




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
豫西荥阳陆相二叠纪—三叠纪之交的微生物成因构造(MISS)及其地质意义
[郑伟1  , 齐永安
, 齐永安1*, * , 张忠慧2 , 邢智峰1 ]
, 齐永安, 张忠慧|
|


作者简介:郑伟(1979-),男,河南商丘人,副教授,主要从事地球生物学和沉积学研究 E-mail:zhengw99@hpu.edu.cn
微生物成因构造(MISS)是微生物活动及其生物地质作用的沉积记录,为人们认识早期地球生物以及微生物活动提供了依据。微生物成因构造在前寒武纪以及显生宙地质危机期海相地层中分布较广,而在陆相地层中则报道较少。在豫西荥阳地区二叠纪—三叠纪之交的陆相地层孙家沟组和刘家沟组中,发现了较为丰富的三大类微生物成因构造(生长构造、破坏构造和腐烂构造),可详分为瘤状突起、变余波痕、隆脊构造、似正弦状脱水裂痕、纺锤状脱水裂痕、树枝状脱水裂痕、多边形脱水裂痕、曲形脱水裂痕、砂火山构造、微生物席砂片和似雨痕构造11个小类。微观分析显示:脊处纵切面显示U型、石英颗粒和黏土矿物条带定向排列、捕捉的云母颗粒不连续和近平行等特征,这些均表明其为微生物成因。研究区发育丰富的微生物成因构造,是二叠纪末灾变事件导致研究区陆地生态系统迅速退化,微生物大量繁殖而形成的,代表着以微生物群落为基础、后生生物少、钙质结核等特殊沉积发育的灾后陆地生态系统。通过对陆相微生物成因构造的研究,有助于人们全面了解二叠纪—三叠纪之交全球灾变事件在陆相环境中的具体表现,为认识微生物席在全球的空间分布提供新资料。
Microbially Induced Sedimentary Structures (MISS) are primary sedimentary structures that arise syndepositionally from microbial community activity. Especially valuable are MISS for the analysis of early Archean (and extraterrestrial) deposits. However, most reports of MISS have focused on the Precambrian and Phanerozoic mass extinction marine sediments, and only a few and terrestrial MISS have been studied. The MISS presented in this paper, which mainly consists of mat growth feature, mat destruction feature and mat decay feature, are well preserved in terrestrial clastic rocks around the P-T boundary in Xiyang area, western Henan. Large U ridges, oriented sand quartz, mica grains and thin clayey laminae arranged parallel to bedding plane are the main features visible under the microscope. Several