

{kind=link}
{kind=link}
{kind=link}
基于摇蚊的古环境和古气候国内外研究进展与展望
[胡玉 , 陈建徽
, 陈建徽* , 王海鹏, 吕飞亚, 魏国英]
, 陈建徽, 王海鹏, 吕飞亚, 魏国英]
|
|


作者简介:胡玉(1990-),女,河南信阳人,硕士研究生,主要从事摇蚊分析与古气候研究.E-mail:huy13@lzu.edu.cn
双翅目摇蚊科(Diptera:Chironomidae)昆虫生命周期相对较短,能够敏感响应环境因子变化,成为国际古湖沼学领域近20年来快速发展的代用指标,其主要近期进展可归结为以下4个方面:①样本训练集和转换函数的大量建立,以及区域数据库的对比和整合;②对湖泊水文、理化等内部因素在影响摇蚊群落方面所起的作用日益重视;③对现代间冰期以来、更为精细的时间尺度的关注;④实验技术和数值处理方法的改进,以及新指标的不断开发。国内的相关研究起步较晚,但迄今为止已经在长江中下游、青藏高原、云贵高原区、蒙新等地区开展了相应工作,并建立了摇蚊—环境数据库,同时关注湖泊内部因素对摇蚊种群的作用;古环境重建工作主要集中在西北干旱区和云贵高原区,东部地区的研究则主要反映了近期人类活动对湖泊状态的影响。就目前而言,基于摇蚊的古环境和古气候研究迫切需要:①开展更多的个体生态学工作,以广域范围内的现生调查为基础,构建属/种—环境因子之间的关系;②利用新技术手段进一步提高实验效率,提升鉴定分辨率;③注重先进的数量生态学方法以及同位素标记等新兴手段在研究中的应用;④充分重视摇蚊指标在湖泊自然状态界定和生态修复方面的作用。
Chironomidae (Diptera) becomes a rapid developing proxy in the international paleolimnology in the recent 20 years due to its short life cycle, strong ability to move and sensitive response to environmental change. The main progress of paleolimnological research based on chironomid can be summarized as the following four aspects: ①The establishment of a large number of sample training sets and transfer functions, as well as the comparison and integration of regional databases; ②More attention on the role of internal lacustrine factors in controlling of the chironomid population; ③Attention on finer time scales since the modern interglacial period; ④Improvement of the experimental technology and numerical methods, and constant development of new indicators. The domestic related research started late, but databases in the middle and lower reaches of the Yangtze river, the Tibetan Plateau, the Yunnan Plateau and the region of inner Mongolia and Xinjiang has been established so far, and also some researches on the internal lacustrine factors on chironomid population has been carried out; paleoenvironmental reconstruction is mainly concentrated in the northwest arid areas and the Yunnan Plateau, and the research in eastern region is primarily focused on reflecting the effects