速率的研究报道不断增加,广泛地应用于不同土壤类型、植被类型、地形部位和农业管理措施等条件下土壤氮素转化特点的研究[19,20,21].通过这些研究,极大地提高了我们对土壤氮循环规律及其影响因素的认识.至今,NO异化还原为铵(Dissimilatory Nitrate Reduction to Ammonium,DNRA)过程尚不能用直接测定方法确定,而数值优化模型提供了一种定量该过程速率的方法.Huygens等[22]使用该方法研究发现,在智利年降雨量达到7 000 mm以上的森林土壤中,DNRA过程消耗的NO占其总消耗量的99%,从而使高降雨量的森林生态系统有效地保持了氮素.此外,运用此方法,还可以有效地区分土壤微生物对NH或NO同化的偏好[23],测定土壤有机氮初级矿化速率、硝化作用速率及无机氮总同化速率可以认识不同土壤保持无机氮的能力大小[24],丰富对土壤动态的认识.作者所在研究团队从2008年开始建立并完善了使用数值分析模型同时定量土壤中10个主要氮过程初级转化速率的方法,系统研究了中国亚热带酸性土壤氮关键过程的初级转化速率.大量的研究结果表明,初级转化速率方法能够深入剖析土壤氮转化特征,阐释土壤无机氮形态和氮保持机理以及农业利用对土壤氮动态的影响.例如,在热带—亚热带高温多雨的气候条件下,土壤发生强烈的脱硅富铁铝过程,为什么易于迁移的氮元素却能相对富集?传统的基于土壤氮净转化速率研究建立的知识体系不能解释这一现象.作者采用氮初级转化速率方法对比研究了温带和亚热带森林土壤氮素关键转化过程的初级转化速率[25],明确了亚热带酸性森林土壤具有以下显著区别于温带土壤的氮转化特点(图1):土壤有机氮周转迅速,无机氮产生量大,可以为生态系统提供充足的无机氮;自养硝化能力弱,使得无机氮以NH形态为主,减少了氮的淋溶风险,而酸性的土壤环境有效避免了NH的挥发损失;具有很强的NO固持能力,通过微生物同化及DNRA途径有效地转化硝化过程产生的NO,从而进一步降低NO淋溶风险;反硝化作用弱,避免了NO的反硝化损失.这些独特的氮转化特点,使得亚热带湿润地区酸性森林土壤无机氮以NH为主,与湿润多雨的气候条件相适应,具有较高的无机氮保持能力,能够很好地解释该生态系统中氮素相对富集的现象.通过对比研究亚热带森林和农田土壤氮转化特点,发现不同于森林土壤,农业土壤的无机氮以NO为主,主要原因是农业利用显著激发了自养硝化过程速率,NO微生物同化能力却显著降低,使得农业土壤NO产生量高,消耗量低,导致无机氮以NO为主,破坏了该区域自然土壤所具有的保氮能力[26].因此,土壤各个氮转化过程相互作用,即氮转化过程特点,会决定无机氮的主导形态,其与环境条件、气候特点(主要是降水)的契合程度可能是决定土壤氮动态的重要因素.可见,初级转化速率的研究方法已经将传统的只关注氮库含量变化的净转化研究真正推进到了关注控制氮库含量变化的各个氮转化过程的研究,能够深入认识土壤氮动态机制,有望为因地制宜地氮素调控提供理论支持. ...
Effects of long-term repeated mineral and organic fertilizer applications on soil nitrogen transformations
1
2012
... 基于河南封丘农田生态系统国家野外研究站和四川盐亭农田生态系统国家野外研究站的长期肥料试验的研究结果表明,不同的肥料管理措施对土壤氮转化过程的初级转化速率有明显的影响,虽然不同地区的响应也存在明显的差异,但是都有一个一致的结论,在等氮量施肥的条件下,土壤氮淋溶损失量或土壤N2O排放均随土壤初级硝化速率的降低呈指数降低,而作物产量随土壤有机氮初级矿化速率增长呈线性增加[21,27],说明土壤氮转化特点与氮去向有密切的关系,土壤氮的淋溶损失、活性氮向大气的扩散和作物产量均受到氮转化过程的控制,不同农业管理方式对氮利用率、氮损失、作物产量等的影响主要是通过调控土壤氮转化过程特点实现的.因此,深入认识自然和人为因素对土壤氮转化特性的影响及其作用机理,理论上将有助于根据土壤与作物特性,采取针对性的措施,通过发挥土壤氮形态“调配器”的作用,提高氮肥利用率,减少土壤活性氮向环境的扩散. ...
Accumulation of soil nitrate nitrogen in the process of ecological and its effcts in plant nitrogen nutrition in semiarid areas
1
2004
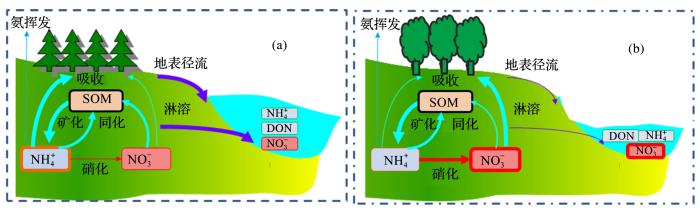
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
半干旱区生态过程变化中土壤硝态氮累积及其在植物氮素营养中的作用
1
2004
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
Significant accumulation of nitrate in Chinese semi-humid croplands
1
2016
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
Terrestrial N cycling associated with climate and plant specific N preferences: A review
2
2018
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
... [30,32]. ...