lines of evidence indicate that the Xingyang MISS are of biogenic origin. Abundant MISS in Xingyang may indicate the degradation of terrestrial ecosystems and proliferation of microbial mats immediately after the severe Permian-Triassic mass extinction. Study aiming at MISS helps to analyze their space distribution in the globe and to probe into links between microbial proliferation and environmental stresses following the end-Permian mass extinction in terrestrial ecosystems.
微生物席是微生物群落在沉积物与水体界面处, 与沉积物相互作用、相互影响形成的一种特殊的生物— 沉积构造[1, 2], 是微生物群落活动的重要产物。Schieber[3]、Gerdes等[4] 、Noffke[5]和史晓颖等[6]把微生物活动与沉积环境交互作用及微生物新陈代谢、生长、破坏和腐烂等在沉积表明形成的各种构造, 称作微生物成因的沉积构造(Microbially Induced Sedimentary Structures, MISS)。它们是认知地球早期生物、硅质碎屑沉积环境下的微生物生命活动及相关地质作用的重要记录[6]。近年来, 一些地质学者在中— 新元古代以及显生宙特殊时期的海相碎屑沉积环境中, 发现、报道、论述了相关微生物成因构造[1~22], 这些研究集中于前寒武纪和显生宙特殊时期海相地层中, 而在显生宙特殊时期的陆相地层中少有报道。作者在豫西荥阳地区二叠纪— 三叠纪之交的陆相地层孙家沟组和刘家沟组中, 发现了大量微生物成因构造, 它们在研究区普遍发育, 较容易识别, 是微生物活动的重要证据以及陆相生物危机期的重要沉积记录, 为研究研究区以及豫西陆相二叠纪— 三叠纪之交的陆地生态系统以及微生物与后生生物的演化规律研究提供了新的资料。
荥阳位于河南中部, 处在中朝古陆华北地台南部区, 嵩箕台隆的东延部分的北侧, 属于豫西地层分区的嵩山小区[23]。主要出露地层为晚二叠世孙家沟组和早三叠世刘家沟组(图1)。
剖面位于荥阳市崔庙镇, 剖面露头较好, 上下地层接触关系清楚。孙家沟组厚度约230 m, 由下而上分为3部分, 下部主要为紫红色泥岩(含钙质结核), 夹少量灰色薄— 中层细粒长石石英砂岩, 发育单斜层理; 中部主要为灰绿色、灰黄色泥岩, 夹薄层钙质粉砂岩、灰岩或灰岩透镜体; 上部为灰黄色厚层中细粒长石石英砂岩, 夹核形石灰岩及暗紫红色泥岩。刘家沟组厚度约55 m(未见顶), 主要为紫红色中厚层、中细粒长石石英砂岩, 发育楔状交错层理、板状交错层理、波痕等(图1)。研究区整体为河流— 湖泊沉积环境, 孙家沟组主要为三角洲平原及三角洲前缘沉积, 刘家沟组主要为三角洲前缘沉积[24~26]。
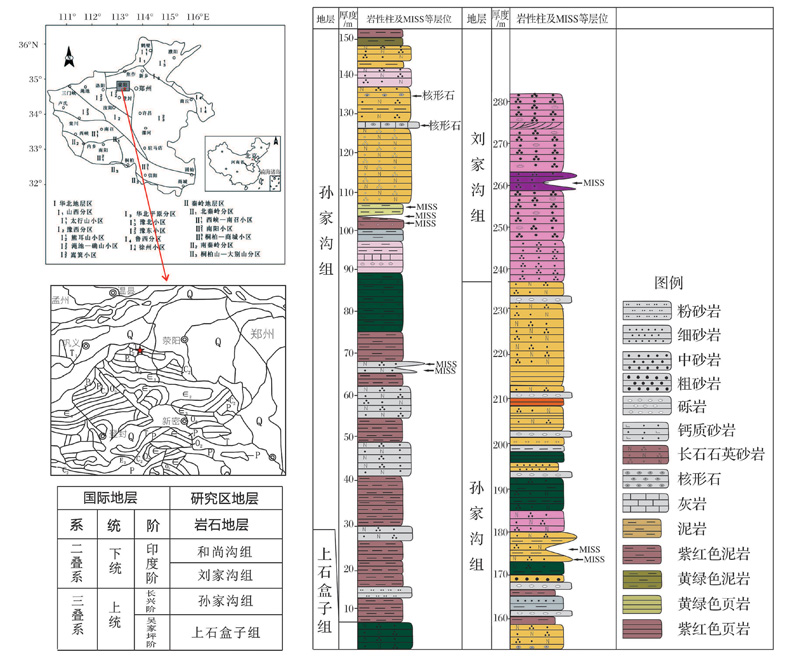 | 图1 豫西荥阳地区地质背景简图及地层序列Fig.1 Geological setting of study area and stratigraphic succession Premian-Triassic boundary in Xingyang, western He’ nan |
