

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
深海热液生态系统特征及其对极端微生物的影响
[张亮1, 2  , 秦蕴珊
, 秦蕴珊1 ]
, 秦蕴珊|
|


作者简介:张亮(1985-),男,山东曲阜人,助理研究员,主要从事海底热液活动研究.E-mail:zhangliang@qdio.ac.cn
深海是地球上最重要的极端环境之一,发育了数量巨大的极端微生物。它们独特的生存环境、生理结构、代谢机制和共生关系成为探讨生命起源及寻找外太空生命的关键。尽管从生物学角度对极端微生物已开展了大量研究,但深海热液系统对极端微生物演化的影响仍不甚清楚。因此,在总结深海极端的理化环境和地质环境特征的基础上,分析了海底热液活动的分布特征、形成机理及其对周围生物群落种类、分布和演替规律的巨大影响。重点探讨了热液环境下各种极端微生物的生命形式及其对深海营养物质循环和生态系统演化的重要意义。目前,极端环境与生命过程的研究仍处于初级阶段,亟待加强深海原位探测和分子生物学技术的研发以及多学科交叉研究。
First author: Zhang Liang (1985-), male, Qufu City, Shandong Province, Assistant Professor. Research areas include seafloor hydrothermal activity.E-mail:zhangliang@qdio.ac.cn
Deep-sea is one of the most important extreme environments on the earth. Numerous and diverse extremophiles thrive in this extreme environment, presenting distinctive physiological structure, metabolic mechanism and symbiosis relations, which provide new methods to study the origin of life and extraterrestrial life. Despite extensive studies on deep-sea extremophiles from the point of view of biology, the impacts of deep-sea hydrothermal activity on the evolution of extremophiles remain largely unknown. On the basis of summarizing features of the deep-sea ziphysicochemical and geological environment, the distribution and formation mechanism of submarine hydrothermal vents were analyzed, respectively. Hydrothermal vents have great effect on the distribution and succession of communities. Our discussion focused on the extreme life forms of microorganisms surviving in the hydrothermal ecosystem and their important significance for the nutrient cycling and ecosystem evolution. However, the research of life processes in extreme environments is still in the primary stage and more work is needed on the in-situ detection technique, molecular biology and interdisciplinary research.
