{kind=link}
{kind=link}
中国陆地生态系统植硅体碳汇研究进展
[潘文杰1, 2  , 杨孝民
, 杨孝民1 , 张晓东1 , 李自民3 , 杨石磊1 , 吴云涛1 , 郝倩1 , 宋照亮1, * ]
, 杨孝民]
|
|


作者简介:潘文杰(1990-),男,河南新郑人,硕士研究生,主要从事植硅体碳汇研究.E-mail:pwj13145@163.com
植硅体碳(Phytolith-Occluded Carbon,PhytOC)是一种相对稳定的碳组分,在生物地球化学碳循环和减缓全球变暖中扮演着重要角色。在总结前人研究的基础上,论述了植硅体碳的形成机制和其碳汇能力的影响因素,综述了当前中国陆地生态系统植硅体碳汇的研究成果,探讨了中国陆地生态系统植硅体碳汇的调控机制,最后对未来中国陆地生态系统植硅体碳汇的研究方向进行了展望。气候、地表植被类型、土壤环境及植硅体自身化学组份等诸多因素都将直接或间接影响植硅体的碳汇能力。中国草地、农田、森林、湿地和灌丛生态系统植硅体碳产生速率分别为(0.6±0.1)×106,(4.9±1.7)×106,(1.9±0.4)×106,(0.6±0.5)×106和(1.3±0.3)×106 t CO2 /a。含硅材料施加、高硅植物栽培和传统的提高植物地上净初级生产力等措施均可显著提高中国陆地生态系统植硅体碳汇潜力。今后应进一步研究不同植物产生植硅体碳的机理,加强不同陆地生态系统中植物地下部分植硅体碳汇能力的研究,对不同陆地生态系统土壤植硅体碳汇量进行量化,并提出更加全面、经济、合理的管理措施以提高植硅体碳汇量。
, Yang Xiaomin
*Corresponding author:Song Zhaoliang (1978-), male, Yiwu City, Zhejiang Province, Professor. Research areas include silicon-carbon coupled biogeochemical cycle.E-mail:songzhaoliang78@163.com
First author:Pan Wenjie (1990-), male, Xinzheng City, He’nan Province, Master student. Research areas include phytolith carbon squestration.E-mail:pwj13145@163.com
Phytolith-Occluded Carbon (PhytOC), a relatively stable carbon fraction, plays an important role in biogeochemical carbon cycle and mitigation of global warming. The formation mechanisms of PhytOC, the influence factors of phytolith carbon sequestration, the advances in study of phytolith carbon sequestration, and the management for enhancing the potential of phytolith carbon sequestration in terrestrial ecosystems of China were summarized in this review. Finally, future researches on phytolith carbon sequestration in terrestrial ecosystems of China were prospected. Climates, vegetation types, soil circumstances, the chemical compositions of the phytoliths and other factors will directly or indirectly affect the potential of phytolith carbon sequestration. In China, the PhytOC production quantity in grassland, cropland, forest, wetland and shrub ecosystems is (0.6±0.1)×106,(4.9±1.7)×106,(1.9±0.4)×106,(0.6±0.5)×106 and (1.3±0.3)×106 t CO2/a, respectively. Application of silicon-containing fertilizer, cultivation of high-silicon plant, and traditional enhancement of the plant net primary productivity can significantly improve the potential of phytolith carbon sequestration in terrestrial ecosystems of China. Future studies should focus more on ①the mechanisms of phytolith formation in different plants, ②the phytolith carbon sequestration in underground parts of plants from different terrestrial ecosystems, ③the quantification of soil PhytOC in different terrestrial ecosystems. Furthermore, more comprehensive, economical and reasonable management practices of improving the potential of phytolith carbon sequestration should also be further studied in different terrestrial ecosystems.