Water balance creates a threshold in soil pH at the global scale
2
2016
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
... [31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
The characteristics of soil N transformations regulate the composition of hydrologic N export from terrestrial ecosystem
5
2016
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
... [32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
... [32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
... [32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
... ,32]. ...
Process characteristics of nitrogen loss by runoff in different land use watersheds in purple soil hilly region
1
2011
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
紫色土丘陵区不同土地利用类型小流域氮素流失规律初探
1
2011
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
Measurements of nitrate leaching from a hillslope cropland in the central Sichuan Basin, China
1
2009
... 土壤氮转化过程如何调控氮去向呢?作者所在研究团队系统研究了土壤氮转化特性在氮生物地球化学循环中的作用.NH和NO是土壤中主要的可利用氮,将两者有效地保持在土壤中是提高有效性氮数量和植物氮利用率的基础.因为土壤胶体普遍带负电荷,所以一般认为NH容易被土壤吸附固定,而NO易随水迁移.但是,事实上土壤保持无机氮的能力与土壤氮转化特点决定的无机氮主导形态、土壤环境条件以及所处的气候条件(主要是降水量)有密切的关系.NH虽然容易被土壤吸附固定,但是在碱性土壤中却很容易发生氨挥发损失,酸性土壤环境更有利于NH保持;而在干旱条件下因缺少水分移动,NO会相对稳定,积累在土壤剖面中,如在我国华北、西北等地经常观测到过量施用氮肥导致大量的NO积累在土壤剖面中,需要相当长的时间才能进入地下水体[28,29].土壤氮转化过程,特别是硝化作用的强弱,是决定土壤无机氮主导型态是以NH为主还是以NO为主的内因,土壤pH是控制该过程的关键因素,在一定范围内,硝化过程速率随土壤pH的增加而增加[8,30].大量的研究数据表明,土壤pH与气候条件(主要是干燥度)有密切的关系[31].一般来讲,干旱、半干旱地区发育的地带性土壤呈中性和碱性,如我国的西北地区等,硝化作用强,土壤无机氮以NO为主;而湿润地区发育的地带性土壤多呈酸性,如我国的南方地区等,硝化作用弱,无机氮以NH为主[31,32].由于地带性土壤氮转化过程特点与气候条件契合程度高,会表现出较强的保持无机氮的能力(图2).Zhang 等[32]的研究结果发现,不同的自然生态系统径流中氮形态和浓度存在明显的差异,有的以NO为主,有的则以溶解有机氮(Dissolved Organic Nitrogen,DON)占主导,而NH(平均0.1 mg N/L)和DON(平均0.4 mg N/L)浓度差别不大,氮形态组成的变化主要是NO浓度引起的.典型湿润亚热带森林系统径流中NO平均浓度只有0.1 mg N/L,因此推测,虽然多雨气候条件下径流量很大,径流氮损失总量可能并不高;半湿润、半干旱—干旱森林/草地系统径流中NO平均浓度大于3 mg N/L,成为主导的氮形态,但由于干燥少雨,理论上径流氮损失总量同样不会太高[32].然而,如果两者契合程度不高,例如非地带性土壤——碱性紫色土区域,地处湿润地区,但是土壤呈碱性,硝化能力很强,使得径流中NO浓度大(平均大于2 mg N/L),雨季大量的径流导致氮损失严重.因此,NO成为紫色土小流域氮素流失的主要形式[32,33,34].溪流中氮形态组成与土壤及土壤溶液中氮形态组成高度一致,说明两者之间有着密切联系,证明土壤氮的转化特点决定无机氮主导形态,其与气候条件的契合程度会控制生态系统径流氮损失[30,32]. ...
The calcicole-calcifuge problem revisited
1
1999
... 提高作物氮利用率是实现减氮增效目标的关键途径,也是土壤氮循环研究的重要目标.氮素利用率取决于2个因素,一是作物氮吸收效率,一是作物对所吸收氮素的利用效率.高作物氮吸收效率是高氮利用率的前提条件.虽然通常认为植物既可以吸收利用NH,也可以吸收利用NO,但大量的研究结果已经证明,一些植物会偏好吸收NH,而另一些植物则会喜好吸收NO,也有一部分植物没有明显的偏好[35,36].植物对于无机氮形态的偏好主要取决于其起源的土壤条件.一般地,起源于无机氮以NH为主条件下的植物喜好吸收NH,相反,起源于以NO为主条件下的植物会偏好NO[37,38](图2).所以,一般情况下水生植物和强酸性土壤上起源的植物喜好NH,如水稻、茶树、松树等.理论上可以预期,如果供应的无机氮形态与植物喜好吸收的氮形态一致,植物将会更好地吸收供应的氮;相反,如果供应的无机氮形态与植物喜好的氮形态不一致,有可能不利于作物对氮的吸收,对生长发育产生不利影响[36].Zhang等[39]研究结果支持这一推测,发现喜硝作物黄瓜对15N标记NO的吸收率(40.1%~47.7%)高于对NH的吸收率(12.6%~39.3%),而喜铵作物马铃薯恰好相反,说明施用的氮肥形态和植物氮形态喜好的契合程度确实可以影响植物氮吸收.作者所在团队在福建红壤和四川石灰性紫色土区域的田间试验也获得了一致的结果,数据显示15N标记的尿素作基肥施用,种植在硝化能力弱的土壤上的水稻氮利用率、产量均高于硝化能力强的土壤,而小麦(喜硝)则相反. ...