1872— 1876年, 英国科学调查船“ 挑战者” 号划时代的科学考察揭开了近代深海大洋调查研究的序幕[1]。但直到近几十年, 深海调查研究才取得革命性的重大突破。如海底扩张与板块学说的提出, 从DSDP到ODP再到IODP大洋钻探计划的实施, 大洋中脊系统与海底下海洋的发现以及海底矿物资源的勘探与开发等, 都对科学和社会的发展起到了重要的作用。1977年, 科学家在东太平洋加拉帕斯裂谷发现了现代热液喷口, 以及在温度高达几百度的热液喷口处仍存活的大量生物群落(含细菌、古菌、真菌等)[2, 3], 被认为是20世纪后期最显著的科学发现之一, 与其相关的资源、环境问题和“ 黑暗食物链” 生命过程也成为当前深海研究的焦点[4], 并取得了一系列成果。对极端环境下的生物基因的开发利用, 已形成数十亿美元的产业规模[5]; 对极端环境生命过程的研究称为天体生物学(Astrobiology), 成为探讨生命起源及寻找外太空生命的关键[6, 7]。然而, 关于深海极端环境与生命过程的研究工作是近三四十年才开辟的新领域, 加之深海探测技术的限制, 仍面临着诸多困难。由于99%的深海微生物是不可培养的[8], 导致传统培养的方式无法很好地研究生物群落多样性。对于深海微生物(如细菌、古菌等)群落的分布、分类、多样性等特征仍缺乏系统认识[9]。因此, 本文在总结深海环境尤其是海底热液系统的各种极端特性的基础上, 分析了深海热液极端环境下生态环境和微生物分布特征, 并结合我国在深海极端环境取得的最新进展, 探讨深海热液生态系统对其周围微生物的影响及极端微生物研究的重要意义。
深海通常指水深大于1 000 m的海域, 占全球海洋体积的75%, 是地球上最为重要的极端环境之一[10, 11], 它具有物理上的极端(如温度、辐射、压力)和化学上的极端(如盐度、pH、氧含量等)。深海环境或叫深海极端环境是由多因子共同塑造的一个统一系统, 拥有深海平原、海山、热液、冷泉及深渊等特殊环境, 导致海底地形、理化因子的剧烈变化。从地球系统科学的理念来看, 深海底部是地球各圈层(岩石圈、水圈、生物圈)之间相互作用, 相互依赖和相互影响最为频繁, 最为活跃的地区[12]。
根据介质不同, 我们将深海极端环境划分为2种:深海理化环境和海底的地质环境。
2.2.1 深海的理化环境特征
由于海洋表层的透光层一般为130~150 m, 深海海底都是黑暗的。同时, 深海海底承受着巨大的静水压力, 最高可达110 MPa(10 m=1 atm)。深海底附近水温较低, 多为0~2 ℃, 但在海底热液喷口则存在另一个极端, 温度高达100~400 ℃, 热液喷口的pH值为3~8, 且富含H2S和重金属等有毒物质和气体[13], 缺乏氧气。可见, 深海是一个黑暗、高压、低温(热液喷口除外)、寡营养以及富含硫化物的极端环境[14]。
2.2.2 深海的地质环境
深海底的地质环境是最复杂、影响最大的主导环境因素, 同时也控制和影响其他环境因子的变化, 可分为大尺度环境因子和小尺度的环境因子。前者如洋中脊、俯冲带及板内次级板块的边界等, 后者如海底沉积物、局部海底地貌、深海热流、海底热液系统等。
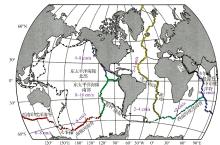