自工业革命以来, 大量化石燃料(煤炭、石油等)的使用造成了大气中CO2浓度和全球气温的不断升高, 导致了一系列环境问题[1]。因此, 寻找一种长期稳定的碳汇机制受到越来越多学者的关注[2~4]。陆地生态系统生物地球化学碳汇作为一种有效降低大气CO2浓度的手段, 在全球气候变化和碳循环过程中发挥着重要作用[5, 6]。植硅体(phytoliths), 也称植物蛋白石, 是植物通过根系吸收土壤中的生物有效硅后, 经过硅化作用, 沉淀在植物细胞、组织(如细胞壁、内腔等)中的非晶质二氧化硅(SiO2)[7~9]。在大多数植物体内, 90%以上的SiO2以植硅体的形式沉积[9]。在植硅体的形成过程中, 部分有机碳通常会被植硅体包裹, 形成植硅体包裹碳, 简称植硅体碳(phytolith-occluded carbon, PhytOC)[4, 10]。由于植硅体具有抗分解、耐腐蚀等特性, 在植物死亡分解以后, 植硅体碳会随着植硅体在土壤或沉积物中长期保存[2, 3]。有学者认为植硅体碳很有可能是全球碳失汇的重要组成部分[8], 因此, 近年来关于植硅体碳汇的研究逐渐展开。本文在总结前人研究基础上, 论述了植硅体碳的形成机制、特性, 综述了目前我国陆地生态系统植硅体碳汇的研究成果, 总结了植硅体碳汇的调控机制, 并对今后植硅体碳汇领域的研究方向进行展望。
植硅体主要由SiO2、水和有机碳组成, 其中SiO2含量可以达到66%~91%, 水的含量为1%~12%, 有机碳含量为0.2%~5.8%, 此外还有少量的氮(N)、磷(P)、钙(Ca)、铁(Fe)、铝(Al)、钾(K)和镁(Mg)等元素[11~13], 这些元素的含量为0.1%~9%[14, 15]。不同植物中植硅体的含量有很大差异[16~18]。蕨类植物通常会利用植物细胞壁中的网状SiO2来增强机械组织的支撑力[17], 因此其体内的植硅体含量比较高。裸子植物产生的植硅体通常少于被子植物, 而在被子植物中, 单子叶植物通常比双子叶植物积累更多的植硅体[3, 16]。如在大多数双子叶植物中, 植硅体含量通常少于0.5%, 而在单子叶的莎草科和禾本科植物中, 植硅体含量可以达到10%~15%[17, 18]。由于具有较强的硅吸收能力, 竹子、水稻和甘蔗等均属于典型的富植硅体植物[2, 19, 20]。通过高分辨率电子显微镜分析发现, 植硅体内的硅主要以Si-O-Si共价键结合的单硅酸形式存在[21]。Carter[22]提出了一种植硅体碳形成模型:具有液泡结构的植物细胞在生长过程中, 其细胞液中通常会包含硅酸; 在细胞与胞外环境进行液体交换的过程中, 硅酸被保留下来并开始浓缩; 当硅酸浓度持续增升高的时候, 其他细胞器或有机物可能会以某种特定的方式进入液泡中; 当液泡中的硅酸浓度上升到能聚合的程度后就会聚合成包裹着上述细胞器或有机物的硅胶, 胞内压和其他条件会导致该硅胶具有特定的形状; 在细胞的衰老过程中, 细胞中的液体逐渐移出, 导致硅胶固化, 因此就形成了包裹着碳并具有特定形状的植硅体。
根据相关研究推测, 植硅体碳主要成分可能是碳水化合物、脂类物质、木质素和糖蛋白[13, 22, 23]。尽管目前的研究尚未发现植硅体碳中存在DNA等生物遗传信息[23], 但是由于植硅体的形状通常是由物种类别和外界环境条件所控制, 因此植硅体的形状组成和植硅体碳同位素定年对重建古植被、古气候、古环境等方面具有重要意义[22~25]。相对于陆地生态系统中其他形式的有机碳, 由于受到植硅体外壳的保护, 植硅体碳可以在土壤或者沉积物中保存千年甚至万年之久[8, 26]。Parr等[8]通过对土壤中植硅体碳的定年发现植硅体碳可以在古土壤中保存8 000年。Wilding[26]报道了植硅体碳可以在泥炭沉积物中保存13 300年。此外, 对土壤有机碳的研究发现, 经过2 000多年的分解, 植硅体碳的含量占到土壤总有机碳含量的82%[8]。因此, 植硅体碳汇作为一种长期稳定的生物地球化学碳汇机制引起了人们的重视[3, 8]。