of recent state of human activities on the lakes. Paleoenvironment and paleoclimate researches based on chironomid urgently need to ①Carry out more individual ecology work to accurately understand the relationship between the species and environmental factors, combined with the investigation of a broader area; ②Further improve the efficiency of experiments by new technology to enhance the resolution of identification; ③Pay attention to the application of the advanced quantitative ecology methods and the novel tools such as isotopic analysis; ④Attach great importance to the role of chironomid in the definition of lake natural state and the ecological restoration.
高质量的古气候数据有助于增进对气候自然变率及其强迫机制的理解, 提高预测人类活动对气候变化影响的能力[1]。由于器测资料时长仅能覆盖地球历史的极小部分(< 10-7), 来自地质和相关载体的古气候代用指标就显得非常宝贵[2]。湖泊沉积记录因其连续性好、地理覆盖面广、指标体系完整等优势, 在重建陆地气候和环境变化方面的作用显得越来越重要, 是古气候、古环境研究的重要对象[3]。保存于湖相沉积中的摇蚊亚化石是新近发展起来的微体古生物代用指标[4], 利用摇蚊亚化石组合开展过去环境变化重建的重要性引起古气候学界广泛关注。
双翅目摇蚊科(Diptera: Chironomidae)昆虫是最为常见的淡水底栖动物类群之一, 其幼虫几丁质化头壳能保存于湖泊沉积物中, 易于提取和鉴定[5]。因其分布范围广、属种丰富(已记录的全球水生摇蚊共339属, 4 147种[6])、生态幅狭窄、生命周期较短、能敏感响应环境因子变化等优点而成为理想的古环境代用指标[7]。欧洲和北美的许多研究已证实摇蚊是古环境的有效生物指标, 在重建温度、湖水盐度、湖泊水深等方面相对准确可靠[7~9]。随着摇蚊— 温度(夏季表层水温、7月平均气温、年均温)[10, 11]、盐度[12, 13]、水深(或湖泊水位)[14, 15]、营养状况(由总磷、叶绿素a、总氮等表征)[16, 17]以及底层滞水含氧量状况[18, 19]推断模型在各地的建立, 基于摇蚊的古环境重建实现了定量化。相比之下, 国内的有关研究起步较晚, 我们曾以重建古温度为切入点, 对摇蚊指标在第四纪环境研究中的应用进行了综述[20]。随着其后基于摇蚊的古生态和古气候研究在国际上的不断深入和在国内的逐步展开, 有必要对该领域的最新进展作一总结、对值得进一步关注的问题进行展望, 以期对推进未来工作有所助益。
“ 转换函数法” 是目前基于微体古生物组合对环境因子开展定量重建最为成熟有效的方法之一[21]。其基本思路是利用分布于不同气候区域的现代样本训练集(training set), 采用多元统计方法, 探寻生物— 环境之间的关系, 并将这种关系运用到地层生物化石组合, 从而实现对过去环境状况的反演。实质体现了“ 将今论古” 前提下“ 以空间换时间” 的策略, 其中, 样本训练集是开展此类研究的基础。
目前, 本文共收集了全球范围内已建立的基于摇蚊的46个样本训练集相关数据(表1), 其分布、湖泊数量及目标环境因子如图1所示。仅就数量而论, 与Brooks等[60]综述时的18个样本训练集相比有显著增加, 在一定程度上反映了该领域良好的发展态势。具体表现出以下特点:
(1) 在地理分布上, 虽然仍以研究基础深厚的欧洲和北美更为集中, 但近年来的工作已经把基于摇蚊的样本训练集扩展到除南极洲之外的世界其他区域:Zhang等[17, 54]在长江中下游建立了亚洲第一个基于摇蚊的样本训练集, 提供了目标为总磷的转换函数; Verschuren等[47]在热带非洲的工作虽然以“ 出现与否(P/A)” 代替百分含量用以进行属种盐度最适值和耐受范围的估计, 仍可视作率先在非洲开展此类样本训练集研究的代表; Woodward等[59]在新西兰和Rees等[56]在澳大利亚的工作把基于摇蚊的样本训练集扩展到大洋洲, 用于夏季气温的重建; 最近Massaferro等[34]在阿根廷建立了南美洲目前唯一的基于摇蚊的样本训练集, 提供了重建区域年平均温度的有力工具。