荥阳地区的MISS, 主要分布在砂泥互层的灰白色长石石英砂岩或中厚层紫红色中— 细粒石英砂岩层面, 砂岩中多发育楔状、板状交错层理、波痕等沉积构造。MISS主要分两大类, 孙家沟组灰白色的MISS和刘家沟组紫红色的MISS。与豫西鲁山地区中元古代海相地层发现的MISS有较多异同[7], 研究区石英砂岩成熟度不高, 各种类型MISS共生、混生的多, 单个类型发育的少。
有关微生物成因构造的分类, 国内外地质学者已有过多种论述[2~4, 6, 7, 20]。根据Schieber[3]的分类方案以及野外观察, 将研究区MISS分为席生长相关构造(Mat growth feature), 席破坏相关构造(Mat destruction feature)以及席腐烂相关构造(Mat decay feature)3类[7, 20]。研究区孙家沟组上、中、下段均发育有 MISS, 大多发育在中细粒、粉砂岩与泥岩的接触面上, 类型多样, 以微生物席生长相关构造、微生物席破坏相关构造为主, 以席破坏构造中的脱水裂痕最多。刘家沟组发现的MISS主要分布于该组的下部, 大多发育在紫红色中细粒石英砂岩与泥岩的接触面上, 多与波痕伴生, 主要类型为微生物席破坏相关的构造, 局部有微生物席腐烂相关构造。
3.2.1 微生物席生长构造
微生物生长相关构造是微生物的生长速率、方式及方向发生改变, 在水体和沉积物附近而留下的构造[6, 7]。常见的生长相关构造主要为变余波痕, 瘤状、簇状突起, 网状生长脊等[6, 20]。
研究区发现的微生物席生长构造主要有以下3种:
(1) 瘤状突起。研究区瘤状突起又可分为3种类型, 一是灰白色小瘤状突起(图2a), 该突起直径0.2~0.9 cm, 瘤高0.1~0.6 cm, 大小不等的分布于砂岩大碎片表面上(红色箭头指示处), 砂岩表面具光滑的微生物膜。二是灰白色小瘤状突起与似圆块疙瘩状突起共生(图2b), 小瘤状突起(红色箭头所示处), 直径0.2~0.6 cm, 瘤高0.1~0.5 cm, 多簇生, 既有单独存在的, 也有生长在大疙瘩状突起上的。似圆块疙瘩状突起多呈不规则状(黑色箭头所示处), 偶有方块状突起, 直径2~6 cm, 突起高0.5~1 cm不等, 2种突起表面均有光滑的微生物膜。三是暗紫红色瘤状突起(图2c, 红色箭头所示处), 直径0.4~1.2 cm, 瘤高0.1~0.3 cm, 多呈圆顶状隆起, 表面残留有深紫红色微生物氧化膜。微生物席具有多层结构, 多为上下2层, 上层以球状菌为主, 下层则主要为丝状菌为主, 上层较致密且具有强韧性和不透气性; 瘤状突起可能是由下层菌光合自养作用排出的气体未能穿透上部菌层顶起而形成的构造[16, 20]。该类构造在现代微生物席环境中较为常见[6]。
(2) 隆脊构造(图2d)。由于微生物席的局部过量生长导致侧向增长形成的隆脊[6, 7]。研究区发现的隆脊构造略突出于砂岩层面, 脊宽0.2~0.5 cm, 相对较规则, 向两端收敛歼灭, 围成狭长近纺锤形, 由于形似纺锤形, 多被误认为是纺锤状脱水裂痕。但与破坏构造里的纺锤状脱水裂痕不同, 一般隆脊构造相对较小, 且脊是从内向外生长突起, 脊与宿主岩石是一体的; 在纵切面上, 隆脊具典型的倒“ U” 顶结构, 宽0.5~2 cm, 高0.5~1 cm, 隆脊顶面不发育裂隙[6, 7]。但也有人认为该类构造由埋藏的有机质腐烂形成的气体排出而形成[27]。