Responses of crop plants to ammonium and nitrate N
3
2013
... 提高作物氮利用率是实现减氮增效目标的关键途径,也是土壤氮循环研究的重要目标.氮素利用率取决于2个因素,一是作物氮吸收效率,一是作物对所吸收氮素的利用效率.高作物氮吸收效率是高氮利用率的前提条件.虽然通常认为植物既可以吸收利用NH,也可以吸收利用NO,但大量的研究结果已经证明,一些植物会偏好吸收NH,而另一些植物则会喜好吸收NO,也有一部分植物没有明显的偏好[35,36].植物对于无机氮形态的偏好主要取决于其起源的土壤条件.一般地,起源于无机氮以NH为主条件下的植物喜好吸收NH,相反,起源于以NO为主条件下的植物会偏好NO[37,38](图2).所以,一般情况下水生植物和强酸性土壤上起源的植物喜好NH,如水稻、茶树、松树等.理论上可以预期,如果供应的无机氮形态与植物喜好吸收的氮形态一致,植物将会更好地吸收供应的氮;相反,如果供应的无机氮形态与植物喜好的氮形态不一致,有可能不利于作物对氮的吸收,对生长发育产生不利影响[36].Zhang等[39]研究结果支持这一推测,发现喜硝作物黄瓜对15N标记NO的吸收率(40.1%~47.7%)高于对NH的吸收率(12.6%~39.3%),而喜铵作物马铃薯恰好相反,说明施用的氮肥形态和植物氮形态喜好的契合程度确实可以影响植物氮吸收.作者所在团队在福建红壤和四川石灰性紫色土区域的田间试验也获得了一致的结果,数据显示15N标记的尿素作基肥施用,种植在硝化能力弱的土壤上的水稻氮利用率、产量均高于硝化能力强的土壤,而小麦(喜硝)则相反. ...
... [36].Zhang等[39]研究结果支持这一推测,发现喜硝作物黄瓜对15N标记NO的吸收率(40.1%~47.7%)高于对NH的吸收率(12.6%~39.3%),而喜铵作物马铃薯恰好相反,说明施用的氮肥形态和植物氮形态喜好的契合程度确实可以影响植物氮吸收.作者所在团队在福建红壤和四川石灰性紫色土区域的田间试验也获得了一致的结果,数据显示15N标记的尿素作基肥施用,种植在硝化能力弱的土壤上的水稻氮利用率、产量均高于硝化能力强的土壤,而小麦(喜硝)则相反. ...
... 但是,也有一些研究与以上结论不一致,认为氮肥形态和植物氮形态喜好的契合程度对氮肥的吸收以及作物产量没有影响[36,40].我们认为一个重要的原因是这些研究没有考虑土壤氮转化过程对于施入氮肥形态的影响.通过本文第4部分的阐述,我们已经明确了无论是土壤有机氮矿化产生的氮,还是施入到土壤的氮肥,都会在土壤中发生转化,残留在土壤中的无机氮最终将以土壤氮转化特性决定的形态存在,因此氮转化过程特点会明显影响氮肥形态和植物氮形态喜好的契合程度.Zhang等[39]发现,在15N标记的NH施入土壤5天后,硝化作用弱的土壤中有56%~75%仍然以NH形态存在,NO仅占2%;而硝化作用强的土壤正相反,只有不到2%的15N 以NH形态存在,NO比例高达31%~45%.对于硝化能力强的土壤,由于施入的NH能够快速转化为NO,喜铵作物对铵态氮肥和硝态氮肥的吸收率差异很小(19.5% vs. 18.6%),而在硝化能力弱的土壤种植的喜铵作物对铵态氮肥的吸收率则显著高于硝态氮肥(31.0% vs. 22.3%)[39].同样,喜硝作物种植在硝化能力强的土壤,其对铵态氮肥的吸收率要明显高于硝化能力弱的土壤[39].可见,土壤氮转化特征决定的土壤无机氮主导形态与作物氮形态喜好的契合程度对作物氮肥利用率具有重要影响. ...