(2) 从湖泊样本数量来看, 绝大多数(40/46)均不少于30个(表1)。其中, 湖泊数量在100个以上的大型数据库仍主要分布在欧洲和北美区域, 原因之一为下文将详述的“ 跨区域数据库整合” 。另一方面, 出现了在单个湖泊内部开展的摇蚊种群与环境因子(主要是深度)关系研究:例如, Kurek等[22]在阿拉斯加Zagoskin湖、Engels等[33]在美国东北部Crooked Pond建立了性能良好的转换函数并进行了水深变化重建, 与局地模型重建结果相比发现类比属种存在与否至关重要, 并强调对埋葬学过程的理解; Chen等[61]在中国最大的内陆淡水湖博斯腾湖, 利用均匀分布在不同水深梯度的表层样品, 发现深度是决定摇蚊种群最显著的环境因子, 初步的古水文重建显示与基于硅藻的结果具有良好的一致性。
(3) 在目标环境因子方面, 以温度为核心, 逐步拓展到其他环境因子。摇蚊作为古气候代用指标发展之初, 就是以重建温度而获得了一系列令人注目的成果[62, 63], 摇蚊也因此被认为是“ 在温带和极地环境重建温度的首选(生物)方法” [3]— — 归结于温度对摇蚊生理机能和生境的显著影响[37]。目前, 温度已成为第四纪摇蚊研究中最集中和最深入的领域, 现有的样本训练集多数仍以温度为转换函数的目标环境因子(29/46), 所获得的末次冰期以来温度重建结果被广泛用于区域对比和模拟研究。当然, 摇蚊— 温度关系远非一剂可以适用于所有环境状况的“ 万灵药(panacea)” [63], 对摇蚊古气候意义的阐释始终需要建立在现代过程研究的基础之上; 一项以重建温度为目标的多样点对比研究也显示, 其他环境因子(如水化学和生产力)的局地变化可能对单点摇蚊种群产生决定性影响[64]。反映在样本训练集上, 除温度之外, 盐度、营养状况、水深、底层滞水含氧量等湖泊内部环境变量在特定的气候区域均存在作为目标环境因子的实例(图1和表1)。例如, 在干旱/半干旱区的咸水湖和盐湖, 盐度对水生生物的胁迫作用使其(而非温度)成为控制摇蚊组合的最主要环境因子, 基于摇蚊的盐度转换函数在赤道非洲[13, 47]、我国青藏高原[53]、加拿大东南部[65]均有分布。
上述大量样本训练集的建立, 事实上为跨区域数据库的开发提供了基础。后者的兴起主要源自通过如下方法提高古环境重建质量的期望:在环境地质背景总体相似的情况下, 尽可能扩大目标环境因子梯度并保持湖泊在此梯度上的均匀分布, 从而使得对各摇蚊属种生态最适值和耐受范围的估计更为准确; 尽可能降低出现种群“ 无类比(non-analogue)” 状况的可能性, 从而使得前述空间到时间的转换更为可靠。
| 表1 图1中46个样本训练集的基本参数 Table 1 Basic parameter of the 46 chironomid training sets in figure 1 |
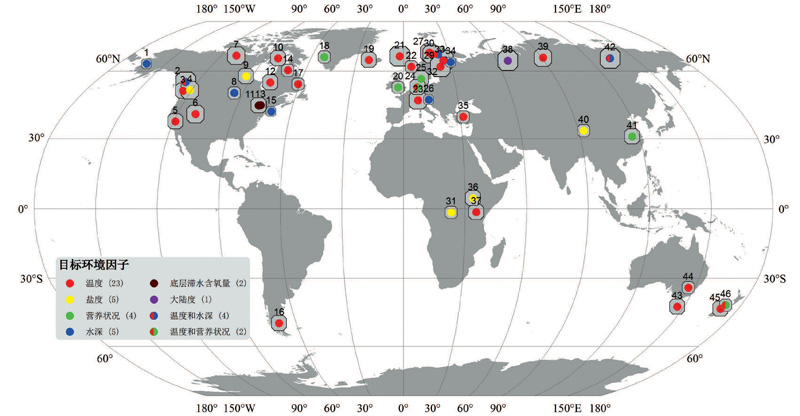 | 图1 世界范围内基于摇蚊的样本训练集分布、包涵湖泊数量和目标环境因子示意图(各样点具体情况见表1)Fig.1 Location of the published datasets documenting the distribution of chironomid training sets, the number of lake and the aiming environmental factor(detailed information of each dataset is in Table 1) |
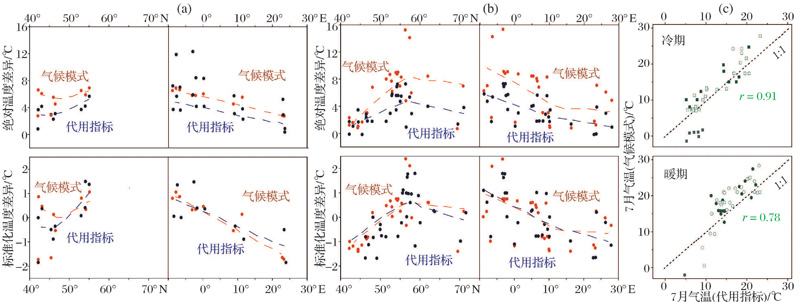 | 图2 晚冰期和早全新世摇蚊重建夏季温度与气候模式(ECHAM-4)输出结果对比(据参考文献[67]修改) (a)晚冰期早期/博令— 阿勒罗德间冰段过渡时期; (b)新仙女木期/早全新世过渡时期; (c)所有涉及的冷期和暖期Fig.2 Comparison of Chironomid-inferred July temperatures with the ECHAM-4 climate model runs(modified after reference[67]) (a) Early late-glacial/Bø lling-Allerø d Interstadial transition; (b) Younger Dryas/early Holocene transition; (c) All the investigated cold and warm periods |