(3) 变余波痕(图2e, f)。微生物席通过稳化、捆绑等作用, 使原沉积表面的沉积形态不被强水流所改造, 所形成的杂乱、多向的与微生物席生长相关的波痕形态构造, 称作变余波痕[12, 28]。研究区的波痕构造, 发育多、分布广, 其中变余波痕主要发育在孙家沟组下段和中段, 波痕指数和波痕方向均有明显的不同及差异, 显示出微生物席通过稳化、捆绑等作用阻止水流改造而使方向发生了变化。其中一些变余波痕层面, 部分被微生物席覆盖, 原始波痕已不可见(图2f)。
3.2.2 微生物席破坏构造
微生物席破坏相关构造是指因气候原因微生物席暴露于沉积表面, 受物理、化学因素作用形成的干裂、卷曲、褶皱、变形、脱水等沉积构造, 如脱水干裂构造、卷边构造、席破坏碎片及部分微皱痕构造[6, 7, 16, 20]。各种裂隙构造是最典型破坏相关构造, 因暴露时间长短、保存条件不同产生的从单一的纺锤状到多边形至网状的脱水裂痕[7, 20]。它们常出现在砂岩中, 因此, 也被称为“ 砂裂” 。破坏相关的构造是荥阳地区地层中, 发育较多, 容易识别的沉积构造, 包括纺锤状脱水裂痕、多边形脱水裂痕、树枝状脱水裂痕、曲形脱水裂痕、似正弦状脱水裂痕和微生物席砂片。
(1) 纺锤状脱水裂痕(图2g)。这种裂痕在研究区较其他已见报道地区发育少[6, 7, 20], 多呈纺锤状, 在波痕波谷中或非波痕处都有发育。该构造多是席暴露后脱水、干裂而形成的, 也有因多次脱水、干裂作用后重叠形成的构造[7, 20]。该种构造主要发育在孙家沟组的中、下段。
(2) 多边形脱水裂痕(图2h)。这种构造在研究区发育较多, 也较容易识别, 根据形态、大小可再分为多种类型, 这些类型内部又分有各种次级分支, 这些次级分支主要为多边形状、树枝状、发散状等; 有些类型微生物席较厚, 裂痕较大, 内部无分支, 形状较规则。研究区发育的大型多边形脱水裂痕(图3a), 多呈四边形, 砂脊宽0.2~1.2 cm, 突出层面0.05~0.5 cm, 脊长3~10 cm, 在纵切面上砂脊表现为附着在层面上的椭圆形脊状体。这些构造因多边形内的微生物席腐烂而消失, 只留下凸出于层面的多边形脊; 它们与泥裂形态较相近, 过去也有叫作“ 假泥裂” [6]。
 | 图2 豫西荥阳地区陆相二叠纪— 三叠纪之交的微生物成因构造 (a)灰白色小瘤状突起; (b)灰白色小瘤状突起与似圆块疙瘩状突起共生; (c)暗紫红色瘤状突起; (d)隆脊构造; (e~f)变余波痕; (g)纺锤状脱水裂痕; (h)多边形脱水裂痕Fig.2 Microbially Induced Sedimentary Structures in terrestrial Premian-Triassic boundary in Xingyang (a)The offwhite small growth postulas; (b)The symbiotic offwhite small growth postulas and lump; (c)The dark purplish-red growth postulas; (d)Growth rigdes; (e~f)Palimpsest ripples; (g)The spindle-shaped cracks; (h)The polygonal desiccation cracks |
