{kind=link}
{kind=link}
洞穴石笋δ13C在古气候重建研究中的现状与进展
[黄伟1, 2, 3  , 刘殿兵
, 刘殿兵1 , 王璐瑶1 , 张振球1 ]
, 刘殿兵|
|


作者简介:黄伟(1989-),男,江西瑞金人,博士研究生,主要从事洞穴古气候研究.E-mail:weihuang52@yeah.net
由于受气候、复杂岩溶过程等影响,石笋碳同位素(δ13C)研究相对于其他代用指标甚少,甚至被忽略。从影响石笋δ13C的气候与非气候因子入手,结合全球数十个洞穴记录及现代观测试验,分析碳同位素信号在岩溶系统内部的传递方式和路径,探讨不同时间尺度下δ13C与气候变化之间的关系。指出石笋δ13C虽然在百年—年代际尺度上表现出的噪音强度明显上升,但从轨道至千年尺度上看,温度、湿度(降水)等气候要素在不同区域所充当的主控因子角色直接或间接地影响上覆土壤植被状况、土壤CO2产率等当地生态环境变化。未来研究应深入挖掘δ13C信号在岩溶系统中的传输共性及其气候、环境控制因子,注重石笋δ13C季节变化特征研究,充分发挥其在揭示当地环境或灾害性事件方面的独特优势,并运用多指标、多种方法手段理解岩溶过程、全球碳循环与气候变化之间的联系。
First author:Huang Wei (1989-), male, Ruijin County, Jiangxi Province, Ph.D student. Research areas include paleo-hydroclimate based on the cave and global teleconnection.E-mail:weihuang52@yeah.net
The application of carbon isotope (δ13C) collected from stalagmite to reconstruct the past climate and/or ecologic evolution, relative to other preserved indicators, is much limited due to its complex influencing factors including climate outside cave and complicated and site-specific karstic process. In terms of various climate and non-climate limiting factors on stalagmite δ13C, and combined with a large number of geological records and modern cave monitoring data of the latest researches, the pathway and behavior of the signal transmission of δ13C in the cave system were analyzed. The possible relationship between δ13C excursions and climate oscillations under different time scales was thus discussed. Although the degree of noise becomes increasing mixed with δ13C on centennial scales or shorter, the climatic elements, such as temperature and humidity (or precipitation), as predominant modulators exert directly or indirectly influence on vegetation overlying the soil and associated soil CO2 productivity on millennial-orbital scales. Future work should be focused on further deeply extracting the common δ13C signals from specific caves by exploring the controlling