Do plants need nitrate?The mechanisms by which nitrogen form affects plants
1
2013
... 提高作物氮利用率是实现减氮增效目标的关键途径,也是土壤氮循环研究的重要目标.氮素利用率取决于2个因素,一是作物氮吸收效率,一是作物对所吸收氮素的利用效率.高作物氮吸收效率是高氮利用率的前提条件.虽然通常认为植物既可以吸收利用NH,也可以吸收利用NO,但大量的研究结果已经证明,一些植物会偏好吸收NH,而另一些植物则会喜好吸收NO,也有一部分植物没有明显的偏好[35,36].植物对于无机氮形态的偏好主要取决于其起源的土壤条件.一般地,起源于无机氮以NH为主条件下的植物喜好吸收NH,相反,起源于以NO为主条件下的植物会偏好NO[37,38](图2).所以,一般情况下水生植物和强酸性土壤上起源的植物喜好NH,如水稻、茶树、松树等.理论上可以预期,如果供应的无机氮形态与植物喜好吸收的氮形态一致,植物将会更好地吸收供应的氮;相反,如果供应的无机氮形态与植物喜好的氮形态不一致,有可能不利于作物对氮的吸收,对生长发育产生不利影响[36].Zhang等[39]研究结果支持这一推测,发现喜硝作物黄瓜对15N标记NO的吸收率(40.1%~47.7%)高于对NH的吸收率(12.6%~39.3%),而喜铵作物马铃薯恰好相反,说明施用的氮肥形态和植物氮形态喜好的契合程度确实可以影响植物氮吸收.作者所在团队在福建红壤和四川石灰性紫色土区域的田间试验也获得了一致的结果,数据显示15N标记的尿素作基肥施用,种植在硝化能力弱的土壤上的水稻氮利用率、产量均高于硝化能力强的土壤,而小麦(喜硝)则相反. ...
The response of grassland species to nitrate versus ammonium coincides with their pH optima
1
2014
... 提高作物氮利用率是实现减氮增效目标的关键途径,也是土壤氮循环研究的重要目标.氮素利用率取决于2个因素,一是作物氮吸收效率,一是作物对所吸收氮素的利用效率.高作物氮吸收效率是高氮利用率的前提条件.虽然通常认为植物既可以吸收利用NH,也可以吸收利用NO,但大量的研究结果已经证明,一些植物会偏好吸收NH,而另一些植物则会喜好吸收NO,也有一部分植物没有明显的偏好[35,36].植物对于无机氮形态的偏好主要取决于其起源的土壤条件.一般地,起源于无机氮以NH为主条件下的植物喜好吸收NH,相反,起源于以NO为主条件下的植物会偏好NO[37,38](图2).所以,一般情况下水生植物和强酸性土壤上起源的植物喜好NH,如水稻、茶树、松树等.理论上可以预期,如果供应的无机氮形态与植物喜好吸收的氮形态一致,植物将会更好地吸收供应的氮;相反,如果供应的无机氮形态与植物喜好的氮形态不一致,有可能不利于作物对氮的吸收,对生长发育产生不利影响[36].Zhang等[39]研究结果支持这一推测,发现喜硝作物黄瓜对15N标记NO的吸收率(40.1%~47.7%)高于对NH的吸收率(12.6%~39.3%),而喜铵作物马铃薯恰好相反,说明施用的氮肥形态和植物氮形态喜好的契合程度确实可以影响植物氮吸收.作者所在团队在福建红壤和四川石灰性紫色土区域的田间试验也获得了一致的结果,数据显示15N标记的尿素作基肥施用,种植在硝化能力弱的土壤上的水稻氮利用率、产量均高于硝化能力强的土壤,而小麦(喜硝)则相反. ...
Ecological and practical significances of crop species preferential N uptake matching with soil N dynamics
5
2016
... 提高作物氮利用率是实现减氮增效目标的关键途径,也是土壤氮循环研究的重要目标.氮素利用率取决于2个因素,一是作物氮吸收效率,一是作物对所吸收氮素的利用效率.高作物氮吸收效率是高氮利用率的前提条件.虽然通常认为植物既可以吸收利用NH,也可以吸收利用NO,但大量的研究结果已经证明,一些植物会偏好吸收NH,而另一些植物则会喜好吸收NO,也有一部分植物没有明显的偏好[35,36].植物对于无机氮形态的偏好主要取决于其起源的土壤条件.一般地,起源于无机氮以NH为主条件下的植物喜好吸收NH,相反,起源于以NO为主条件下的植物会偏好NO[37,38](图2).所以,一般情况下水生植物和强酸性土壤上起源的植物喜好NH,如水稻、茶树、松树等.理论上可以预期,如果供应的无机氮形态与植物喜好吸收的氮形态一致,植物将会更好地吸收供应的氮;相反,如果供应的无机氮形态与植物喜好的氮形态不一致,有可能不利于作物对氮的吸收,对生长发育产生不利影响[36].Zhang等[39]研究结果支持这一推测,发现喜硝作物黄瓜对15N标记NO的吸收率(40.1%~47.7%)高于对NH的吸收率(12.6%~39.3%),而喜铵作物马铃薯恰好相反,说明施用的氮肥形态和植物氮形态喜好的契合程度确实可以影响植物氮吸收.作者所在团队在福建红壤和四川石灰性紫色土区域的田间试验也获得了一致的结果,数据显示15N标记的尿素作基肥施用,种植在硝化能力弱的土壤上的水稻氮利用率、产量均高于硝化能力强的土壤,而小麦(喜硝)则相反. ...