(3) 曲形脱水裂痕(图3b)。这是一种特殊的多边形脱水裂痕。研究区发育较少, 主要位于孙家沟组中段。砂脊弯曲程度较大, 交互形成环状、近环状等, 砂脊宽 0.2~1 cm, 高0.2~0.5 cm。砂脊明显附着于层面, 并突出于两侧围岩, 可与其他类型砂质裂痕共生, 砂脊发育有一级、二级、三级等不同级次, 各级次发育裂痕的大小有着不同的差异。
 | 图3 豫西荥阳地区陆相二叠纪— 三叠纪之交的微生物成因构造 (a)大型多边形脱水裂痕; (b)曲形脱水裂痕; (c)树枝状脱水裂痕; (d)似正弦状脱水裂痕, (e)灰白色微生物席砂片; (f)紫红色微生物席砂片; (g)砂火山构造; (h)似雨痕构造Fig.3 Microbially Induced Sedimentary Structures in terrestrial Premian-Triassic boundary in Xingyang (a)Large polygonal desiccation cracks; (b)The curved desiccation cracks; (c)The dendritic desiccation cracks; (d)Manchuriophycus; (e)The offwhite microbial sand chips; (f)Purplish-red microbial sand chips; (g)Sand volcano; (h)Rain-print |
(4) 树枝状脱水裂痕(图3c)。形状像树枝状的脱水裂痕构造, 由较粗大的主枝与细小的分支组成, 其中较小的分支可再分成更细小的分支。一般主干裂痕宽0.5~1.5 cm, 脊高0.05~0.3 cm, 长度可达10 cm, 分支裂痕宽0.1~0.8 cm。主要发育于孙家沟组中、下段, 规模大小不等。
(5) 似正弦状脱水裂痕(即“ Manchuriophycus” 构造)(图3d)。是微生物席脱水裂痕的一种特殊构造, 由生长在波谷的相对较厚的微生物席干裂产生, 在波谷内多呈似正弦曲线状蜿蜒分布[6, 7]。荥阳地区发现的此类构造在波谷内呈纺锤形或似正弦形, 在波谷内延伸较长(红色箭头所示处), 可达10 cm, 个别裂痕穿越波峰。这种裂痕在波谷内限制性发育, 表面微生物席会选择性的生长。似正弦状脱水裂痕是元古代陆源硅质碎屑潮坪相中常见的一种构造, 以前多被学者定义为遗迹化石, 在我国贺兰山黄旗口组、豫西云梦山组均有报道[6, 7]。
(6) 微生物席砂片。是微生物席被物理、机械作用(如干旱、强水流或风力等)完全破坏, 席及其包裹的沉积物一起被搬运、改造, 形成变形、重叠的、大小不等的席片[6, 7]。许多席片是强水流从高韧性的席上撕下而形成的, 席的韧度和物理作用的强度决定了席片的大小[7, 12, 29]。荥阳地区砂片构造可分为2种类型:一是灰白色砂片(图3e), 主要发育在孙家沟组, 砂片大小不一, 小的宽约0.3 cm, 大的宽约6 cm; 形状多呈多边形状, 也有四边形。砂片多脱落, 其下具有灰白色微生物膜。二是紫红色砂片(图3f), 主要发育在刘家沟组, 规模较大, 也有小规模的, 砂片最大可达4 cm宽、5 cm长, 形状呈三角形、四边形、多边形等, 砂片也多脱落, 其下具紫红色微生物氧化膜。其中, 一些脱落砂片被冲刷成凹坑, 微生物氧化膜也被冲刷掉。一些学者认为这层紫红色膜可能是微生物膜(biofilm)中, 蓝细菌诱发黄铁矿沉淀, 发生氧化作用的残留物[1]。这种砂片构造在南非新元古界的Nama群、摩洛哥Wanimzi群、澳大利亚Pound超群[16], 北京南口古元古界大红峪组、豫西鲁山中元古界云梦山组、山西黎城中元古界常州沟组等均有发现[7, 17, 20]。但这些砂片都比荥阳地区发现的砂片小。