factors, both including climatic and non-climatic ones, attaching importance to the seasonal characteristics of stalagmite δ13C, and taking full advantage of its potential in the indication of local environmental events. The use of multi-proxy and multi-method will contribute to better understanding the interesting linkages among the δ13C characteristics, karstic process, global carbon cycle and associated climate change.
近20年来, 石笋作为古气候重建的重要载体之一, 引起了国际国内学术界的广泛关注, 并在认识亚洲季风与全球气候联系方面取得了重要进展[1~11]。上述研究, 广泛采用氧同位素(δ 18O)重建和反演季风降水或季风强度平均状态演变历史[12]。而其中的碳同位素(δ 13C)指标由于受到洞穴复杂因素的影响未被充分利用。近年来, 随着现代观测及模拟实验对δ 13C信号传递过程及机制的深入解析[13~27], 石笋δ 13C在古气候/古生态重建、喀斯特岩溶系统与区域或全球碳循环等方面的优势和潜力也日益受到关注[14, 28~38](图1)。
由于石笋δ 13C信号特征本质上取决于土壤或大气CO2浓度的波动[13, 14, 85]; 因此, 从这个角度看, 石笋δ 13C所蕴藏的局域或区域气候信号在解释生态环境与气候演变关系方面具有重大潜力。Genty等[30]利用法国南部洞穴石笋δ 13C重建了5万年(80~30 ka BP)植被和土壤生物的演化过程, 间接重建了过去气候变化历史, 并与格陵兰冰芯记录的DO(Dansgaard-Oeschger)事件一一对应; 其δ 13C显示了较δ 18O更为敏感的波动幅度(表1)。
类似地, 中国东北暖和洞δ 13C同样记录了较石笋δ 18O更为显著的8.2 ka事件[53], 其可能的原因是中国北部更多地受到西风环流的控制, 土壤生物活性及植被变化对温度波动较为敏感。最近, Jo等[60, 61, 87]系统重建了朝鲜半岛多个洞穴石笋δ 13C时间序列, 显示出与以往已发表的东亚季风指标数据的变化具有显著的相似性。他们认为, 无论在轨道尺度还是在千年尺度, δ 13C波动反映了东亚夏季风强度(温度和湿度的综合效应)的变化, 并在此基础上提出了大气Seesaw的动力学机制[60, 61, 87]。在热带、亚热带较为理想的岩溶条件地区, 石笋δ 13C甚至能够指示百年— 年代际尺度的水文循环和气候波动历史[31, 32, 70, 77, 80], 通过与其他指标相结合, 寻找区分气候的自然变率和人为干扰的可能性[64, 70, 80]。这表明, 石笋δ 13C在解释植被变化、土壤生物产率和洞穴局部生态过程, 以及最终重建过去气候变化方面具有值得期待的潜力。
然而, 不同区域或洞穴石笋δ 13C记录显示, 其序列重现性较差, 无法在区域或更大空间范围内进行对比分析; 在气候信息的解译和提取方面也受到重大挑战, 其中最大的不确定性来自气候、局域生态环境以及洞穴岩溶过程的多重限制因子[14, 70, 88], 模糊了石笋δ 13C在指示气候意义上的解释, 从而使其在古气候/古生态重建工作中的广泛应用受到阻碍。为此, 本文回顾了洞穴石笋δ 13C在古气候/古生态重建研究的最新进展。在概述各种可能的限制因子基础上, 分析δ 13C在地表岩溶过程中的信号传输及继承机制; 探讨其在不同时空尺度上对气候、生态、洞穴岩溶环境和人类活动的响应方式, 以及可能的气候学/环境学解释。最后, 针对目前研究存在的一些问题做了初步探讨, 理解石笋δ 13C在指示气候演化历史过程中的优势及挑战, 以期在未来古气候/古生态重建研究中取得进一步突破。