... 但是,也有一些研究与以上结论不一致,认为氮肥形态和植物氮形态喜好的契合程度对氮肥的吸收以及作物产量没有影响[36,40].我们认为一个重要的原因是这些研究没有考虑土壤氮转化过程对于施入氮肥形态的影响.通过本文第4部分的阐述,我们已经明确了无论是土壤有机氮矿化产生的氮,还是施入到土壤的氮肥,都会在土壤中发生转化,残留在土壤中的无机氮最终将以土壤氮转化特性决定的形态存在,因此氮转化过程特点会明显影响氮肥形态和植物氮形态喜好的契合程度.Zhang等[39]发现,在15N标记的NH施入土壤5天后,硝化作用弱的土壤中有56%~75%仍然以NH形态存在,NO仅占2%;而硝化作用强的土壤正相反,只有不到2%的15N 以NH形态存在,NO比例高达31%~45%.对于硝化能力强的土壤,由于施入的NH能够快速转化为NO,喜铵作物对铵态氮肥和硝态氮肥的吸收率差异很小(19.5% vs. 18.6%),而在硝化能力弱的土壤种植的喜铵作物对铵态氮肥的吸收率则显著高于硝态氮肥(31.0% vs. 22.3%)[39].同样,喜硝作物种植在硝化能力强的土壤,其对铵态氮肥的吸收率要明显高于硝化能力弱的土壤[39].可见,土壤氮转化特征决定的土壤无机氮主导形态与作物氮形态喜好的契合程度对作物氮肥利用率具有重要影响. ...
... [39].同样,喜硝作物种植在硝化能力强的土壤,其对铵态氮肥的吸收率要明显高于硝化能力弱的土壤[39].可见,土壤氮转化特征决定的土壤无机氮主导形态与作物氮形态喜好的契合程度对作物氮肥利用率具有重要影响. ...
... [39].可见,土壤氮转化特征决定的土壤无机氮主导形态与作物氮形态喜好的契合程度对作物氮肥利用率具有重要影响. ...
... 因此,在制定氮肥调控措施时需要考虑以上这种契合关系.例如,使用硝化抑制是调节土壤氮转化速率和无机氮形态的主要手段之一,当前更被作为减少温室气体N2O排放的有效措施而被科学工作者推荐.但是,大量的试验表明,硝化抑制剂虽然减少N2O排放的效果显著,然而常常不能增加作物对氮的吸收利用,减少氮素损失.基于以上契合理论,使用硝化抑制剂对作物氮吸收利用的效果取决于它是否能提高土壤无机氮形态与作物喜好的氮形态的契合程度,若使二者的氮形态更加吻合,则氮肥吸收利用率增加,损失率减少;反之,则不仅不会增加作物对氮的利用率,还会减少氮素损失,相反,将使氮利用率下降,损失率增加.Zhang等[39]观测到,使用硝化抑制剂不仅未提高喜硝作物黄瓜氮肥吸收率,减少氮素损失,甚至在一定程度上降低了氮吸收,增加了氮损失;相反,对于喜氮的马铃薯,使用硝化抑制剂无论在硝化作用弱的红壤还是在硝化作用强的碱性紫色土,均一定程度地提高了氮肥利用率,减少了氮损失. ...
Conifer root discrimination against soil nitrate and the ecology of forest succession
1
1997
... 但是,也有一些研究与以上结论不一致,认为氮肥形态和植物氮形态喜好的契合程度对氮肥的吸收以及作物产量没有影响[36,40].我们认为一个重要的原因是这些研究没有考虑土壤氮转化过程对于施入氮肥形态的影响.通过本文第4部分的阐述,我们已经明确了无论是土壤有机氮矿化产生的氮,还是施入到土壤的氮肥,都会在土壤中发生转化,残留在土壤中的无机氮最终将以土壤氮转化特性决定的形态存在,因此氮转化过程特点会明显影响氮肥形态和植物氮形态喜好的契合程度.Zhang等[39]发现,在15N标记的NH施入土壤5天后,硝化作用弱的土壤中有56%~75%仍然以NH形态存在,NO仅占2%;而硝化作用强的土壤正相反,只有不到2%的15N 以NH形态存在,NO比例高达31%~45%.对于硝化能力强的土壤,由于施入的NH能够快速转化为NO,喜铵作物对铵态氮肥和硝态氮肥的吸收率差异很小(19.5% vs. 18.6%),而在硝化能力弱的土壤种植的喜铵作物对铵态氮肥的吸收率则显著高于硝态氮肥(31.0% vs. 22.3%)[39].同样,喜硝作物种植在硝化能力强的土壤,其对铵态氮肥的吸收率要明显高于硝化能力弱的土壤[39].可见,土壤氮转化特征决定的土壤无机氮主导形态与作物氮形态喜好的契合程度对作物氮肥利用率具有重要影响. ...