洋中脊是全球性海底山脉, 全长有8万多公里(据NOAA统计), 其中仅对1/3的洋中脊做过热液活动的调查[15]。洋中脊被认为是离散板块边界, 据其两侧向外扩张的速度可分为超快速扩张(> 14 cm/a)、快速扩张(8~14 cm/a)、中速扩张(5.5~8 cm/a)、慢速扩张(5.5~2.0 cm/a)、超慢速扩张(< 2 cm/a)[16](图1)。由于洋中脊处于扩张环境, 海底火山活动频繁, 热液系统十分发育。洋脊处热流值很高, 对上覆海水温度的升高影响很大, 同时也是地震多发地区。一般来说, 快速扩张的洋中脊地震频次高、强度小, 热液系统以分散、孤立的烟囱体为特征, 且高度一般小于15 m[17]; 中等扩张速率洋中脊上热液系统多表现为多期烟囱体堆积成不规则丘状, 如胡安· 德富卡海岭[18]; 而慢速扩张的洋中脊则通常地震频次低、强度大, 常发育大型的热液系统, 高度超过50 m, 如大西洋TAG热液活动区[19]。
俯冲带为汇聚板块边界, 当2个板块发生碰撞后, 厚度较薄、密度较大的板块会沿海沟俯冲到另一个板块之下, 形成岛弧、弧后盆地等, 并伴随着强烈的地震、火山活动, 如西太平洋沟弧盆体系(图1)。另外, 在板块内部及其边界处, 可划分出许多次级板块, 这些地带的火山、热液、地震都十分活跃, 对环境造成巨大影响。
在大尺度环境因子内部, 同时存在多种不同的小尺度环境因子。首先, 深海底的地形和地貌对局部海洋环境影响很大, 包括海山、深渊、峡谷、平原、高盐湖、泥火山、冷泉及热液喷口等。起伏不平的地貌对多尺度的海洋动力环境有重要影响[20]。海底地形也是决定热液喷口位置的重要因素之一, 如热液喷口常发育于中央裂谷内凸起部位[21]。其次, 海底沉积物性质对底栖生物有重要影响, 不同沉积时代和沉积条件下的生物种群存在着明显变化[22], 因而沉积层记录了环境的变化和生物种群的进化历史。最后, 海底热环境是深海地质环境的一个重要构成部分, 不同海区海底热流差异巨大。一般来说, 海底热流随大洋板块年龄增长而逐渐降低[23]。因此, 海底热液活动多发育于大洋中脊、弧后盆地扩张中心等构造活跃地区。
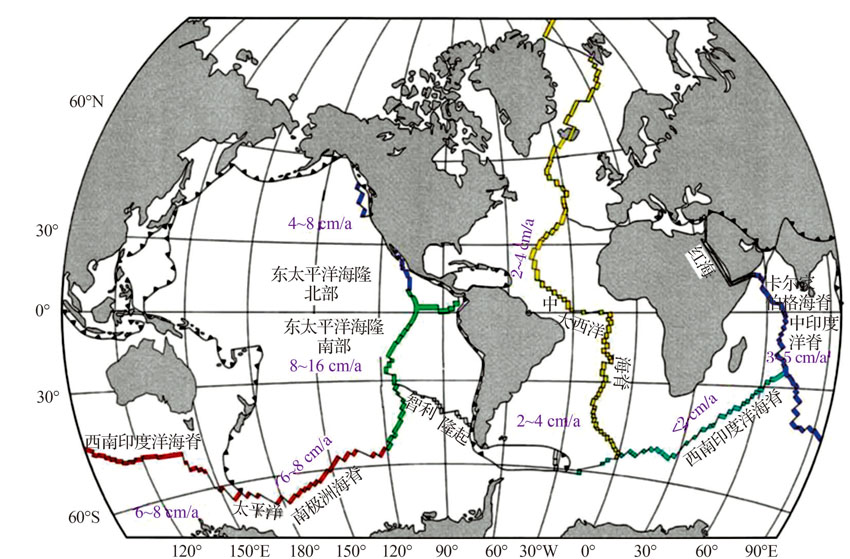 | 图1 全球洋中脊扩张速度示意图Fig.1 The spreading rate of the global mid ocean ridge system |
海底热液系统是深海极端环境的重要组成部分和典型代表, 汇集了多种极端物化环境且复杂多变。同时, 相比深海非热液区, 现代热液喷口及周围存在着丰度更高、更多样性的生物群落[24], 成为名副其实的海底沙漠中的“ 绿洲” [25]。此外, 热液喷口处的嗜热微生物的生存环境与地球早期环境类似, 因此被认为是研究生命起源的关键[7, 26]。
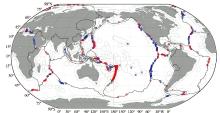
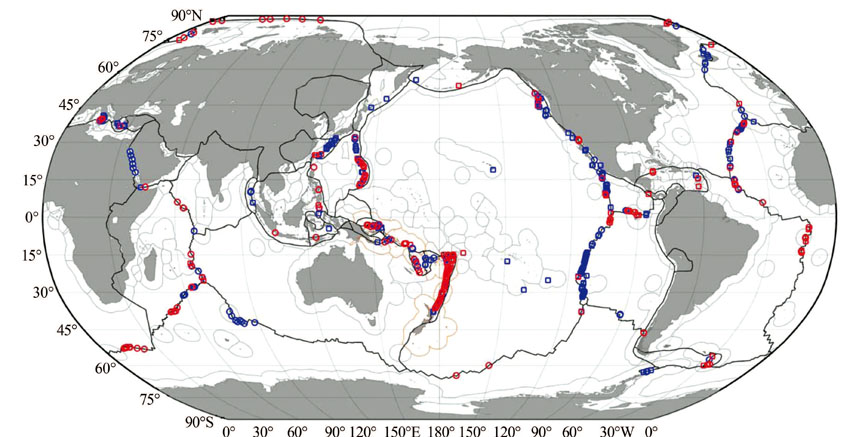
目前, 全球已发现的热液活动区约有521个, 大部分位于洋中脊中轴谷、弧后盆地、三连点和海底火山附近等[27](图2)。统计表明, 发育于洋中脊的热液活动区占总数的52%, 火山岛弧区占25%, 弧后盆地扩张中心占21%, 板内及其他构造环境约占2%。从分布的大洋看, 太平洋集中了约75%的热液活动区, 其次为大西洋(16%)、印度洋(3%)及其他海域(6%)[28]。