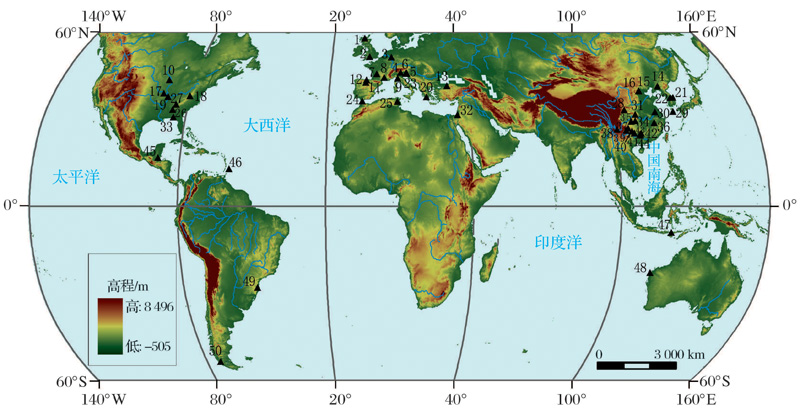 | 图1 部分洞穴石笋(或滴水)δ 13C记录分布图 1.Uamh an Tartair[39]; 2.Brown’ s Folly Mine[40]; 3.Bunker Cave[41, 42]; 4.Grotta di Ernesto[43]; 5.Postojnska jama[44]; 6.Grotta Savi[45]; 7.Villars Cave[15, 30, 46, 47]; 8.Chauvet Cave[46]; 9.Antro del Corchia[48, 49]; 10.Cold water Cave[28]; 11.La Garma Cave[50]; 12.西班牙北部洞穴群(Caves system in northern Spain)[51]; 13.Sofular Cave[52]; 14.暖和洞(Nuanhe Cave)[53, 54]; 15.石花洞(Shihua Cave)[55]; 16.苦栗树洞(Kulishu Cave)[56]; 17.Devil’ s Icebox Cave[57]; 18.Buckeye Creek Cave[34, 58]; 19.Crevice Cave[29]; 20.Kapsia Cave[59]; 21.Gwaneum Cave[60]; 22.Daeya Cave[61]; 23.Obir cave system[18]; 24.New St. Michaels Cave[19]; 25.La Mine Cave[46]; 26.Raccoon Mountain Cave[62]; 27.DeSoto Caverns[63]; 28.Dayu Cave[64]; 29.Yongcheon Cave[35]; 30.葫芦洞(Hulu Cave)[1, 65]; 31.黑龙洞(Heilong Cave)[66]; 32.Soreq Cave[67, 68]; 33.Hollow Ridge Cave[69]; 34.莲花洞(Lianhua Cave)[70]; 35.芙蓉洞(Furong Cave)[71]; 36.神农洞(Shennong Cave)[72]; 37.织金洞(Zhijin Cave)[73]; 38.Fulong Cave[74]; 39.竹蹓坪洞(Zhuliuping Cave)[37]; 40.七星洞(Qixing Cave)[22~24, 75]; 41.董哥洞(Dongge Cave)[36, 76]; 42.响水洞(Xiangshui Cave)[77]; 43.蟠龙洞(PanlongCave)[78, 79]; 44.Fengyu Cave[80]; 45.Actun Tunichil Muknal[31, 32]; 46.Harrison’ s Cave[16, 17]; 47.Liang Luar[81]; 48.Cave C126[82]; 49.Botuver’ Cave[33]; 50.Marcelo Aré valo Cave[83, 84]Fig.1 Distribution map showing locations for the published δ 13C records of cave |
| 表1 不同地区洞穴石笋δ 13C记录及其气候意义 Table 1 The δ 13C records from widely-spatial regions and its climatic interpretations |
洞穴滴水(或石笋)δ 13C信号响应于温度、降水等气候要素[28, 29, 67, 68], 这些气候要素主要通过调控洞穴上覆土壤生产率及局域生态环境而影响洞穴滴水溶解无机碳(Dissolved Inorganic Carbon, DIC)的浓度或δ 13C值。在亚热带地区, 通过定期洞穴观测研究发现, 岩溶滴水中的DIC含量与其δ 13C值均呈季节性波动, 两者表现为反相关关系[63, 78]。滴水δ 13C值在雨季较旱季偏负2‰ ~5‰ , 表明洞穴滴水所溶解的土壤CO2总量受外部气候要素(如降水、温度等)季节性变化的影响。这在一定程度上支持了石笋在重建古气候工作上的可靠性。