Gross nitrogen mineralization-, immobilization-, and nitrification rates as a function of soil C/N ratio and microbial activity
1
2003
... 土壤氮转化过程的速率大多是参与反应的底物浓度的函数,例如,土壤有机氮的初级矿化速率与土壤有机氮含量呈正相关,初级硝化速率与NH含量呈正相关,反硝化速率与NO含量呈正相关[41,42].植物对无机氮的吸收,降低参与转化的氮底物含量,因而,改变其参与的转化过程的速率,包括土壤微生物的同化速率.当氮肥施入土壤后,土壤微生物和作物会竞争吸收施入的氮.研究表明,在氮丰富的土壤中,短时间(小时时间尺度)内土壤微生物的竞争能力强于作物吸收;但较长时间(天时间尺度)内,作物的吸收竞争能力强于土壤微生物[43].在氮丰富的土壤上种植喜好吸收NO的作物时,如果施用铵态氮肥,硝化微生物将NH硝化为NO的过程具有对NH最强的竞争力[43].由此可见,土壤氮的供应和保持能力和作物喜好吸收的氮形态可能是影响作物与土壤微生物竞争氮的重要因素. ...
Gross N transformation rates and net N mineralisation rates related to the C and N contents of soil organic matter fractions in grassland soils of different age
1
2004
... 土壤氮转化过程的速率大多是参与反应的底物浓度的函数,例如,土壤有机氮的初级矿化速率与土壤有机氮含量呈正相关,初级硝化速率与NH含量呈正相关,反硝化速率与NO含量呈正相关[41,42].植物对无机氮的吸收,降低参与转化的氮底物含量,因而,改变其参与的转化过程的速率,包括土壤微生物的同化速率.当氮肥施入土壤后,土壤微生物和作物会竞争吸收施入的氮.研究表明,在氮丰富的土壤中,短时间(小时时间尺度)内土壤微生物的竞争能力强于作物吸收;但较长时间(天时间尺度)内,作物的吸收竞争能力强于土壤微生物[43].在氮丰富的土壤上种植喜好吸收NO的作物时,如果施用铵态氮肥,硝化微生物将NH硝化为NO的过程具有对NH最强的竞争力[43].由此可见,土壤氮的供应和保持能力和作物喜好吸收的氮形态可能是影响作物与土壤微生物竞争氮的重要因素. ...
Short-term competition between crop plants and soil microbes for inorganic N fertilizer
2
2010
... 土壤氮转化过程的速率大多是参与反应的底物浓度的函数,例如,土壤有机氮的初级矿化速率与土壤有机氮含量呈正相关,初级硝化速率与NH含量呈正相关,反硝化速率与NO含量呈正相关[41,42].植物对无机氮的吸收,降低参与转化的氮底物含量,因而,改变其参与的转化过程的速率,包括土壤微生物的同化速率.当氮肥施入土壤后,土壤微生物和作物会竞争吸收施入的氮.研究表明,在氮丰富的土壤中,短时间(小时时间尺度)内土壤微生物的竞争能力强于作物吸收;但较长时间(天时间尺度)内,作物的吸收竞争能力强于土壤微生物[43].在氮丰富的土壤上种植喜好吸收NO的作物时,如果施用铵态氮肥,硝化微生物将NH硝化为NO的过程具有对NH最强的竞争力[43].由此可见,土壤氮的供应和保持能力和作物喜好吸收的氮形态可能是影响作物与土壤微生物竞争氮的重要因素. ...
... [43].由此可见,土壤氮的供应和保持能力和作物喜好吸收的氮形态可能是影响作物与土壤微生物竞争氮的重要因素. ...
Roots, nitrogen transformations, and ecosystem services
2
2008
... 根际土壤氮转化过程不同于非根际土壤[44].植物根际反馈作用于土壤氮转化过程和速率的驱动因素极为复杂,涉及生物(土壤微生物生物量及其群落结构、菌根真菌等)、生物化学(酶、生物活性物质等)、土壤物理性质(土壤通气性、团聚体粒径组成及质量等)和化学性质(pH、有机碳源、营养元素含量等)等各个方面[45,46].一般情况下,根际土壤具有较高的有机氮初级矿化速率,但不一定具有较高的有机氮净矿化速率.植物对土壤有机氮净矿化速率的影响在很大程度上取决于根分泌物及凋落物的C/N,若根分泌物及凋落物的C/N高,则土壤微生物与植物竞争有效氮,降低土壤有机氮净矿化速率;反之则可能提高土壤有机氮净矿化速率[45].植物对硝化作用的影响则在很大程度上取决于植物喜好的无机氮形态,若植物喜好吸收NO,则可能促进土壤硝化作用,反之则可能抑制硝化作用[47].植物通过刺激包括硝化微生物在内的根际微生物生长,促进根际土壤硝化作用,通过降低根际土壤pH和分泌具有硝化抑制作用的生物硝化抑制剂而降低土壤硝化速率[44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
... [44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
Feedback in the plant-soil system
2
2005