在地理位置上, 已发现的热液活动区主要集中在40° N~40° S的中、低纬度地区, 南极地区尚未发现[28]。热液活动区分布的水深范围跨度很大, 中国台湾龟山岛只有几十米, 而最深的Mid-Cayman扩张脊Beebe热液区位于水下4 960 m[29]。但全球热液活动区的水深大部分集中于1 300~3 700 m, 平均水深为2 500 m, 出现热液活动概率最高的水深范围为2 400~2 600 m, 相当于全球快速扩张脊的深度, 其次为2 200~2 400 m[28, 30]。此外, 热液活动发育频次与扩张脊扩张速率也密切相关, 在洋中脊表现为“ 双峰式” , 在全扩张速率为10~20 mm/a和60~70 mm/a 2个区间热液活动的频次最高; 而在弧后盆地扩张中心表现为“ 单峰式” , 在全扩张速率为50~60 mm/a出现次数最高, 向两端逐渐减少。根据两者之间统计规律预测, 扩张脊上约有900个热液活动区还未被发现, 并且绝大部分位于全速率小于60 mm/a的洋中脊和小于20 mm/a的弧后盆地扩张中心[31]。
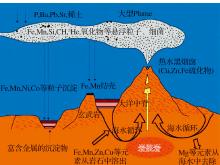
关于海底热液系统的形成机理, 当前普遍认可的观点是:在静水压力作用下, 海水沿地壳裂隙向下渗透, 受地热作用, 逐渐被加热的海水在地壳裂隙系统中的形成对流循环。同时, 在高压和高温环境下, 海水与地壳岩石发生反应, 最后以流体和气体的形式喷射出地表。在热液流体向上运移的过程中, 据热液流体与冷海水混合的早晚, 喷出地表时热液温度可为10~350 ℃[32], 已发现的温度最高可达407 ℃[33]。这些热液流偏酸性, 具还原性且富含重金属(包括Fe, Mn, Zn, Cu等)及无机分子(如 H2, CH4, H2S, CO2, CO等)。当它们喷射而出时, 溶解于其中的矿物质元素与冷海水接触, 就不断有金属硫化物沉淀形成。这些沉积物沉积在海底, 形成热液硫化物矿床, 有时会形成高达 10~15 m 的海底“ 黑烟囱” [34], 及其他不同类型的热液喷口构造[35](图3)。通过海水在大洋地壳裂隙的循环流动, 地热能被转化为化学能, 储存在还原性的无机化合物中, 为喷口生物群落的繁殖提供了基本的物质条件[36]。
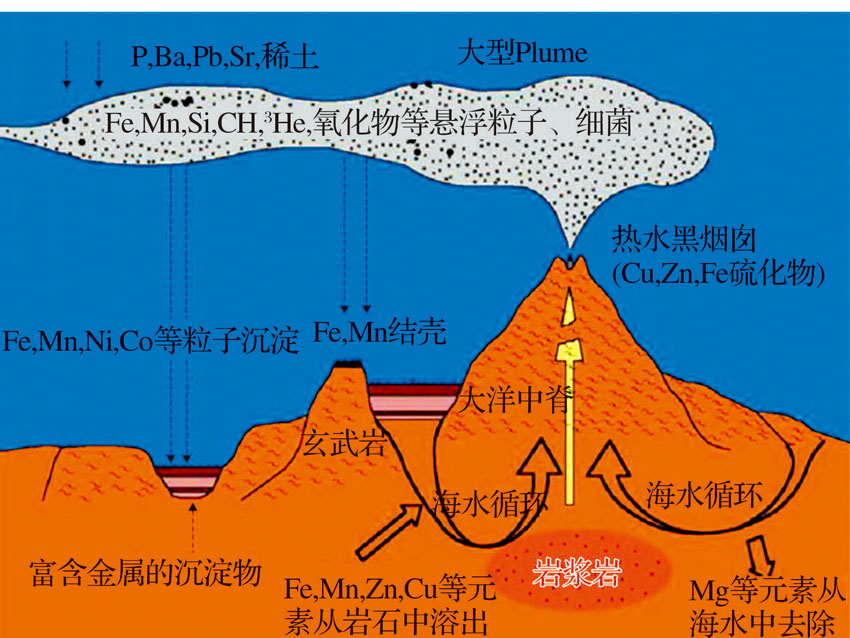 | 图3 热液系统形成过程和机理的示意图Fig.3 Schematic diagram offormation process and mechanism of hydrothermal vents |
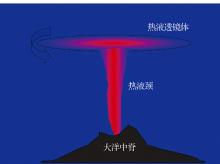
3.3.1 异常水体— — 热液柱
热液柱与周围海水表现出明显的异常特征。一是热液柱的化学成分异常, 如CH4, 3He和Mn等元素含量的异常增高(表1); 二是热液柱的CTDT(Chlorine, Temperature, Density and Transmittance)异常, 即海水的温度、盐度、密度和透光度发生异常。
| 表1 热液柱化学成分与正常海水对比[37] Table 1 The comparison of chemical composition between hydrothermal plume and normal sea water[37] |
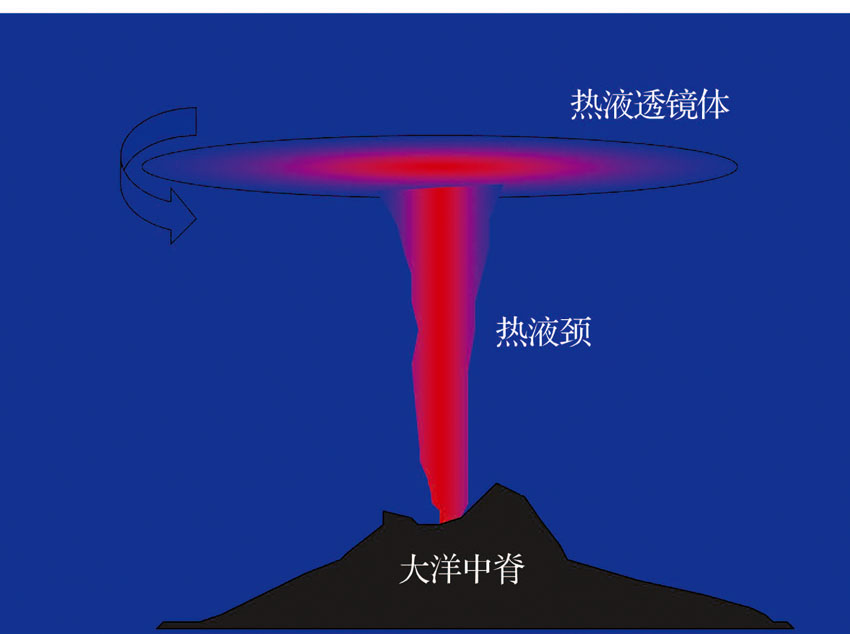
热液柱的形态是不断变化的。热液流体在上升过程中自身温度不断下降, 最终到达一个中性浮力面, 进而在这个面上逐渐聚集并向周围扩散, 从而形成一个透镜体。在柯氏力的作用下, 热液柱的透镜体发生旋转, 从而减少自身的能量向外扩散, 也保持了自身形态的稳定性。受深层海流影响的, 透镜体会发生横向运移, 扩散范围可从几十公里到几百公里, 甚至可引起深海大尺度环流(图4)[38]。
3.3.2 热液流体的热通量对海洋的影响