根据以往研究, 其主要的控制因素有:①植被类型的变化(C3/C4光合作用路径)[28, 29, 67, 68]。一般而言, 湿凉的气候条件更倾向于C3(木本植物及部分喜阴凉的草类)植被的生长, 植被的光合作用遵循Calvin循环路径, 其碳同位素值在-20‰ ~-34‰ 。相反, 干热的气候条件倾向于C4(禾本科植物为主)植被类型, 其光合作用的路径遵循Hatch-Stack, 同位素值为-9‰ ~-16‰ [89, 90]。因而, 当C3型植被占主导时, 其根呼吸和微生物活动所产生的土壤CO2同位素值相对小, 导致石笋δ 13C值总体偏负。当C4型植被占主导时, 土壤CO2的同位素值相对大, 导致石笋δ 13C值总体偏正[14]。②上覆土壤植被密度或生物量的变化。生物量与气候条件的变化有着密切关联, 影响土壤CO2产率及植物根的呼吸速率[75]。③土壤CO2产率和微生物活性的变化。当土壤湿度适宜, 温度升高有助于提高微生物活性, 增强分解土壤有机质的能力; 同时, 温暖湿润的气候条件有利于植被的生长, 从而增强根的呼吸, 提高了土壤CO2产率, 即提高了12CO2在碳同位素中的比例[14, 33]。④大气CO2浓度或源的变化。一方面, 大气CO2能够直接进入土壤带或岩体裂缝[13, 14, 18, 50, 91], 并与土壤CO2混合而最终影响次生碳酸盐的δ 13C值; 因此, 在更长的时间尺度上(如轨道尺度)大气CO2 δ 13C值的变化可能对石笋δ 13C信号调节有重要影响。另一方面, 大气CO2浓度变化对植物光合作用所引起的同位素分馏产生重要调控作用[85, 92], 即植物δ 13C值随着大气CO2浓度的升高而减小。然而, 刘贤赵等[93]通过回顾过去大量研究结果指出, 植物碳同位素分馏对大气 CO2 浓度的响应不是一种简单的线性关系, 大气 CO2 浓度对植物 δ 13C 的影响十分复杂; 甚至大气CO2浓度升高并非是导致植物δ 13C偏负的真正原因。
除了气候因子, 石笋δ 13C变化还受到从岩溶表层至洞穴内部复杂的地球化学过程以及水文过程的影响。①岩溶系统条件的变化(开放系统/封闭系统)。岩溶系统封闭(或开放)程度对基岩溶解量的控制起着重要的调节作用[50, 94]。Hendy[94]在早期的室内模拟实验中指出, 在封闭系统条件下, 母岩溶液的δ 13C主要受到CO2分压(pCO2)的影响, 两者呈反相关; 而在开放体系下, 由于无限制的气相CO2向水体溶液供给, DIC中的δ 13C变化主要受到温度的控制(0.6‰ /℃)。因此, 当岩溶系统处于开放状态, 从基岩中溶解的“ 重同位素” 在液相中通过持续地与土壤(或大气)CO2交换, 两相始终处于平衡状态[94], 从而导致母液中“ 老碳” 含量减少, 甚至完全被土壤(或大气)CO2所交换, 最终的碳酸盐沉积物δ 13C值偏小; 当岩溶系统处于封闭状态时, DIC与土壤(或大气)CO2之间的平衡关系不能保持, 来自基岩的无机碳存留于液相中直至沉积, 此时“ 重同位素” 含量较高, 相应的滴水或沉积物δ 13C值偏大[92, 94]。②碳酸盐沉积过程中的动力分馏。石笋沉积动力分馏与滴水速率、CO2的脱气速率、先期沉积以及蒸发有着密切关系[94~96]。当气候变干或特殊水文条件变化时, 水汽蒸发量增大, 且地表水源供给较少, 溶岩裂隙或流水通道处于少水或缺水状态; 滴水流速减慢, 渗流带溶液处于过饱和状态, 此时容易发生CO2的脱气溢出现象[16, 95], 从而造成在最终滴水沉积之前的碳酸盐沉积现象(称为“ 先期沉积” ), 即出现δ 13C值偏正[41]。另一方面, 当滴水速率过快或CO2脱气速率过快时, 溶液中的HC
此外, 洞穴内部空气湿度以及气流流动性决定了碳酸盐在二次沉积过程中的蒸发程度[21, 25, 27, 88, 99]; 甚至由于洞内外季节性温差和气压的变化导致洞穴内部pCO2也呈季节性变化, 使其能够独立于滴水速率变化而成为影响石笋δ 13C值的主因[18, 43, 100]。Baker等[39]在研究苏格兰河流洞穴滴水发现, 动力分馏导致CO2加速溢出, 滴水δ 13C偏正也高达5.5‰ ; 而奥地利Obir洞滴水表现出的动力分馏程度仅是前者的一半[18]。同样由于洞穴内外显著的温差和气压差导致的气流和CO2交换, 意大利Ernesto洞穴滴水沉积观测和模拟实验表明, 其造成了滴水δ 13C值从滴水到石笋顶部水膜的动力分馏达到了3.5‰ [43]。然而, 需要注意的是, 不同的分馏系数的引用对“ 气— 水— 岩” 分馏程度的判定存在一定误差[16, 41, 94, 101], 若实测值与理论平衡值差异不明显时, 多种分馏系数用于综合分析和判断则显得非常必要。由于自然洞穴沉积过程较室内模拟实验所受到的控制因素更为复杂, 前者的水— 岩分馏系数以及沉积物的同位素值远高于后者[41, 69], 突出了在自然条件下持续的洞穴观测研究的重要性和必要性。