此类研究始于20世纪末期, Lotter等[66]开展了欧洲和北美数据库的跨大洲合并, 并利用合并数据库和典型湖泊岩心样品重建了晚冰期温度变化, 虽然变幅和变率在不同模型间存在差别, 但总体表现出一致的变化趋势。Brooks等[43]以早先在挪威的数据库为基础合并了来自斯瓦尔巴群岛的样品, 有效扩大了温度梯度(尤其是低温端), 构建的西北欧摇蚊— 夏季气温转换函数性能获得显著提高, 迄今仍是同类研究中最为准确可靠的模型之一(表1)。Holmes等[36]将冰岛和挪威的校准数据集进行合并, 发现不同模型获得的重建结果趋势相似, 但基于合并数据库的重建获得的温度变幅更大。Heiri等[45]将瑞士、挪威和斯瓦尔巴的数据库合并, 构建了目前湖泊数量最大(274个)的摇蚊校准数据集, 涉及整个欧洲地区晚更新世沉积物中出现的大多数属种, 为大陆尺度的长序列温度重建提供了工具— — 最近, 利用这套合并数据库, 欧洲各区域晚冰期和早全新世夏季温度实现了统一方法和统一指标下的重建[67], 结果与ECHAM-4模拟数据总体上具有很好的一致性(图2), 为气候模式的改进和预测评估提供了重要参考。此外, Luoto等[48]合并了芬兰的2个数据库, 有效提高了变幅较小的全新世温度重建的精度。
上述研究展示了跨区域合并数据库在第四纪环境和气候研究中的巨大价值和潜力。同时需要指出, 合并本身也是引入异质性的过程, 过程中需要确保分类学和方法学的一致性, 因此分析质量控制至关重要。另外, 属种生态最适值的生物地理学差别、多重类比等问题也值得关注[36, 66]。
从以上全球各地理区域已建立的数据集可以看出, 虽然自20世纪90年代初至今建立了较多关于温度的样本训练集, 研究者却也逐步认识到盐度、深度、营养状况、pH值等湖泊内部环境因子对摇蚊种群的影响。由于摇蚊生命周期的大部分时间是以幼虫的形式生活在水中, 其所处栖息地内的各种环境因子理应决定着它们的生长发育过程, 因而湖泊内部因素在控制摇蚊种群方面所起作用日益得到重视, 表现为对特征属种生态意义日益深入的理解。
关于湖水盐度对摇蚊种群的影响:在湖水电导率3 000 μ S/cm以上的肯尼亚湖泊发现高耐盐种Tanytarsus minutipalpus, Microchironomus deribae和Kiefferulus disparilis的存在[13]。在加拿大西南盐度为10~400 g/L的湖泊中Cricotopus/Orthocladius和Tanypus的某些种广泛分布[12]。哈德逊湾低地摇蚊全部分类单元最适盐度从1.653(对应Paralimnophyes)到4.234(对应喜盐Orthocladiinae)μ S/cm不等[29]。对青藏高原地区42个湖泊组成的训练集进行分析, 发现高盐度湖泊如海丁诺尔(23 575.9 mg/L)、苏干湖(25 728.0 mg/L)和库赛湖(21 534.6 mg/L)的典型摇蚊种属组合为Orthocladius/Cricotopus和Psectrocladius barbimanus, 相对低盐湖泊如羊卓雍湖(1 025.0 mg/L)普莫雍错(452.6 mg/L)和空母错(311 mg/L)中的低盐属种Psectrocladius sp.3和Euryhapsis等占主导地位[53]。这些研究结果充分显示, 在干旱/半干旱区湖泊盐度变化对摇蚊种群的影响甚至比温度更重要[13, 20]。
湖泊水位变化常会导致近岸带和深水带的面积与比例(受湖盆形状影响)、沉积物的分布和沉积过程、水生植物数量和种类等方面的改变, 并对水体氧状况、水温、食物可供应量等因素产生影响[3]— — 水深对摇蚊种群的作用事实上具有综合性。Kurek等[22]在阿拉斯加湖泊研究中, 发现喜好浅水的Stictochironomus, Pentaneurini以及Tanytarsus与喜好深水的Micropsectra, Protanypus和Pseudosmittia/Smittia表现有清晰的湖内分布特征。在美国马萨诸塞州建立的本地校准集中, Ablabesmyia, Paratanytarsus penicillatus和Psectrocladius sordidellus等常出现在各湖泊的相对浅水区域, 而Chironomus anthracinus, Zalutschia zalutschicola和Cladopelma lateralis在超过5 m的深水区丰度更大[33, 68]; 此项研究还显示出提高Tanytarsus鉴定分辨率的必要性。我国新疆博斯腾湖摇蚊— 水深关系研究发现, 存在4个浅水种(Tanytarsus, Polypedilum nubifer, Cricotopus和Psectrocladius sordidellus)和3个深水种(Chironomus plumosus, Microchironomus和Cryptochironomus)[61, 69]; 整个种群变化还表现出8 m深度的阈值, 可能与水生植物分布与湖底地形有关[61]。