热液系统通过热液柱向海洋中输送巨大的热量, 影响海水的循环和气候的变化。不同的计算模型得到的结果差异较大。根据烟囱体计算的热通量为97.40 GW[40]; 根据扩散流和羽状流计算的热通量为84.90 GW; 根据热通量指数衰减法, 由洋壳传导通量估算的热通量为4.11 TW[41]。此外, 根据全球热液柱覆盖率推算的全球热液热通量为880 GW, 而根据岩浆热侵入的估计结果为445 GW[42]。尽管差别很大, 但这么大的热通量必然对海洋环流的变化造成巨大的影响, 需密切关切。
3.3.3 对周围生态系统的影响
热液活动是有生命限制的, 周期长短不一。如横跨大西洋地质综合调查(Trans-Atlantic Gaotraverse, TAG)区的热液活动延续了2万年, 而生命短的热液流体只有几十年或十几年[43], 消亡的热液区也存在重新活化的情况, 这些变化都会给生物群落带来极大的影响。对东太平洋海隆(East Pacific Rise, EPR)9° N不活动热液区的调查表明, 相对于活动热液区, 不活动热液区的细菌种群类型发生了明显变化, 其特点是热液区生活的细菌均与S, N, Fe及CH4循环密切相关[44]。此外, 热液的喷发不是连续的, 是脉动式、短周期的, 因此在喷发过程中会发出脉动的声音, 这可能是周围生物体活动或在不同热液区迁移的导航器。
1977年, 科学家乘“ 阿尔文” 号深潜器在东太平洋海隆约2 600 m深处发现了大量生物, 包括微生物、细菌[45]及无脊椎动物[32]等, 揭开了深海生物群落研究的序幕。20世纪80年代初期, 日本海洋科学技术中心(Japan Agency for Marine-Earth Science and Technology, JAMSTEC)也在冲绳海槽中部发现了热液堆积体和烟囱状热液喷口系统, 并在其周围观察到有巨大的海绵、虾类、海星等底栖生物。人们逐渐认知到地球上存在2种大洋:蓝色大洋(传统的大洋, 平均水深近4 km)和黑色大洋(洋底下的大洋, 流体活动可达海底以下约7 km), 并存在2种初级生产力及其食物链, 蓝色大洋初级生产力表现为浮游植物, 而黑色大洋的初级生产力则为热液细菌[46]。深海极端环境、热液系统以及极端环境下生物发展、演化, 引起了科学家的极大兴趣, 成为新兴的研究热点。
4.1.1 研究的意义
地质历史上, 海洋生物经过进化, 种群发生了巨大变化。而在现代的深海海底仍存在很多保持着远古特征的生物种类, 因此可以通过研究深海底的堆积物, 来了解地层的远古环境动态[47]。如在冲绳海槽中部伊平屋热液区发现的3种蔓足类生物, 其中有2种仍保持着原始体形。令人惊讶的是, 在冲绳海槽这一最年轻的弧后张裂体系中竟然保存着这种最古老形态的蔓足类[48]。
由于深海的极端环境和构造的变动, 在海底、板块交界处都分布着不同种类的生物群体, 但生物群落的迁移与板块构造历史密切相关[49]。因此, 研究这些生物群体有助于认识板块的运移和生物群体的进化, 填补生命发展史上的空白。
4.1.2 群落分布
一般来说, 不同的热液区表现为不同的生物群落[50]。据统计, 在热液区共发现了500多种真核细胞新物种, 包括12个门和150多个属。节肢动物(占38.8%)、软体动物(占28.6%)和环节蠕虫(占17.7%)是最主要的3种大型底栖生物, 刺胞动物(占4.6%)、脊索动物(占3.7%)和海绵类(占1.9%)则比较罕见[51]。并呈现以下特征:
(1) 不同热液区孕育着独特的生物群落
尽管深海洋流可以将有机体带到很远的其他热液喷口区, 但调查发现许多热液喷口均呈现出独特的生物多样性及某些特有种的富集[52]。例如, 加拉帕戈斯裂谷和东太平洋海隆具有相似的生物群落, 但与胡安· 德富卡海岭热液活动区的生物群落明显不同[48]。在东太平洋海隆热液活动区, 可见散布或呈“ 灌木” 状的多毛类生物聚集体(无消化系统的巨型管状蠕虫), 偶尔可见虾和雪人蟹; 在中大西洋海脊, 则多为成群结对的虾(Rimicaris)围绕在热液流体周围, 而很少见各种蠕虫[32]。同样, 印度洋Kairei和Edmond热液区也发育了独特的生物种群[53]。