由此可见, 石笋δ 13C具有复杂的响应性, 受到多种非气候因素的叠加影响, 至于何种因素占主导地位取决于具体的岩溶和气候条件[14, 92, 101], 但这些控制因子在很大程度上受到当地气候环境变化的驱动; 例如, 在干旱— 半干旱或蒸散作用强烈的气候背景下, 洞穴滴水CO2脱气速率及先期沉积的变化在指示气候异常方面具有重要的科学意义[51, 101, 102]。
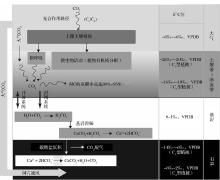
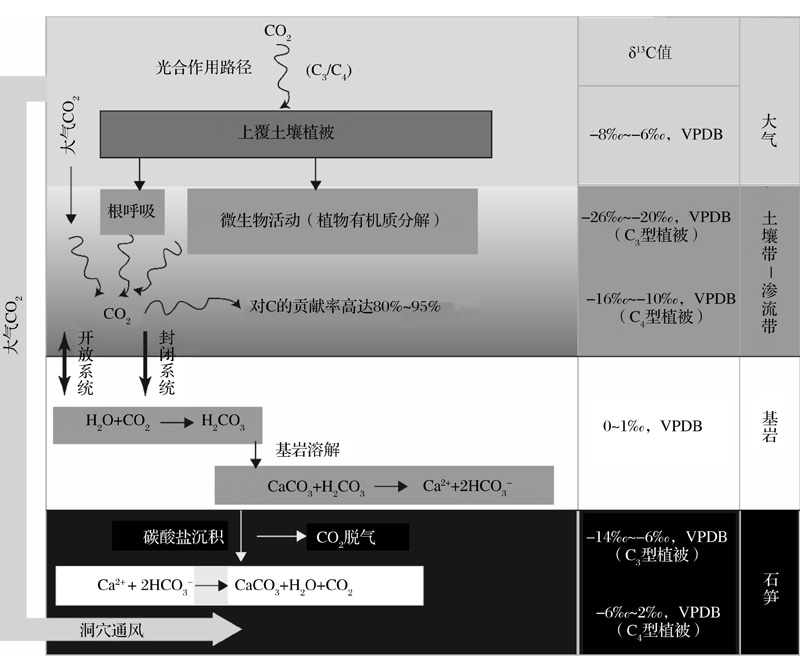 | 图2 洞穴石笋δ 13C信号来源及迁移示意图(根据参考文献[14, 50]修改)Fig.2 Schematic of the stalagmite δ 13C signal source and its transmission process (modified after references[14, 50]) |
正如前文(2.2)所述, 洞穴滴水(或石笋)δ 13C的信号主要来源于以下途径:①大气CO2; ②土壤CO2, 其中包括植物根的呼吸、有机质降解以及生物活性变化的产物; ③基岩溶解[13, 14, 91]。在岩溶包气带, 土壤CO2溶于水生成碳酸溶液。其通过裂隙通道与基岩作用而溶解碳酸钙, 直至母液达到饱和状态。当母液通过裂隙或滴水通道进入洞穴内部与空气接触时, 由于滴水与洞穴空气中的pCO2差异(通常情况下, 前者的pCO2较大), 滴水CO2脱气外溢而使溶液处于过饱和状态, 产生碳酸钙沉淀并沉积于石笋表层(图2)。由此可见, 从地表至洞穴内部, δ 13C信号在迁移过程中, 至少发生了2次同位素分馏过程:水
H2O+CO2+CaCO3↔ 2HC
以往研究表明, 石笋δ 13C信号主要受到土壤CO2的控制[29, 30, 33, 75, 103], 甚至洞穴空气pCO2的季节变化亦受到上覆土壤CO2产率的调节[44], 表明石笋δ 13C作为代用指标具有显著的气候意义。通过对中国南方洞穴的系统研究表明, 植被生物量的变化与石笋δ 13C有着密切甚至直接的联系[71, 75]。其机理可能是, 生物量所产生的植物根的呼吸以及有机质分解产生的CO2通量变化通过洞穴滴水传输至洞内并产生碳酸盐沉积, 从而实现δ 13C信号的迁移。然而, 由于影响因素的复杂性, 不同洞穴及区域所呈现的(洞穴滴水或沉积碳酸盐)δ 13C信号在相同时段具有显著差异[16, 17, 23, 50, 97, 98]。换言之, 若相同气候背景下的不同洞穴石笋δ 13C之间表现出个异性, 那么问题很可能来源于洞穴系统内部理想的δ 13C信号传输模式被多种局域地球化学过程导致的同位素不平衡分馏所干扰。
C
母液通过流水通道渗入洞穴内部, 并最终以水膜形式在石笋表层生成次生碳酸盐沉积, 即再一次发生水
当岩溶滴水滴入石笋表面时, 其水
尽管石笋δ 13C的气候意义存在争议, 但无论从理论的信号传输过程还是从地质记录看, 其在反映过去气候演变历史方面具有令人期待的潜力(表1)。但也不可避免地受到多重因素的限制, 在真实地记录古环境/古气候的过程中又不同程度地参杂了噪音成分。因此, 石笋δ 13C在多大尺度上能够有效地指示气候环境的变化, 或者不同时间尺度的δ 13C记录与气候大背景之间存在何种关联?哪些指标对于石笋δ 13C的准确解译具有更为可靠的参考意义?
在轨道尺度上, 冰期— 间冰期旋回产生了巨大的温湿差异, 赤道太平洋海表温差高达5 ℃[106]; 在其他地区, 如大陆或中高纬, 其产生的温差甚至更大, 极有可能造成植被类型的更替(C3/C4)[28, 29, 82]。由于光合作用路径的差异, C3型植被的δ 13C值(-20‰ ~-34‰ )较C4型植被(-9‰ ~-16‰ )低[82, 88]。在亚热带/温带季风区, 当气候处于温暖湿润的背景时, 洞穴上覆土壤植被类型多以乔木为主, 洞穴沉积物δ 13C变化主要继承了乔木光合及呼吸作用所转化而来的土壤CO2(-26‰ ~-20‰ )的信号, δ 13C值相对偏小; 反之, 当气候处于干热或干冷条件时, 植被类型可能转变为以禾本科类为主(尽管可能有部分C4型喜温凉的草存在), 其所产生的土壤δ 13