营养物质如氮、磷等的增加直接控制栖息地的营养水平和结构, 进而对摇蚊种群组成产生影响[70]; 此类研究一般用总氮(TN)、总磷(TP)、叶绿素a(Chl-a)等来表征湖泊的营养水平[10, 71], 探讨其与摇蚊种群的关系, 并将该关系用于湖泊分类、水质监控、界定生态恢复基线等工作中。芬兰的相关研究显示, Micropsectra radialis, Sergentia coracina, Heterotrissocladius maeaeri等表现为狭贫营养型, 而Chironomus plumosus, Glyptotendipes pallens, Paracladopelma等则喜好高营养水体; 属种多样性也与营养状况有明确的关系[71]。在西格陵兰南部地区营养丰富的湖泊中, 摇蚊种群主要由Ablabesmyia pulchripennis, Dicrotendipes modestus, Chironomus spp.等构成, 寒冷寡营养湖泊的Heterotrissocladius spp., Micropsectra insignilobus和Paracladopelma sp.处于优势地位[16]。在我国长江中下游地区, 总磷含量高的湖泊Microchironomus tabarui和Tanypus丰度相对大, 中等水平的湖泊Procladius以及M. tabarui之外的Microchironomus占主导地位, 总磷含量低的湖泊以Cricotopus sylvestris和Paratanytarsus占优势[54]。大洋洲新西兰样本训练集揭示的摇蚊— 营养状况关系与北半球同类工作有一定的相似性, 例如Chironomus的特定种可以耐受高营养水体, 而Naonella kimihia和Cricotopus aucklandensis更多出现于超微营养到中营养湖泊[59]。
事实上, 与营养状况有密切联系的湖水溶氧量本身是摇蚊种群结构[72]和丰度[73]的直接调节器。
摇蚊对缺氧环境的生理适应包括降低代谢率或转换成无氧代谢以降低对氧的需求[74], 以及淋巴生成更多血红蛋白以吸收结合更多的氧[75], 且大体积血红蛋白含量高的摇蚊能够更好地在相对深而缺氧的沉积物中存活[76]。安大略东南湖泊群研究显示出底层滞水含氧量对摇蚊种群的控制作用, Micropsectra等需要高溶氧才能生存, 而Chironomus和Dicrotendipes等可以耐受低溶氧状况[18]。其中, Chironomus plumosus等属种对低溶氧状况的耐受性在安大略中南部[19, 77]、芬兰南部[78]、法国东部[79]、格陵兰西部[80]以及我国东部[81]等区域均有报道。
得益于早期的经典工作— — 例如在北美地区确认Younger Dryas事件的存在[82]、又如对Laurentide冰原南部区域存在陡峭温度梯度的证实[83]、再如欧洲西北部与GRIP记录极为一致的晚冰期和早全新世温度重建[37]— — 目前, 对于摇蚊可准确记录冰期— 间冰期尺度上或冰期内部(如冰消期)的大幅气候(主要是温度)波动已基本成为共识。
从21世纪初开始, 逐渐有研究开始关注变幅相对小的全新世气候变化。例如, 在澳大利亚东北部Dimitriadis和Cranston利用共同气候范围法定量重建了区域全新世温度变化[84], 与孢粉分析的结果基本一致; 欧洲区域相继在芬兰[46, 48]、瑞士[38, 85, 86]、英国[87, 88]、瑞典[89]、冰岛[90, 91]、奥地利[92]、法国[93]、波兰[94]等地利用摇蚊指标全新世气候进行重建; 在魁北克南部重建了全新世8月气温, 并通过与硅藻、孢粉指标的对比, 证明了摇蚊气温重建的有效性[95]。Velle等[64, 96]通过在挪威和瑞典的工作提出, 鉴于区域内不同湖泊的摇蚊重建温度结果表现出不一致, 摇蚊不宜用作有效的全新世温度代用指标, 其他环境因素(如营养状况、水深、pH值)的作用可能更为重要。Brooks等[97]对此进行了逐条评论, 认为尽管Velle等提出的改进摇蚊— 温度重建的多数建议值得采纳(实际上在实践中已经注意到, 例如随着湖泊变浅存在高估重建温度的可能性[46], 其他环境因素也确实可能对温度重建造成影响, 但这种影响是可以通过摇蚊属种以及多指标研究来辨识的(如Berntsson等[89]对温度重建序列中降水信号的辨识)), 在以温度为目标环境因子所构建转换函数表现出高性能的区域, 温度仍然是摇蚊分布和丰度的最主要控制因素。