3.2.3 微生物席腐烂相关构造
微生物席腐烂相关构造是指被埋藏的微生物席腐烂分解而产生的气体逃逸出来或未逃逸出来而在沉积物表面形成的各种构造[4, 6, 16]。由于强韧性、不透气性的微生物席阻碍了其下面砂质沉积物与大气或水之间的气体流通, 致使由埋藏的有机质腐蚀或腐烂形成的气体, 向上逃逸形成的拱起、隆起状的气隆构造; 而在席相对比较薄或气体压力相对较大的情形下, 气体则穿破席表, 形成中空、凹陷的“ 砂火山” 构造[3, 6]。
荥阳地区发现的这种构造较少, 主要为砂火山构造和似雨痕构造。①砂火山构造(图3g, 红色箭头所示处), 主要发育在刘家沟组, 与波痕共生, 形似凹陷火山口状, 可能是气体穿破微生物席后隆起部位凹陷形成的。荥阳地区的砂火山构造, 因风化作用影响而呈现清晰的圆状或椭圆状, 中部直径0.3~1 cm, 环绕的外壁厚1~2 mm。外壁富含泥质或有机质, 粒度比内壁细, 易受风化影响形成浅沟, 其可能是风化的微生物席层留下的[7]。②似雨痕构造(图3f, 红色箭头指示处), 主要发育在孙家沟组, 与多边形裂痕共生, 但是该裂痕多被侵蚀呈凹陷裂缝。其与砂火山的区别是中间气隆被侵蚀成凹坑, 可能与该层的灰白色长石石英易被风化有关。
通过磨片、切片, 偏光显微镜、实体镜、扫描电镜观察, 对荥阳地区MISS的微观特征和成因进行了探讨和分析:
(1) MISS脊纵切面显示U或大U型, 且一些样品中脊底部发育有暗色黏土矿物。研究表明, 泥裂是泥质沉积物的脱水作用所致, 是一种单纯的物理作用, 且纵向切割比较深, 会形成典型的“ V” 字型。而荥阳地区一些脊和宿主岩石均为砂岩的MISS, 在纵切面显示为U或大U型, 且在U型脊底部有一层暗色物质(图4a, 红色箭头指示), 通过能谱测定, 这些暗色物质成分主要为O, Si, Al等, 为黏土岩的成分, 可能为伊利石等(图4b)。一般来讲, 砂岩颗粒没有黏性, 是不会收缩的, 很难形成V型的泥裂[30], 这表明研究区MISS与泥裂有着本质的区别, 应该是研究区微生物席暴露干燥、脱水而形成的, 这也是微生物成因构造区别于泥裂的一个标志[31]。
(2) MISS脊处颗粒与围岩颗粒明显不同, 且边界明显。研究区一些样品中, MISS脊处的颗粒与围岩有明显的不同, 且边界清晰(图4c, 红色箭头所示)。脊与宿主岩石均为砂岩, 脊处砂岩颗粒较为纯净, 胶结好, 而脊边缘宿主岩石颗粒中暗色物质较多, 胶结程度不如脊处颗粒。脊处纯净的颗粒可能是微生物席干裂后, 上层沉积砂岩颗粒充填以及微生物活动参加较少所致, 而宿主岩石颗粒间暗色物质则为微生物席腐烂残留被黏土矿物层代替形成的。
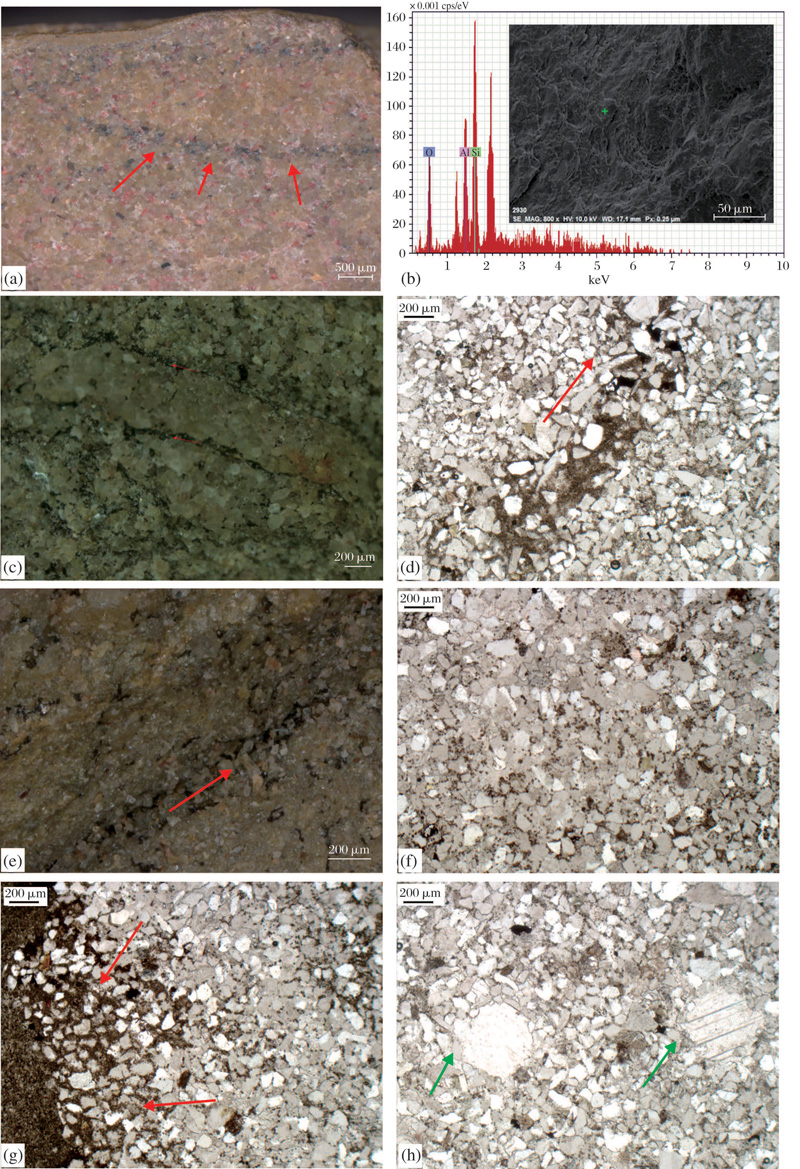 | 图4 豫西荥阳地区陆相二叠纪— 三叠纪之交的微生物成因构造镜下微观特征 (a)大U型脊(红色箭头所指); (b)暗色黏土矿物(SEM); (c)脊及围岩石英颗粒(红色箭头所指); (d)石英颗粒定向排列(红色箭头所指); (e)脊下石英颗粒定向排列(红色箭头所指); (f)黏土矿物广泛分布在岩石颗粒间隙; (g)脊边缘颗粒间较多的黏土矿物(红色箭头所指); (h)石英次生加大现象(绿色箭头所指)Fig.4 Microscopic feature of Microbially Induced Sedimentary