在西太平洋冲绳海槽区, 伊平屋热液活动区共发现了1种热液伴溢蛤属、3种蔓足类、1种贻贝类、2种大肚须足虫和2种长尾类等。大肚须足虫、热液伴溢蛤等是优势种, 其湿重超过20~30 kg/m2[54]。其中, 蔓足类的同种属在东太平洋海隆并不多见。同时距伊平屋不远的伊士名热液喷口处却没有发现伊平屋热液区繁盛的蔓足类生物。如此近的距离上, 生物学上的种属有如此大的变化差异[48]。
(2) 不同的海底火山岩和沉积物有不同的生物种属
热液区不同的矿物、岩石或沉积物上附着的生物群落有很大的不同, 如在热液附近的一些浅色沉积物中分布着许多大型海参种的生物, 其体长可达30~40 cm; 而在枕状熔岩和带皱纹的板状岩石露头上(富含SiO2的喷出岩)则分布着棒球形玻璃海绵、海葵类等。但它们之间存在何种关系, 仍不得而知。或许尚需从矿物的成分、结构构造上去寻找答案。
(3) 分布的分带性
热液喷口的温度从中心向周围快速降低, 造成了不同温度带的生物群落以喷口为中心呈环带状分布。如靠近喷口处水温为60~110 ℃, 生存着大量的细菌和真菌, 一般都附着在沉积物和玄武岩表面; 再向外水温降到10~40 ℃, 则生活着各类蠕虫, 是与深海热液相关的无脊椎生物群落丰度最高的水温范围[32]; 在水温2~10 ℃, 生物门类大增, 以管状蠕状类为主, 但是也有不同生物门类混生的现象[55]。虽然热液区总的生物量很大, 但大型底栖生物很少。这可能与多变的流体动力、成分有关, 不符合大型底栖生物所需的特殊的生理生化习性[56], 从而其优势物种更为集中[57]。
(4) 生物的丰度和演替
一般深海生物群落的特点是种群密度和生物量都很低, 新陈代谢和生长速度都低于浅海。在热液喷口附近, 生物密度和生物总量却很高, 生长速度快, 但生命周期短, 平均生活年龄为6年, 平均生长速率为1~4 cm/a[58]。由于热液喷口生物群落直接依赖喷溢热液体获得能量维持生命, 因此海底热液的活动周期直接决定了生物体的产生和消亡。对于中— 快速扩张脊上频繁的火山活动区, 热液喷口生物群落常呈现月或年际变化, 更替频繁[59]。例如, 在胡安德富卡海岭上Cleft段火山喷发后2年内, 热液喷口出散布着管状蠕虫; 2~8年, 管状蠕虫则很少见, 生物群落由蠕虫类演替为贝帽类[25]。这表明, 流体温度较高的热液活动区以管状蠕虫为主, 而低温喷口区则以贻贝类为主(图5)。
 | 图5 “ 科学号” ROV在南海北部冷泉区拍摄的贻贝等生物群落及采集的甲烷气体(此图引自WPOS专项2014年第2期(总第3期)简报)Fig.5 The seep mussel pictured by ROV of vessel “ KE XUE” and methane gas collected in tube, northern South China Sea |
随着深海调查手段的发展, 深海微生物(细菌、古菌、真菌和病毒等)得到广泛而深入的研究, 越发凸显出其在深海极端环境扮演了重要角色[60]。深海微生物数量巨大, 占地球微生物总量的90%, 是主要的初级生产力之一[61]。在深海环境中, 病毒通过感染、裂解古生菌, 释放出大量的有机碳, 对深海微生物群落结构具有重要的调控作用, 对促进全球海洋碳循环也具有重要意义[62]。