植硅体碳受植硅体外壳的保护可以长时间保存在土壤中, 其稳定性取决于植硅体的稳定性。土壤中植硅体的生物地球化学稳定性受多种因素影响。Alexandre等[27]研究发现, 热带雨林凋落物中的植硅体只有7.5%可以稳定保存在土壤中, 土壤植硅体储量约为56.9 t/hm2; 而温带森林(温带人工林)土壤植硅体储量高达169.4 t/hm2[28]。在地表不同的植被类型土壤中, 针叶林中形成的植硅体在土壤中的稳定性通常比在草本植物中形成的要高[4]。在土壤条件方面, 随着土壤可利用水分的增多, 植硅体的溶解性也会随之增强[29]; 植硅体在酸性土壤环境如水稻土中保存较好, 不易受到侵蚀, 相反植硅体的溶解性在碱性土壤(pH> 8)中会增强[30~32], 植硅体的溶解速率与pH呈显著正相关[33]; Zhang等[34]研究发现不同母岩发育的土壤对植硅体稳定性产生显著影响。通过对植硅体化学组分的研究发现表面吸附有金属离子(如Al3+)的植硅体的抗分解能力相对较强[11]。因此, 气候、地表植被类型、土壤条件(pH、含水量、成土母质)及植硅体自身化学组分等诸多因素都将直接或间接地影响植硅体的碳汇能力。
草地是全球分布最广泛的植被类型之一。由于草地生态系统具有面积大、地表净生产力高和富硅植物多等特点, 在全球陆地生态系统植硅体碳汇研究中具有重要意义[35, 36]。中国草地面积约占中国陆地面积的1/3, 主要包括典型草原、草甸草原、荒漠草原、高寒草原、高寒草甸和其他草地等。Song等[36]根据目前中国各类型草原的分布面积、地表净初级生产力、优势草本植物的SiO2含量和植硅体平均含碳量, 在区域尺度上建立了中国草地植被植硅体碳汇的计量方法。中国草地植硅体碳的产生量约为0.6 × 106 t CO2/a[36]。由于各个类型草地之间的分布面积、地表净初级生产力、植被组成、气候、地形等差异很大, 因此各个类型草原的植硅体碳汇量不尽相同[36]。例如, 草甸草原的植硅体碳产生通量(1.35 kg/(hm2· a))是其他草原的2~5倍, 但由于其分布面积较小, 所以其对整个草地生态系统植硅体碳汇的贡献仅为7%(表1)。
| 表1 中国草地生态系统植硅体碳汇产生通量和产生量 Table 1 The production flux andquantity of PhytOC in Chinese grassland ecosystem |
Qi等[37]的研究结果显示典型草原植被地上部分的植硅体碳的产生通量为0.32 kg /(hm2· a), 其结果略低于Song等[36]的研究, 这可能是由于草地植被群落结构分布具有空间异质性。研究发现地表植物植硅体产生通量与地表净初级生产力之间呈现极显著的正相关[29, 32, 36], 因此地表净初级生产力对植硅体碳产生通量也具有重要的影响作用。与北美草地相比, 中国草地相对较低的地表初级净生产力导致中国草地植硅体碳产生通量(1.81 kg CO2 /(hm2· a))仅为北美草地的22%[29, 36]。Qi等[37]研究显示, 典型草原植物地下部分的植硅体碳产生通量(11.95 kg /(hm2· a))约为植物地上部分的40倍, 这表明草地地下部分植硅体碳汇具有巨大的潜力。在温带干旱和半干旱的地区, 多数草地的地下净初级生产力通常是地上净初级生产力的10倍左右[37, 38]。然而, 现阶段对草地地下部分植硅体的研究较少, 各类型草地地下部分植硅体产生通量目前还没有进行估算。因此, 加强对草地植物地下部分植硅体的研究对于精确评估草地生态系统植硅体碳汇潜力十分重要。