... 根际土壤氮转化过程不同于非根际土壤[44].植物根际反馈作用于土壤氮转化过程和速率的驱动因素极为复杂,涉及生物(土壤微生物生物量及其群落结构、菌根真菌等)、生物化学(酶、生物活性物质等)、土壤物理性质(土壤通气性、团聚体粒径组成及质量等)和化学性质(pH、有机碳源、营养元素含量等)等各个方面[45,46].一般情况下,根际土壤具有较高的有机氮初级矿化速率,但不一定具有较高的有机氮净矿化速率.植物对土壤有机氮净矿化速率的影响在很大程度上取决于根分泌物及凋落物的C/N,若根分泌物及凋落物的C/N高,则土壤微生物与植物竞争有效氮,降低土壤有机氮净矿化速率;反之则可能提高土壤有机氮净矿化速率[45].植物对硝化作用的影响则在很大程度上取决于植物喜好的无机氮形态,若植物喜好吸收NO,则可能促进土壤硝化作用,反之则可能抑制硝化作用[47].植物通过刺激包括硝化微生物在内的根际微生物生长,促进根际土壤硝化作用,通过降低根际土壤pH和分泌具有硝化抑制作用的生物硝化抑制剂而降低土壤硝化速率[44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
... [45].植物对硝化作用的影响则在很大程度上取决于植物喜好的无机氮形态,若植物喜好吸收NO,则可能促进土壤硝化作用,反之则可能抑制硝化作用[47].植物通过刺激包括硝化微生物在内的根际微生物生长,促进根际土壤硝化作用,通过降低根际土壤pH和分泌具有硝化抑制作用的生物硝化抑制剂而降低土壤硝化速率[44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
Contribution of exudates, arbuscular mycorrhizal fungi and litter depositions to the rhizosphere priming effect induced by grassland species
1
2015
... 根际土壤氮转化过程不同于非根际土壤[44].植物根际反馈作用于土壤氮转化过程和速率的驱动因素极为复杂,涉及生物(土壤微生物生物量及其群落结构、菌根真菌等)、生物化学(酶、生物活性物质等)、土壤物理性质(土壤通气性、团聚体粒径组成及质量等)和化学性质(pH、有机碳源、营养元素含量等)等各个方面[45,46].一般情况下,根际土壤具有较高的有机氮初级矿化速率,但不一定具有较高的有机氮净矿化速率.植物对土壤有机氮净矿化速率的影响在很大程度上取决于根分泌物及凋落物的C/N,若根分泌物及凋落物的C/N高,则土壤微生物与植物竞争有效氮,降低土壤有机氮净矿化速率;反之则可能提高土壤有机氮净矿化速率[45].植物对硝化作用的影响则在很大程度上取决于植物喜好的无机氮形态,若植物喜好吸收NO,则可能促进土壤硝化作用,反之则可能抑制硝化作用[47].植物通过刺激包括硝化微生物在内的根际微生物生长,促进根际土壤硝化作用,通过降低根际土壤pH和分泌具有硝化抑制作用的生物硝化抑制剂而降低土壤硝化速率[44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
Potential nitrification as an indicator of preferential uptake of ammonium or nitrate
1
2000
... 根际土壤氮转化过程不同于非根际土壤[44].植物根际反馈作用于土壤氮转化过程和速率的驱动因素极为复杂,涉及生物(土壤微生物生物量及其群落结构、菌根真菌等)、生物化学(酶、生物活性物质等)、土壤物理性质(土壤通气性、团聚体粒径组成及质量等)和化学性质(pH、有机碳源、营养元素含量等)等各个方面[45,46].一般情况下,根际土壤具有较高的有机氮初级矿化速率,但不一定具有较高的有机氮净矿化速率.植物对土壤有机氮净矿化速率的影响在很大程度上取决于根分泌物及凋落物的C/N,若根分泌物及凋落物的C/N高,则土壤微生物与植物竞争有效氮,降低土壤有机氮净矿化速率;反之则可能提高土壤有机氮净矿化速率[45].植物对硝化作用的影响则在很大程度上取决于植物喜好的无机氮形态,若植物喜好吸收NO,则可能促进土壤硝化作用,反之则可能抑制硝化作用[47].植物通过刺激包括硝化微生物在内的根际微生物生长,促进根际土壤硝化作用,通过降低根际土壤pH和分泌具有硝化抑制作用的生物硝化抑制剂而降低土壤硝化速率[44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
A colourimetric microplate assay for simple, high throughput assessment of synthetic and biological nitrification inhibitors
1
2017
... 根际土壤氮转化过程不同于非根际土壤[44].植物根际反馈作用于土壤氮转化过程和速率的驱动因素极为复杂,涉及生物(土壤微生物生物量及其群落结构、菌根真菌等)、生物化学(酶、生物活性物质等)、土壤物理性质(土壤通气性、团聚体粒径组成及质量等)和化学性质(pH、有机碳源、营养元素含量等)等各个方面[45,46].一般情况下,根际土壤具有较高的有机氮初级矿化速率,但不一定具有较高的有机氮净矿化速率.植物对土壤有机氮净矿化速率的影响在很大程度上取决于根分泌物及凋落物的C/N,若根分泌物及凋落物的C/N高,则土壤微生物与植物竞争有效氮,降低土壤有机氮净矿化速率;反之则可能提高土壤有机氮净矿化速率[45].植物对硝化作用的影响则在很大程度上取决于植物喜好的无机氮形态,若植物喜好吸收NO,则可能促进土壤硝化作用,反之则可能抑制硝化作用[47].植物通过刺激包括硝化微生物在内的根际微生物生长,促进根际土壤硝化作用,通过降低根际土壤pH和分泌具有硝化抑制作用的生物硝化抑制剂而降低土壤硝化速率[44].筛选和分离植物源的生物硝化抑制剂,用于替代化学合成的硝化抑制剂,被看作是缓解施用化学合成硝化抑制剂造成的生态环境负面效应的有效途径之一[48]. ...