为适应深海中存在的各种极端环境, 微生物形成了极为独特的生物结构、代谢机制, 已发现的极端生命形式包括嗜热菌、嗜冷菌、嗜碱菌、嗜酸菌、嗜盐菌、嗜压菌等, 被统称为极端微生物[63, 64]。它们独特的基因类型、生理机制及代谢产物, 为更好地认识生命现象、发展生物技术提供了宝贵的知识源泉。对此, 美国启动了专项“ Life in Extreme Environments(LEXEN)” , 集中围绕3个课题:①物种多样性; ②功能多样性; ③生命进化— — 生命如何从前生条件进化而来?并在极端微生物的生物技术利用方面取得进展, 如PCR酶、ENT酶、碱性酶及极端“ 石油工作者” 已在产业上产生了重要影响, 同时在基因芯片、新材料、新药等方面, 对极端微生物也抱有很大的期望。20世纪90年代初期, 日本执行了“ 深海之星(Deep-star)” 计划, 从深海中获得了1 000多株嗜压、嗜冷、嗜热、嗜碱及耐有机溶剂的多类型的极端微生物, 这些极端微生物在新酶、新药开发及环境整治等方面的应用潜力很大[65]。
4.2.1 微生物主要的分布区域
(1) 热液流体
热液本身含有的嗜热细菌随着其他热液物质一起喷出海底, 自生细菌和古菌呈“ 雪花状” 悬浮于热液柱水体中(图6), 并在热液喷口附着沉积下来。此外, 火山岩中也含有大量细菌。人们推测, 在深海热液活动区的海底表面下深部, 很有可能存在着一个极端嗜热的主要由古菌组成的无机自养微生物生态系统, 该系统可能与地球上的原始生命系统非常相似, 并有可能类似于地球外的生命形式[66]。
(2) 海底沉积物和海底以下地层
深海沉积物中生存着地球上数量最多的细菌、古菌、病毒等微生物[67]。在所谓的“ 洋底下的海洋” 里, 是深部生物圈中巨大的微生物群落聚居地。据统计, 地球上有高达2/3的微生物可能深埋在洋底的沉积物和地壳中。此外, IODP钻探揭示的海底深部生物圈的最大深度已延伸至海底以下800 m[66]。
4.2.2 极端环境的耐受性
目前已发现的极端生命形式包括嗜热菌、嗜冷菌、嗜碱菌、嗜酸菌、嗜盐菌、嗜压菌等, 分布于不同的极端环境中。当热液流体温度由150 ℃增长到300 ℃时, 嗜热古菌数量逐渐占据主导地位[68]。多种厌氧细菌则生活在海底深部[24]和烟囱体内[69], 其内酸度很高且温度变化大(25~350 ℃)。此外, 在深海热液喷口分离出的一种承压最大的耐压古菌(Pyrococcus yayanosii), 可生活在20~120 MPa。同时发现, 静水压力在原核生物在深海中的分布上扮演了重要角色[70]。因此, 对热液、冷泉区极端微生物进行遗传基因分析和进行组织学研究, 有助于认识深海极端环境下生物适应极端环境的生理机制。
 | 图6 冲绳海槽热液区喷口处“ 雪花状” 的细菌和古菌Fig.6 The snowflake shaped of bacteria and ancient bacteria above the hydrothermal vent in the OK |
4.2.3 化能合成作用
极端生命过程的研究包括一系列的异养和化能自养过程。然而, 对于深海海底这样一个似乎缺乏营养资源的环境下存在着巨量的生命体, 向生物地球化学、微生物生理学和微生物生态学等提出了新的课题[71]。
现代深海热液喷口调查发现, 热液中富含的多种矿物、还原性化合物(H2S, CH4)及CO2等为原料, 由化学能代替太阳能, 通过细菌和古菌的化能合成初级生产[17, 32]。同时, 由于热液系统中温度和化学组分的梯度变化, 为微生物体自由生长提供了多样的生理条件。热液区生物体可根据外界环境的变化, 在自养(非有机碳源)和异养(有机碳源)之间转换。因为在条件适合时, 异养可以产生更多的能量。然而深海热液区缺少有机质供应, 化能合成细菌可为热液生物群落提供有机碳, 也是物质循环中一个重要的汇[32]。由于热液喷口区H2S含量非常高(3~110 mmol/kg水), 硫化物氧化细菌是最主要的化能合成来源[17]。此外, 对热液喷口羽状流中的18种噬菌体基因组测序发现, 其中有15个病毒基因组含有与硫氧化酶相关的代谢基因, 这表明噬菌体在地球的硫循环中扮演着一种关键性角色, 且携带有热液系统中进行化学合成作用所需的遗传多样性[72]。