作为最大的农作物生产国之一, 中国约有160× 106 hm2耕地[39]。Zuo等[40]通过对8个品种的粟和黍研究发现, 黍和粟的植硅体碳含量分别为1.36和1.29 g/kg, 并估算出中国旱作农业植硅体碳汇量为2.37× 106 t CO2/a。相关研究表明, 稻谷(21~144 g/kg)[20]、小麦(27~79 g/kg)[41]、玉米(37~67 g/kg)[42]的植硅体含量通常比其他作物高。稻谷、小麦和玉米种植面积大, 约占全国农田面积的一半以上(表2)。根据Song等[42]的研究, 我国甘蔗的植硅体碳产生通量(96 kg CO2 /(hm2· a))明显高于其他农作物, 但是甘蔗属于热带作物, 种植面积仅为1.9 × 106 hm2, 因此其植硅体碳产生量仅为三大粮食作物(稻谷、玉米和小麦)的9%~21%(表2)。由于水稻、小麦和玉米的种植范围广、植硅体含量高, Song等[42]根据水稻、小麦和玉米的植硅体产生量估算了我国农田生态系统的植硅体碳汇量, 研究结果显示, 我国稻谷的植硅体碳产生通量(68 kg CO2/(hm2· a))高于玉米(44 kg CO2/(hm2· a))和小麦(38 kg CO2/(hm2· a))(表2)。根据2015年国家统计局公布的最新数据, 中国中南、华东地区的水稻和小麦的种植面积较其他地区大, 而除东北和华北以外, 各地玉米种植面积差别不大。因此, 从区域种植模式分析, 华东和中南地区的农田植硅体碳汇量比其他地方高, 分别为1.23 × 106 t CO2 /a和1.19 × 106 t CO2 /a(图1)。
| 表2 中国主要农作物植硅体碳产生通量和产生量 Table 2 The production flux and quantity of PhytOC in main arable crops in China |
根据近20年中国农田生态系统植硅体碳汇量的动态变化, 在时间尺度上可将中国农田生态系统植硅体碳汇分为3个阶段(图2)。1996— 1999年, 玉米、小麦和稻谷3种主要作物的植硅体总碳汇量变化不大(4.34× 106 ~4.36× 106 t CO2/a)。1999— 2003年, 由于种植面积减少, 植硅体碳汇量从4.36× 106 t CO2/a迅速下降到3.70× 106 t CO2/a。2003— 2015年, 植硅体碳汇量又迅速增加到4.65× 106 t CO2 /a, 这主要是因为种植面积的增加和作物产量的增加, 尤其是玉米产量的增加。
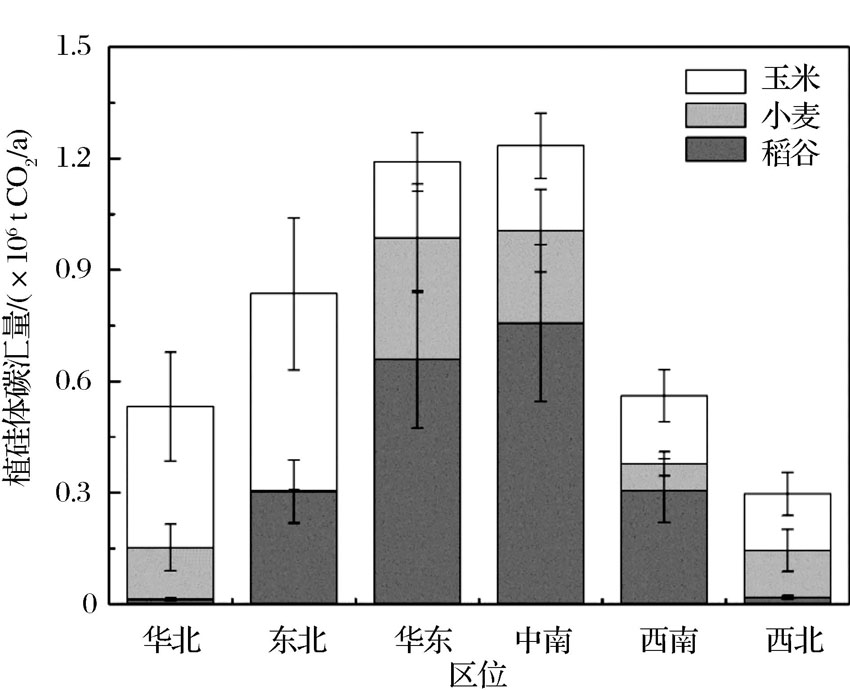 | 图1 2015年中国不同地区3种农作物的植硅体碳汇量 数据来源于国家统计局和参考文献[42]Fig.1 PhytOC production quantity of three arable crops in different regions of China in 2015 Data from National Bureau of Statistics of the People’ s Republic of China and references[42] |