Research progress of ‘4R’ nutrient stewardship in sustainable agricultural development
1
2015
... 随着研究方法、仪器分析技术以及分子生物学技术的进步,我们对于土壤氮转化过程的认识不断深入.但是,已有的关于土壤氮转化过程的研究,多数是在室内或原位不考虑植物生长的条件下进行的.虽然这些研究结果对于认识土壤氮转化过程的特点、产物、影响因素以及微生物机制等发挥了很大的作用,但事实上,土壤氮转化过程和植物之间存在密切的反馈作用.由于现在缺少同步测定土壤—植物系统中土壤氮转化速率和植物氮吸收速率的方法,关于植物对土壤氮转化反馈作用的研究始终停留在定性探讨的阶段,难以深入认识植物对土壤氮转化反馈作用的复杂机理.在理论上,限制了植物对土壤氮转化过程反馈作用机理的认识及对反馈作用的定量描述;在氮肥管理的生产实际上,难以制定出针对土壤—植物相互作用特点的氮肥管理策略,贯彻落实“4R养分管理”概念,即选择正确的肥料品种(Right source)、采用正确的肥料用量(Right rate)、在正确的时间(Right time)将肥料施用在正确的位置上(Right place)[49].现在急需建立能够同时定量土壤—植物系统中土壤氮初级转化速率和作物氮吸收NO和NH速率的方法,以实现定量化分析土壤氮转化过程与植物间的反馈作用关系.针对这一问题,作者所在团队申请了国家自然科学基金重点项目“植物对土壤氮转化的反馈作用及其机理研究”(批准号:41830642),并获得资助.本项目将建立和完善一种同时定量土壤—植物系统中土壤氮初级转化速率和作物氮吸收NO和NH速率的方法,探讨不同土壤性质和氮形态喜好植物条件下,植物对土壤氮转化的反馈作用及其作用机理,深入认识更接近于自然状态的土壤—植物系统中土壤氮的转化规律及其影响因素,探讨植物对土壤氮转化反馈作用的机理,研究结果可以深化我们对土壤—植物系统中土壤氮转化特征的认识,为根据作物—土壤系统氮转化特征制订氮肥管理方案提供理论依据. ...
农业可持续发展中的“4R”养分管理研究进展
1
2015
... 随着研究方法、仪器分析技术以及分子生物学技术的进步,我们对于土壤氮转化过程的认识不断深入.但是,已有的关于土壤氮转化过程的研究,多数是在室内或原位不考虑植物生长的条件下进行的.虽然这些研究结果对于认识土壤氮转化过程的特点、产物、影响因素以及微生物机制等发挥了很大的作用,但事实上,土壤氮转化过程和植物之间存在密切的反馈作用.由于现在缺少同步测定土壤—植物系统中土壤氮转化速率和植物氮吸收速率的方法,关于植物对土壤氮转化反馈作用的研究始终停留在定性探讨的阶段,难以深入认识植物对土壤氮转化反馈作用的复杂机理.在理论上,限制了植物对土壤氮转化过程反馈作用机理的认识及对反馈作用的定量描述;在氮肥管理的生产实际上,难以制定出针对土壤—植物相互作用特点的氮肥管理策略,贯彻落实“4R养分管理”概念,即选择正确的肥料品种(Right source)、采用正确的肥料用量(Right rate)、在正确的时间(Right time)将肥料施用在正确的位置上(Right place)[49].现在急需建立能够同时定量土壤—植物系统中土壤氮初级转化速率和作物氮吸收NO和NH速率的方法,以实现定量化分析土壤氮转化过程与植物间的反馈作用关系.针对这一问题,作者所在团队申请了国家自然科学基金重点项目“植物对土壤氮转化的反馈作用及其机理研究”(批准号:41830642),并获得资助.本项目将建立和完善一种同时定量土壤—植物系统中土壤氮初级转化速率和作物氮吸收NO和NH速率的方法,探讨不同土壤性质和氮形态喜好植物条件下,植物对土壤氮转化的反馈作用及其作用机理,深入认识更接近于自然状态的土壤—植物系统中土壤氮的转化规律及其影响因素,探讨植物对土壤氮转化反馈作用的机理,研究结果可以深化我们对土壤—植物系统中土壤氮转化特征的认识,为根据作物—土壤系统氮转化特征制订氮肥管理方案提供理论依据. ...





 甘公网安备62010202000687
甘公网安备62010202000687{kind=link}
{kind=link}
{kind=link}
{kind=link}