冰量在轨道— 千年尺度上的进退对中高纬地区植被类型的更替也产生了重要影响。美国密苏里州地区的植被类型由间冰阶(MIS3和MIS5)时的乔木转变为冰阶(MIS4)时的禾本科植物[29], 反映出植被生长环境的重大调整, 而温度变化似乎依然是植被类型更替的限制因子。北美东部的石笋δ 13C记录则表现出与全球冰量进退高度耦合性, 暗示两者之间存在紧密的气候联系[34]。然而, δ 13C与δ 18O在受到共同降水环境的调控下呈现出不同变化模式的机理仍然难以解释, 需要更多的记录加以支持与论证[34]。综上所述, 在轨道尺度, 无论是通过全球冰量变化或高低纬之间气压梯度变化, 石笋δ 13C所反映的洞穴上覆土壤植被变化至少受到区域或全球温度变化的影响; 特别是在冰期— 间冰期全球气候边界条件发生重大调整, 即发生了“ 源” 的变化, 巨大的温度变化很可能成为驱动大气环流调整、气候转型以及地表生态演变的主要触发器。
相对于冰期— 间冰期的气候调整而言, 千年尺度上的温度、降水等气候要素变幅较小, 发生植被类型更替事件的概率显著降低, 但对陆地生态系统的水文循环产生了不可忽视的影响。在降水季节性小或干旱半干旱地带, 温度对土壤CO2的影响似乎更为明显(表1), 与格陵兰冰芯记录、欧洲孢粉记录以及重建的大气温度记录具有良好的一致性[45~47, 81]。一般而言, 温度升高有利于增强土壤有机质分解速率及植物根的呼吸效率, 增大土壤pC
值得注意的是, 即使在相同的气候环境下, 局域的上覆土壤或植被条件影响着δ 13C与δ 18O的相位关系(正相关或负相关)。例如, 在MIS6/7界面以下层段, 南京葫芦洞石笋δ 13C突然增至-4‰ ~-6‰ , 研究人员认为这种同位素正向偏移现象反映了一次显著的地貌演化事件, 即洞穴上覆土壤被侵蚀殆尽, 洞穴滴水直接涌入岩溶系统内部而继承了大气CO2的δ 13C信号, 而并非指示植被类型的变化[107]。后期的研究进一步支持了洞穴水文效应导致δ 13C与δ 18O在DO旋回等千年尺度上呈负相关关系[65], 即由于上覆土壤较薄, 渗滤性较强, 大气降雨快速流过土壤带, 导致土壤粒间生物CO2与快速通过的雨水未达到同位素平衡状态(或称“ 过湿模式” )。这种现象同时得到以色列Soreq洞穴石笋记录的支持[68]。在不同气候带或气候调控因子条件下, δ 13C与δ 18O的相位关系并不一致; “ Hendy检验” 虽具有一定合理性, 但“ 同一纹层δ 13C与δ 18O具有相关性” 这条准则并不是检验其是否受到明显动力分馏的充分条件[16], 或者并不能否定将其用于古气候重建工作的意义[70]。在本质上, 石笋δ 13C及δ 18O的外部甚至洞穴内部一些影响因素均受到气候及生态环境演变的控制, 其变化方向具有一致性[88, 95]。
对于更短的时间尺度, 如百年— 年代际尺度, 石笋δ 13C由于在信号传递过程中与δ 18O的差异而表现出天然独特性。来自中美洲的现代石笋记录发现, 其δ 13C在年际尺度上与厄尔尼诺— 南方涛动指数(Southern Oscillation Index, SIO)强烈耦合, 表明植被生态系统对地球系统内部振荡响应的高度敏感性[31], 也为探讨过去更长尺度下是否存在厄尔尼诺现象提供了良好的材料和机会。同时, 石笋δ 13C在诊断δ 18O是表征长期的气候变化还是反映短时的强降水或飓风活动方面显示出独特的优势[32]。特别对于热带/亚热带飓风(或称“ 台风” )活跃地区, 其带来的短暂强降水能够对洞穴滴水δ 18O产生“ 掩盖效应” (Masking effect), 而对δ 13C则无明显影响, 从而使石笋δ 13C成为鲜明的“ 对照表” 。尤其是对于具有年纹层特征的石笋材料而言, 其δ 13C的季节或年际波动能够作为另一种时标校正工具, 从而弥补年代学所带来的较大误差的缺陷, 为研究重大气候转型突变特征提供全新的视野。
然而, δ 13C与δ 18O在百年— 年代际尺度上的复杂关系使学界对δ 13C是否具有广泛的气候意义产生质疑。一些研究显示, 在全新世时期, δ 13C与δ 18O变化在长期演化趋势上表现出一致性[45, 46, 90], 指示受到共同气候因子的驱动, 但仔细分析发现, 两者在若干百年— 10年际的波动变化上显示出明显的差异性; δ 13C的波动幅度甚至较δ 18O大(表1), 表明岩溶系统中的无机动力过程, 如岩溶系统的封闭— 开放程度、CO2脱气速率以及蒸发等因素对δ 13C施加了影响[14, 16, 95]。相反, 中国南方多支石笋记录发现, 虽然δ 13C在轨道— 千年尺度上表现出与δ 18O轨迹的显著差异性, 但在百年— 年代际尺度上两者显示出强烈的耦合性, 并在同一尺度上与格陵兰温度记录高度相关[36]; 欧洲及北美洲石笋δ 13C与δ 18O记录在反映全新世百年尺度事件上同样表现出趋同性[58, 108], 表明石笋δ 13C记录至少在响应显著的气候/环境事件方面具有广泛的气候意义。而季风环流所带来的降水及温度变化促使石笋δ 18O和δ 13C信号同向变化, 至少在季风区是可能的。显著的冷热、干湿波动引起的百年尺度的弱季风事件以及显著的强降水事件在我国西南季风区中晚全新世石笋δ 13C和δ 18O记录中得到证实[74]。