20多年的研究工作证实了Brooks等[97]的观点:①基于摇蚊的冰消期和全新世温度重建得到了其他独立代用记录(包括冰芯氧同位素、孢粉等指标)的支持[37, 99~102]; ②摇蚊温度重建能够捕捉到全新世的一系列短尺度事件, 例如11.3, 10.3, 9.3, 8.2, 5.0 ka以及最近的一次冷期小冰期[85, 90, 92, 103~106], 可见其对幅度相对较小的温度变化也足够敏感; ③更为精细的记录表明, 在过去100年间, 甚至可以实现摇蚊重建温度与器测记录的校准[107~110]。
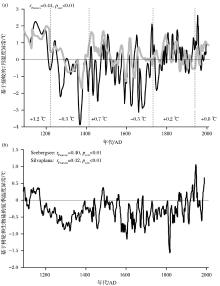
事实上, 为提高对当今气候问题的理解及对未来气候走势进行预测, 用摇蚊指标来重建千年甚至百年尺度气候状况的研究成为近几年本领域的热点之一。例如, Porinchu等[111]重建了美国中央大盆地近百年来的温度状况, 发现7月平均气温在20世纪早中期经历了大的波动, 在20世纪晚期持续高于平均值且与观测数据表现出很强的一致性; Medeiros等[30]在加拿大Nunavut地区的Baker湖重建了100多年来的湖泊表层水温波动, 发现在过去60年Baker湖的盛夏表层水温上升了2 ℃, 这一结果也得到了临近气象站点实测记录的验证; Bunbury等[112]在加拿大Yukon地区西南湖区重建了近2 000年的7月平均气温, 发现其温度变化不仅具有明显的阶段性, 而且各阶段的温度变率也有差别。欧洲相关研究的发展则更为迅速, 如冰岛1650 AD至今[110]及过去千年[113]、挪威过去1 760年[114]、芬兰近700年[106]、法国近2 000年[93]、瑞士近千年[85]和近百年来[115]、奥地利过去300年[116]、俄罗斯近150年来[52]的温度重建。尤其值得一提的是, Larocque-Tobler等[117]在瑞士东西侧2个湖泊做出了迄今为止年代控制最好(纹泥)、分辨率最高(1~20年)的过去千年基于摇蚊的温度重建(图3), 显示了清晰的中世纪暖期和小冰期, 以及更短尺度的气候波动; 2个湖泊的温度记录之间、它们与区域乃至半球尺度温度变化都具有很好的一致性, 并进而得出近期的增温仍在中世纪暖期增温范围内的认识。
当然, 对短尺度环境变化的关注不仅限于温度, 利用摇蚊种群变化来反映近千年、甚至近百年营养状况[118~120]、水深[15, 69, 79]、盐度[121, 122]、底层滞水含氧量[78, 123, 124]的研究也日益增多, 并取得了一系列有意义的结果。
2.4.1 实验技术和数值处理方法的改进
自20世纪90年代至今, 各国学者对摇蚊化石的分析方法进行了一系列的探究, 形成了样品分散— 分级过筛— 头壳拣选— 上片鉴定的基本流程[60], 是目前大多数第四纪摇蚊研究采用的一套标准方法。在实际操作中, 针对不同性质的样品, 存在一些细节上的调整— — 例如对于碳酸盐含量高的样品, 通过超声波水浴更利于分散[125], 或者在完成第一遍拣选之后加入稀盐酸进行二次拣选以确保头壳提取的完整性[121]。然而, 这套标准方法毕竟耗时长、效率低(即便是熟练的实验人员, 完成一个样品的制备和鉴定也需要1~1.5个完整工作日), 特别是难以对头壳浓度较低的样品开展高分辨率分析。因此, 研究者从未停止过对高效提取沉积物摇蚊亚化石方法的探索, 并取得了一些进展。
Rolland等[126]尝试用煤油浮选来提取沉积物中的摇蚊头壳, 获得了较好的结果, 也逐渐用于实际研究[127, 128], 但这种方法对于较大个体(头壳里填满沉积物)的分离效果仍有待改进。Larocque-Tobler等[129]利用棉蓝染色的方法来处理样品, 结果显示能提高摇蚊头壳拣选和鉴定的效率。Velle等[130]通过在样品中加入外来标记物来估算头壳浓度, 发现结果与分析全样并无显著区别。Verschuren等[131]在非洲湖泊的实践表明, 可以通过增大网筛筛孔的方式来减少摇蚊样品制备时间, 同样能够获得可靠重建。然而, 在高纬较寒冷的湖泊中, 由于个体较小的摇蚊所占比例很高, 采用大筛孔方法无法代表种群的真实组成, 所获重建结果也出现较大偏差[132]。