森林是世界上最重要的生态系统之一。与草地生态系统相比, 大部分森林凋落物的植硅体含量较低[4, 43, 44], 但是森林生态系统具有更高的地表净初级生产力[45], 其植硅体产生量可占陆地生态系统植硅体产生量的20%以上[4, 29, 35]。
Song等[45]根据气候和植被类型, 选择中国8种不同的典型森林生态系统, 估算了中国不同典型森林生态系统植硅体碳产生通量和产生量(表3)。研究结果表明, 亚热带和热带竹林植硅体碳产生通量最高, 这与竹子较高的植硅体含量有关; 寒温带和温带针叶林、亚热带和热带针叶林则最低, 这与针叶林中较低的植硅体含量有关[45, 46]。与竹林相比, 针叶林的植硅体含量(11.9~13.2 g/kg)明显低于竹林的植硅体含量(101 g/kg)。热带雨林虽然有最大的地表净初级生产力(10.89 t/(hm2· a)), 但热带雨林在中国分布面积较少[47], 所以我国热带雨林的植硅体碳产生量仅为0.03 × 106 t CO2 /a; 虽然竹林的地表净初级生产力(7.37 t /(hm2· a))低于热带雨林, 但由于我国竹林植硅体含量高, 因此我国竹林植硅体碳产生量高达0.59 × 106 t CO2/a(表3)。据估算, 中国森林生态系统植被的植硅体碳汇量为1.7 × 106 t CO2 /a, 其中亚热带和热带竹林植硅体碳汇约为0.53× 106 t CO2 /a, 贡献约30%的植硅体碳汇, 这充分说明了竹林植硅体碳汇在森林生态系统植硅体碳汇中的重要性[45]。
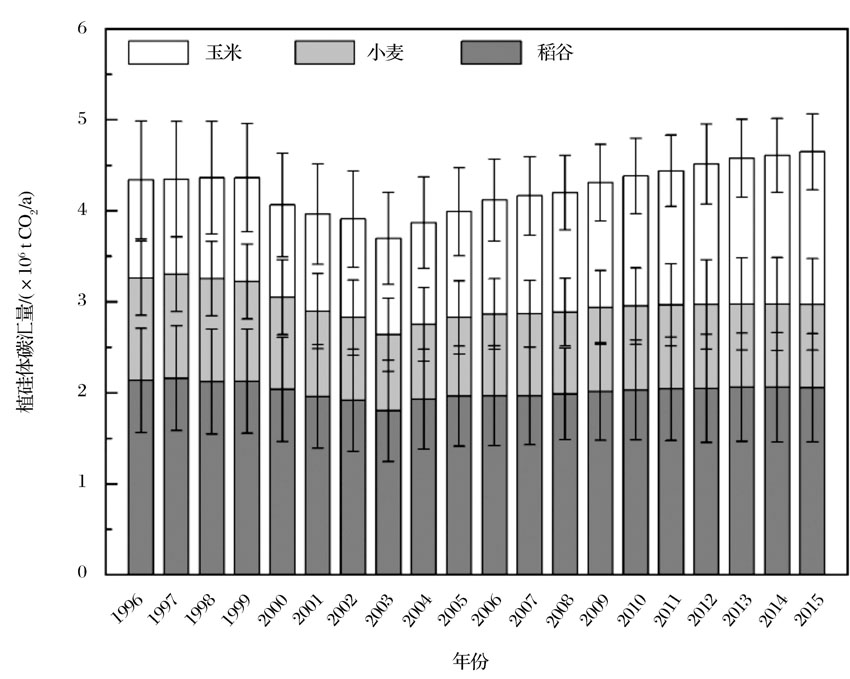 | 图2 1996— 2015年中国3种农作物植硅体碳汇量 数据来源于国家统计局和参考文献[42]Fig.2 PhytOC production quantity of three arable crops in China between 1996 and 2015 Data from National Bureau of Statistics of the People’ s Republic of China and reference[42] |
| 表3 中国主要森林类型植硅体碳产生通量和产生量 Table 3 The PhytOC production flux and quantity of main forest types in China |
此后, 应雨骐等[48]对中国亚热带3种森林类型的7个树种的植硅体碳汇能力进行了研究, 研究结果显示, 亚热带杉木林和马尾松林植硅体碳产生量分别为6.3 × 104 t CO2/a和1.3 × 104 t CO2 /a, 然而亚热带阔叶林植硅体碳平均产生量则高达24.7× 104 t CO2 /a。研究结果与Song 等[45]的报道相近(表 2), 并再次验证了针叶林的植硅体碳产生速率较低的观点。