4.2.4 共生关系
现代海底热液喷口生态系统呈现出明显的大型生物与化学自养细菌、古菌等微生物共生特征。异养细菌与海洋病毒之间也存在着错综复杂的共生、寄生、基因转移等互作关系[73]。多种证据表明, 基本所有的热液区无脊椎动物都需要通过硫化物氧化细菌共生来获得碳和氮[17]。已知一种无消化系统的成年管状蠕虫, 获取养分的唯一途径即是通过体内一种称为营养体的组织, 该组织内即由硫化物氧化细菌组成[74], 其密度可达109~1011个细胞, 能使巨大的无消化系统的管状蠕虫(Riftia pachyptila)长到2 m[51]。对冲绳海槽伊平屋热液活动区热液伴溢蛤研究发现, 其鳃组织内充满了共生的细菌, 同时镜下可观察到大量的无机硫磺晶体[48]。日本深海微生物研究组发现一种类似酵母菌的单细胞真菌, 与管状蠕虫(Lamellibranchia)和白色巨蛤(Calyptogena)伴生, 以冷泉环境中高浓度的甲烷和硫化氢气体为原料[75]。附着在玄武岩表面、烟囱体上以及热液周围沉积物中微生物, 也可直接作为大量无脊椎动物的食物[76], 从而在新的热液喷口形成大型底栖动物定植的过程中扮演了重要角色。可见, 在深海底的生物群落是依靠化能合成为基础的生态系统。这种极端环境下生存的真核生物, 可能与细菌共生以摄取营养。
近年研究发现, 深海病毒在全球生物地球化学循环、深海代谢和生态系统演化中具有重要作用。病毒是深海数量最多的生物有机体, 病毒感染可占真核生物产量减少量的80%以上, 同时大量溶解的有机碳又作为其他细菌的营养来源, 从而维持了深海生态系统的平衡[62, 77]。
海洋及其海底沉积物孕育了地球上最大的生态系统之一, 其生存的细菌和古菌的数量占到全球生物圈原核生物总数的70%[78]。然而, 在深海极端环境的影响和制约下, 深海生物体分布是极不均匀的, 形成了独特的热液、冷泉等深海化能合成生态系统以及多种极端生命形式, 拓展了生命研究的深度和广度, 有助于揭示生命起源、生命极限、生命本质甚至其他生命形式等生命科学的悬念。
极端微生物作为一种特殊的生物资源, 蕴藏了巨大的经济利益。不仅可以提供新的遗传信息, 而且其独特的细胞结构、代谢机制以及各种多样的特殊功能, 可直接达到基因工程、蛋白质工程、代谢工程的某些目的, 产生新物质。如古菌拥有更大的染色体, 为天然药物开发提供了丰富的基因资源[79], 病毒被认为是地球上未知基因的最大贮藏库[62]。微生物代谢也成为人类疾病药物开发的主导[80]。但近50年来, 来自深海(水深> 200 m)的天然产品的数量只占到海洋天然产品的1%~2%[25]。
近年来, 我国在深海热液及其生态系统方面也取得了丰富成果, 通过ROV等先进深海探测技术, 在冲绳海槽热液区发现了新的蔓足类种属(Probathylepas faxian Ren & Sha, 2015)[81], 并获得了热液活动区高精度的地形数据。但由于厚层水体的覆盖, 深海微生物的调查、取样仍非常困难, 加之绝大多数的极端微生物不可培养, 利用微生物研究地质过程和环境也具有很大困难。因此, 应利用分子生物学原理, 以提取微生物DNA或利用地球化学方法提取有机物(如类脂物)研究。加强硫化物烟囱体本身的研究, 烟囱体具有“ 纵向分期, 横向分层” 的特征, 但关键是利用ROV等先进取样技术获得部位准确的样品。同时, 应加强学科交叉, 例如地质与生命科学、环境科学等学科的交叉, 从而更深入完整地研究深海热液生态系统及其对极端微生物的影响。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|