另一方面, δ 13C与δ 18O响应模式的差异同样存在于百年— 10年尺度的气候变化过程, 即δ 13C可能对温度的变化更为敏感, 而δ 18O则在很大程度上继承了大气降水同位素值的信号, 反映了在同一气候背景下两者对不同气候因素的选择性继承差异[35]。石笋δ 13C在百年尺度上的变化甚至认为是上覆土壤恢复/流失循环变化的过程[103], 其准周期(350~470年)独立于轨道岁差控制下区域大气降水δ 18O周期的变化, 在一定程度上反映了中国西南石漠化动力过程。至于其周期性石漠化的机制是否源于气候要素规律性的波动还有待于今后进一步研究。因此, 在短尺度上(百年— 10年际), 不同的局部生态环境以及洞穴系统内部受到动力分馏、地球化学过程等噪音影响的程度各异, 从而表现出δ 13C对真实气候信息响应的灵敏性差异。
除了自然过程导致石笋δ 13C波动外, 人类活动导致上覆土壤植被条件在短尺度上的变化也是一个不可忽视的因素。土壤侵蚀增加以及草本植物的扩张表明了人类活动可能对洞穴上覆土壤植被产生破坏而在洞穴石笋δ 13C记录中表现出来[109]。当遭遇气候变冷(如小冰期), 木材需求量的增长导致森林开发量大增以满足取暖和经济需求, 土壤层失去植被的缓冲保护, 从而导致δ 13C显著正偏[80]。历史上的朝代更替及政治战争引起的人口迁移及经济活动同样能够导致当地植被结构格局的改变, 而非气候因素造成[72, 73]。19世纪以来, 工业革命产生的大量12CO2排放至大气中, 土壤pCO2升高; 同时, 参观洞穴的游客人数不断增多, 导致洞穴内部pCO2也居高不下, 将不同程度地引起滴水CO2同位素值负偏, 最终造成石笋δ 13C相对偏负的现象[55, 64]。由此可见, 石笋δ 13C在百年— 10年际尺度上变化关系非常复杂, 既有与气候效应相一致的一面, 同时参杂了局域生态过程以及岩溶系统内部的水文动力、地球化学过程, 其变化甚至独立于外部气候驱动。尤其在中晚全新世时期以来, 自然生态环境同时受到气候、人类活动以及土壤、植被自身适应与演化的三重影响[59, 73, 74], 不同洞穴的δ 13C记录差异显著扩大。需要运用多指标, 如地球化学指标、石笋岩相变以及洞穴周边生态环境记录综合地解释δ 13C的气候意义[62]。
综上所述, 无论从现代洞穴观测/室内模拟实验还是从不同区域洞穴地质记录本身来看, 石笋δ 13C具备敏感地响应气候变化的潜力。从上覆土壤植被类型、密度、土壤pCO2到裂隙通道内δ 13CDIC信号迁移, 其均与气候变化引起的温度、降水等要素波动紧密相关[88]。然而, 石笋δ 13C具有自身的特殊性, 即其对当地生态环境的依赖性以及影响因素的复杂性。除了空气/土壤温湿度、上覆土壤发育、植被条件以及由此产生的土壤pCO2影响外, 石笋δ 13C还受到局部地形、地质条件、地球水文化学过程以及自身岩溶发育条件的限制, 各区域、各洞穴之间石笋δ 13C信息记录重现性较差, 其在用于重建古生态/古环境的过程中解译难度大, 受到重大挑战。因此, 今后的研究工作应集中在以下几个方面。
(1) 不同时空尺度的石笋δ 13C驱动因子研究。虽然对洞穴滴水δ 13CDIC信号传递和迁移过程中的影响因子研究有不少的文献报道, 但目前没有得到一个令人满意的结果, 对主导因子存在争议。传统观点认为, 洞穴上覆土壤CO2贡献了60%~95%的碳源[13, 14, 91], 从而成为洞穴石笋(或滴水)的主要信号来源。而最近部分学者指出, 大气CO2浓度或来源的变化以及包气带中下层植物深部根呼吸、有机质的分解成为主要的碳源[110]。尽管不能排除后者对δ 13C信号主控作用的可能性, 但这种现象可能具有特殊的时空特征, 是否能代表普遍的传递机制有待进一步研究和探讨。综合大量洞穴沉积记录, 石笋δ 13C在轨道尺度、千年尺度以及百年— 年代际尺度上对气候的响应方式差异明显, 反映出不同的主控因子变化。值得注意的是, 洞穴上覆土壤自身演化更替历史可能反映气候系统边界条件所引起的水热运移特征的变化, 也可能指示喀斯特地区独有的石漠化演化规律和历史[103], 两者均对洞穴沉积物δ 13C有重要的调控作用。另一方面, 不同气候要素(温度、降水、冰量)在不同地区对δ 13C的权重贡献率有所差异[29, 30, 33, 34, 46, 81], 应系统地研究各区域之间气候调控因子以及他们之间可能的数学转换关系。因此, 加强石笋δ 13C信号的归因研究有助于更好地解析不同区域、不同时间尺度主控因子的变化, 为建立石笋δ 13C与气候因素的函数关系奠定理论基础。