在数值处理方法上尚未有太大突破, 值得注意的是对转换函数和对重建结果的检验:
(1)传统方法对转换函数评价的标准来自对同一空间数据库的“ 交叉检验” (实际并非基于独立数据的评价结果), 近年来在瑞典[107]、瑞士[108, 109]、冰岛[110]等地相继开展了一些利用器测资料校准来评价转换函数性能的工作, 实现了基于独立数据的评估; 遗憾的是, 受材料(高沉积速率和可靠年代学)所限, 此类研究难以大范围推广。
(2)更具一般性的是Telford等[133]提出的利用随机数据检验环境因子重建结果的新方法, 其优势在于考虑了转换函数和钻孔数据的结合。也就是说, 利用钻孔数据和转换函数重建目标环境因子和随机因子, 将2类重建结果对化石数据变化的解释量进行对比, 从而确定目标环境因子重建的显著性— — 这种方法实际考虑了构建转换函数的最终目的, 即实现古环境重建; 若最终所获重建不具备统计上的显著性, 则无论多么优秀的“ 纸面” 性能都毫无意义。该方法已在欧洲和北美得到愈来愈多的应用[48, 134, 135]。此外, 也有学者开展了利用贝叶斯模型[46]、人工神经网络[136]等手段来构建转换函数的尝试; Kohonen人工神经网络(自组织映射SOM)还被用于层序限制的聚类分析, 被认为是传统CONISS方法的有效补充[137]。
2.4.2 新指标的开发
传统上, 一般用摇蚊属种组合变化来反映古环境变化; 近年来, 稳定同位素分析方法正被应用到基于摇蚊的古环境研究中。
一方面, 由于摇蚊幼虫从它所在水环境获取化学元素进行生物合成[138], 使摇蚊亚化石头壳稳定同位素(δ 18O, δ D, δ 15N和δ 13C)能反映其水生环境的同位素变化。另一方面, 稳定同位素分析技术和仪器的进步, 极大地降低了对分析所需样品量的要求[139], 使沉积物中摇蚊幼虫亚化石同位素分析成为可能。Wooller等[140]率先采用摇蚊稳定同位素分析方法, 研究了美国、加拿大和格陵兰岛4个湖泊沉积物中摇蚊亚化石δ 18O与降水δ 18O之间的关系, 进行了晚第四纪温度定量重建的尝试, 显示了这种新方法在古环境定量研究方面的极大潜力。随后, Wooller等[141]又测定了冰岛东北部湖泊无脊椎动物残体及有机质的多种稳定同位素, 发现δ 18O与δ D, δ 15N, δ 13C变化不一致, 推断摇蚊头壳δ 18O的变化并不是由食物所决定, 可能是大幅度的气候(温度、降水来源、降水季节性)变化的结果。之后, Verbruggen等[142]测定了瑞士Rotsee湖晚冰期沉积物中摇蚊幼虫δ 18O, 其变化与碳酸盐记录的湖水δ 18O趋势一致, 以此证明摇蚊δ 18O可以准确记录湖水δ 18O的过去变化, 为缺乏碳酸盐的湖泊δ 18O测定提供了一种可行的方法。
至于摇蚊残体的δ 13C和δ 15N, 通常认为其反映摇蚊幼虫食物的同位素结构, 受栖息环境营养水平的制约, 可将其作为摄食生态学研究的代用指标[143]。对加拿大2个受海鸟影响较大的高纬湖泊沉积物中摇蚊残体进行了δ 15N分析, 发现摇蚊残体δ 15N值和有机物中δ 15N变化一致[144]; 对摇蚊残体δ 13C与湖泊甲烷通量之间关系的分析研究表明, 摇蚊残体δ 13C值可作为湖泊过去甲烷变化的证据, 展示了利用摇蚊开展环境变化下甲烷动力学研究的潜力[145~147]; 根据摇蚊头壳和枝角目残体的δ 13C变化趋势, 推断了湖泊近150年碳循环的变化[148]。
由于摇蚊头壳稳定同位素与栖息地(如沉积物、湖泊水体和水生植物)同位素之间关系复杂, 尚存在较多不确定性。为了更好地揭示湖水和摇蚊幼虫所摄取食物同位素组成与摇蚊组织和化石结构同位素组成之间的关系, 还需要开展系统的摇蚊幼虫培养实验[149]。此外, 提高对摇蚊幼虫稳定同位素组成的季节变化和季节性是如何影响化石样品稳定同位素值的理解也至关重要。