除了草地、农田和森林生态系统外, Li 等[49]通过对西溪湿地植硅体碳汇的研究表明, 西溪湿地的植硅体碳产生通量(3~77 kg CO2 /(hm2· a))的变化范围较大, 并以此估算我国湿地的植硅体碳产生量为0.04× 106~1.05× 106 t CO2/a。相对于物质比较丰富的西溪湿地而言, 以芦苇为主的白洋淀地上部分植硅体碳产生通量差异性较小(48~54 kg CO2/(hm2· a))[50], 其湿地土壤(0~15 cm)中植硅体碳积累速率为5.49 t CO2 /a[50]。最近, Ru等[51]通过对中国北方温带灌丛植硅体碳汇的研究报道了中国灌丛生态系统植硅体碳汇量为1.28 × 106 t CO2/a。到目前为止, 国内对于其他生态系统(如荒漠生态系统)的植硅体碳汇报道比较少, 相关研究有待开展。
由表4可见, 中国主要陆地生态系统植硅体碳汇量差别很大。为了增加植硅体碳汇在减缓全球气候变暖中的作用, 应加大对植硅体碳汇的管控。例如, 为了提高中国草地生态系统植硅体碳汇量, 提高草地的地表净初级生产力是一种有效手段, 如避免过度放牧、减少收割、恢复退化草地和施肥等。由于相对较高的稻谷和玉米种植比例, 中国农田生态系统植硅体碳汇量(6.0 × 106 t CO2 /a), 约占世界耕地植硅体碳汇量的18%, 高于印度(3.6 × 106 t CO2/a)和美国(3.3 × 106 t CO2/a) [42]。最近的研究发现, 对稻田施用额外的硅肥, 如生物质炭[52]、富含硅的有机肥[53]、玄武岩岩粉[54]和钢渣硅肥[55]等, 可以显著提高稻田生态系统的植硅体碳产生通量。除此之外, 扩大高植硅体作物如水稻、甘蔗等的种植面积, 选择高植硅体碳产量的品种, 均会增加中国农田生态系统植硅体碳汇量[56, 57]。如上所述, 竹林为中国森林生态系统贡献了约30%的植硅体碳汇量, 然而不同竹子品种的植硅体含量差异很大, 在40~160 g/a变化[58]。中国现有竹林面积7.2 × 106 hm2, 且面积每年都在增长[59]。因此, 在今后的竹子造林和再造林过程中, 选择高植硅体含量的竹子品种替换掉低植硅体含量的品种, 将大大增加竹林的植硅体碳汇量, 进而增加中国森林生态系统碳汇量。李自民等[49]通过对西溪湿地生态系统的研究发现, 不同湿地植物植硅体产生通量差异较大。因此, 通过选择种植高植硅体碳产生量的湿地植物, 可以提高湿地生态系统植硅体碳汇潜力。
| 表4 中国主要陆地生态系统植硅体碳产生通量和产生量 Table 4 The PhytOC production flux and quantity of main terrestrial ecosystem in China |
中国草地、农田、森林、湿地和灌丛生态系统植硅体碳产生速率分别为(0.6± 0.1)× 106, (4.9± 1.7)× 106, (1.9± 0.4) × 106, (0.6± 0.5)× 106和(1.3± 0.3)× 106 t CO2/a。这显示出植硅体碳汇是一种有潜力的生物地球化学碳汇机制, 可有助于缓解全球气候变暖。尽管如此, 现阶段对整个陆地生态系统植硅体固碳量的量化研究较缺乏, 未来的研究将包括:
(1) 进一步研究不同植物产生植硅体碳的机理, 并研究植物类型、气候、地形等因素对植物植硅体的形状、含量和包裹碳量的影响。同时, 探讨植硅体形态和化学组成等因素对植硅体固碳速率的影响。
(2) 加强不同陆地生态系统中植物地下部分植硅体碳汇能力的研究, 进而估计其对整个生态系统碳汇的贡献度。
(3) 量化不同陆地生态系统中土壤植硅体碳汇量。
(4) 目前提出的管理措施主要针对提高植硅体碳汇能力, 缺乏经济、社会等方面的考虑, 需要综合研究更加全面、合理的管理措施以提高植硅体碳汇量。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|