(2) 石笋δ 13C在季节尺度上的潜力挖掘问题。尽管目前有一些具有年纹层石笋记录报道, 但关注点仍然在δ 18O与气候要素的关系对比上[19, 111], δ 13C的价值仍然没有得到有效挖掘和利用。对于滴水通道顺畅且滴水滞留时间较短的岩溶条件而言, 洞穴滴水(或石笋)δ 13C信号特征能够更清晰地刻画气候要素在季节尺度上的轨迹, 为石笋年代学以及理解气候信号在洞穴系统中的传输过程及机制具有重要的科学意义。另一方面, 通过定期的现代洞穴滴水观测或现代方解石年纹层特征解析, 并与现代器测数据进行对比研究, 从而建立不同区域石笋δ 13C的季节变化特征与主控因子的准数量关系。从这个角度出发, 这种季节性特征联系可能延伸至千年尺度甚至轨道尺度, 探究主控因子分析的尺度转换问题[93], 从而实现“ 通过季节尺度来衡量千年或轨道尺度上δ 13C的气候指示意义” 。
(3) 利用石笋δ 13C加强局域生态环境研究。目前, 重建当地生态环境工作主要通过孢粉、泥炭或者洞穴有机沉积物, 而利用石笋中的无机碳进行生态重建的报道甚少。正如本文所呈现的那样, 无论地质记录还是现代滴水研究均表明, 其δ 13C的波动幅度较δ 18O大; 暗示了石笋δ 13C在响应气候或局域生态环境变化方面更为敏感, 如地球内部系统年际尺度波动(如厄尔尼诺事件)所引起的土壤水热条件、植被发育及生长条件等生态成分的特征变化[31]。另一方面, 随着人类活动印记的增强, 晚全新世气候变化显得更为复杂, 识别自然变率与人类活动的影响对准确地理解气候因子在全新世不同阶段的贡献权重具有重要意义。例如, 早期人类农业活动可能导致大量森林植被破坏或土地利用结构改变, 这些印记能够很好地被洞穴次生碳酸盐δ 13C所记录, 相反, δ 18O变化可能并不明显。因此, 石笋δ 13C在指示短尺度地球内部系统振荡以及揭示当地生态环境或灾害性事件[37, 72, 80]方面具有潜在的优势。
(4) 多指标、多方法研究。在更短的时间尺度上, 如百年— 年代际, 石笋δ 13C反映了复杂的混合信号, 不同洞穴系统所参杂的非气候信号比例出现不同程度地上升, 在对其气候意义的分析解译过程中则需要谨慎。在观测基础上通过模拟定量化计算出石笋“ 老碳” 含量[14, 92, 101], 从而科学地扣除δ 13C所包含的基岩干扰等非气候信息是今后广泛应用石笋δ 13C重建古气候/古环境信息的重要条件之一。微量元素在示踪岩溶系统水文地球化学特征方面具有独特优势, 例如, Mg/Ca能够很好地指示先期沉积引起的动力分馏[24, 92]; 87Sr/86Sr则反映了水— 岩接触时间[101], 即基岩对石笋δ 13C的贡献率。这些指标和方法能够有效地识别δ 13C信号传递过程中非气候因素成分。对于中晚全新世以来的δ 13C记录应该考虑人类活动对土地利用结构及植被覆盖条件的影响[112, 113], 运用史料及孢粉记录对其综合判断有利于解析人类活动与社会经济、政治发展以及气候之间的联系。
至今, 由于石笋δ 13C受控因素的复杂性以及不同岩溶洞穴的差异性, 其不能作为独立的代用指标建立古气候/古环境演变序列。在解释具体洞穴石笋δ 13C变化时, 应与δ 18O、地球化学指标、岩相、纹层学、器测历史记录以及生物学指标(如孢粉记录)等多指标相结合, 综合分析和判断, 科学地提取原始的区域气候信号。但石笋δ 13C记录至少有助于进一步解释δ 18O在区域或全球范围内的气候意义, 以及对理解温度与降水、当地生态环境与区域大气环流的联系具有重要意义。我们相信, 通过不断深入地研究碳(C)信号在岩溶系统中的传输共性以及对差异性因子的定量化, 石笋δ 13C在重建古气候/古环境演变以及理解全球碳循环与气候变化之间的联系方面将会发挥出重大潜力。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|