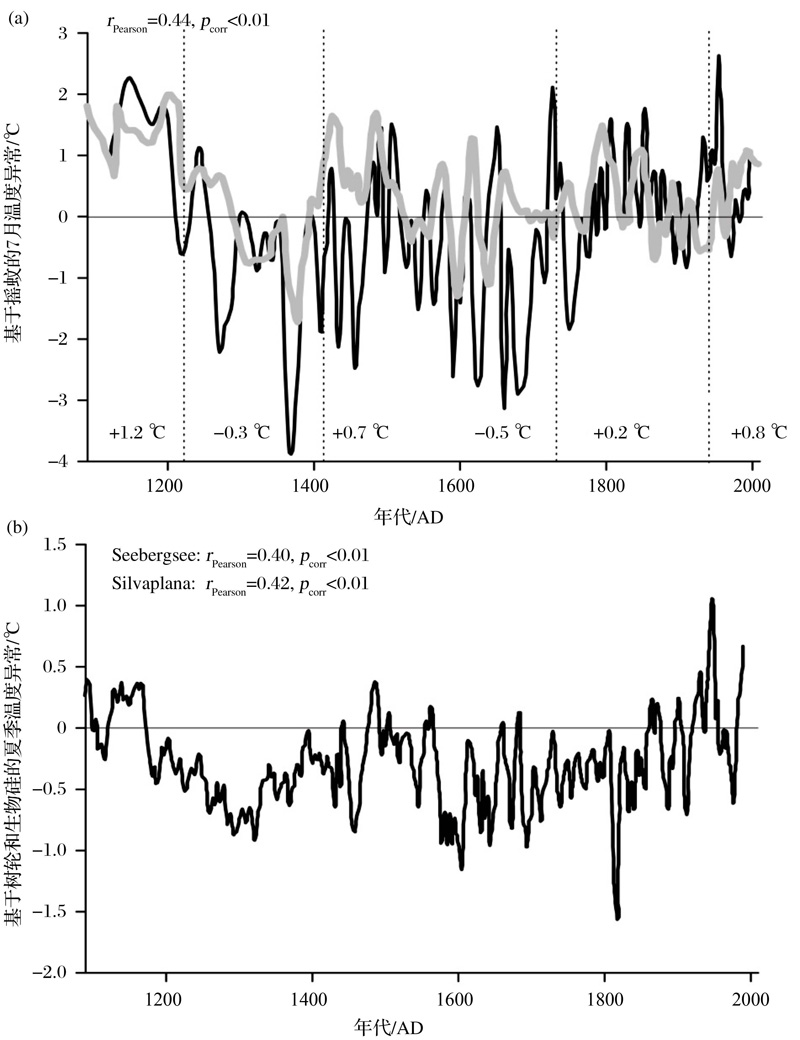 | 图3 近千年来摇蚊重建瑞士温度记录及其与邻近区域树轮和生物硅综合重建温度记录的对比(据参考文献[117]修改) (a)瑞士东部(黑线)和西部(灰线)湖泊基于摇蚊的过去千年7月温度重建; (b)瑞士东部树轮和生物硅综合夏季温度重建Fig.3 Comparison between the chironomid-inferred temperature records and the tree-ring and biogenic silica-based temperature record during the last millennium in Switzerland (modified after reference[117]) (a)Chironomid-inferred mean July temperature anomalies from western (gray line) and eastern Switzerland (black line); (b) A composite ofJune-July-August temperature reconstruction based on six tree-ring records and a biogenic silica record from eastern Switzerland |
综上所述, 随着多指标样本训练集的大量建立和跨区域数据库的整合及其在全球范围内的广泛应用、对湖泊内部因素在控制摇蚊种群方面所起作用的日益重视、对全新世以来更为精细时间尺度气候变化的关注、以及实验技术和数值处理方法的改进和新指标的不断开发, 基于摇蚊的古环境和古气候研究已取得了很大进展, 摇蚊也成为国际第四纪研究的重要生物代用指标, 在增进对地球环境系统近期变化的理解方面发挥着愈加重要的作用。在未来的研究中, 需要更加重视以下几个方面:
(1) 加强个体生态学研究。摇蚊属种— 环境因子之间的关系是基于摇蚊的古环境重建研究的基础, 加强对个体生态学的理解, 一则便于在研究基础相对薄弱的区域更为可靠地应用指示种方法, 二则有利于提高转换函数的效能, 特别是在样本训练集涵盖区域范围较大时, 同一属种的生态最适值可能存在生物地理学上的差异时更是如此。
(2) 提高实验效率, 提升鉴定分辨率。实验效率的提高使得高分辨率的快速分析成为可能, 这就要求利用新的技术手段不断进行探索。就鉴定分辨率而言, 同属中, 种间最适栖境千差万别, 无疑是提高鉴定分辨率的最重要理由; 但同时需要指出, 鉴定分辨率的进一步提高通常会降低实验效率。特别在定量重建中, 不影响模型性能的前提下, 最优方法可能来自鉴定分辨率与实验效率的权衡结果。
(3) 注重先进的数量生态学技术及新指标和新方向在研究中的应用。积极吸收和借鉴数量生态学研究的最新成果开展工作, 例如共同对应分析(Co-CA)、分类和回归树(CART)、主曲线(PC)、普氏分析与校准分类对比、前文已提及的对于环境重建的显著性检验等手段; 同时, 重视新指标(例如幽蚊/摇蚊比、同位素标记)和新方向(例如放射性碳测年)在研究中的应用, 为诸如湖泊沉积碳循环等学术界的热点问题做出贡献。
(4) 充分重视摇蚊指标在湖泊自然状态界定和生态修复方面的作用。结合其他指标和社会经济数据, 区分人类活动对湖泊生态系统的影响, 特别是界定湖泊在未受人类活动扰动之前的状态, 为湖泊的生态修复目标设置提供依据— — 这也正是古湖沼学研究服务于社会需求的重要体现。
最后应该指出, 中国的摇蚊— 古环境研究虽起步较晚, 但迄今为止已经在长江中下游[17]、青藏高原[53]、云贵高原区[150]和蒙新地区(张恩楼, 个人通讯)建立了数据库, 也开展了湖泊内部因素对摇蚊群落的影响研究[53, 54, 61, 81]。西北干旱区[121]和云贵高原区(张恩楼, 个人通讯)的研究以古环境重建为主; 东部地区的研究则主要反映了近期人类活动对湖泊状态的影响[118, 151]。有关国内此领域工作的具体进展, 可参考近期发表的综述论文[152]。基于国内的研究现状, 除上述4个方面亟需加强之外, 还应:①重视摇蚊指标在古温度重建上的作用; ②推进典型气候区基于摇蚊的湖泊深钻研究; ③结合我国现生摇蚊分类学研究成果, 建立健全的摇蚊亚化石分类体系。充分利用我国高分辨率湖泊沉积和丰富多样的摇蚊属种, 开展多指标综合应用, 以期实现对过去环境和气候变化图景更全面的理解。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|
| [130] |
|
| [131] |
|
| [132] |
|
| [133] |
|
| [134] |
|
| [135] |
|
| [136] |
|
| [137] |
|
| [138] |
|
| [139] |
|
| [140] |
|
| [141] |
|
| [142] |
|
| [143] |
|
| [144] |
|
| [145] |
|
| [146] |
|
| [147] |
|
| [148] |
|
| [149] |
|
| [150] |
|
| [151] |
|
| [152] |
|