Structures in terrestrial Premian-Triassic boundary in Xingyang (a)Large U ridges(red arrows); (b)Dark clay minerals(SEM); (c)The quartz grains in ridge and host rocks(red arrows); (d)Directed arrangement of quartz grains (red arrows); (e)Directed arrangement of quartz grains under ridges (red arrows); (f)Clay minerals in quartz grains; (g)More clay minerals in edge of rigde(red arrows); (h)Secondary enlargement of quartz(green arrows) |
(3) 在多个样品中显示石英颗粒或暗色矿物条带的定向排列。在薄片、实体镜下均发现有一些样品中石英颗粒的定向排列, 在图4d中, 可以看到长条状的石英颗粒沿着暗色物质条带定向排列(红色箭头方向), 条带周围石英颗粒磨圆度较差, 棱角分明。实体镜下, 在脊边界下也显示出暗色矿物条带的石英颗粒的定向排列(图4e, 沿红色箭头方向)。该特征是微生物席、微生物膜或丝状体有选择性地捕获、障积矿物颗粒形成的, 上部定向排列的石英颗粒是微生物席或膜抵制水流或气体等改造的结果, 这被认为是微生物席或膜存在的重要证据之一, 同时也是识别硅质碎屑岩中微生物席存在的重要标志[6~7, 20, 32]。
(4) 石英颗粒空隙间普遍存在暗色的黏土矿物, 且MISS脊周围明显增多。薄片及实体镜镜下发现, 岩石石英颗粒间普遍存在着黑色的黏土矿物(图4f), 在脊或黏土条带边缘等地方会明显增多(图4g, 红色箭头指示), 也有些存在着石英颗粒次生加大现象(图4h, 绿色箭头指示)。说明在条件适宜的情况下, 微生物在颗粒周围生长蔓延并捕获矿物颗粒来充填缝隙并在缝隙处堆积形成的。同时也发现, 有次生加大现象的石英颗粒周围基本上无暗色黏土矿物保留或保留较少, 说明在后期成岩过程中, 在温度和压力的影响下, 微生物残留物或有机质以降解或其他形式消失掉, 这也许是微生物化石较难保存的一个原因。
(5)一些样品中, 在脊处或纵切面上存在着黑色黏土矿物条带层, 边界明显, 存在不连续、亚平行的云母颗粒。从图5a, b中, 可以清晰看出脊的边缘发育黏土矿物条带, 且有疑似定向排列的丝状体。在实体镜和扫描电镜下可以看到纵向生长构造多层的黏土矿物层(图5c, d, 图5c为黏土矿物层的局部放大), 且一些黏土矿物层出现了缝隙。在破坏构造上脊处也发现有定向排列黏土矿物条带(图5e, f)。Schieber[3]指出微生物席或黏土矿物层均能暴露、收缩形成微生物成因构造。样品中, 多层的黏土矿物层表明了微生物席的多期频繁的生长, 干燥脱水后被黏土矿物层所替代, 缝隙的出现可能为微生物席残留物腐烂或黏土矿物被风化所致。破坏构造纵向层中、脊处以及在生长构造样品中均发现有不连续、近平行、定向排列的云母颗粒(图5g和h, 绿色箭头指示), 且在生长构造一些样品中颗粒较大, 也显示浅黄色黏土岩条带平行, 其中2个云母颗粒(红色箭头处)可能被微生物席生长所携带而向上迁移(图5i)。这些特征被认为与微生物席阻碍、捕获和捆绑有关[2, 11]。一些样品中, 显示为疑似云母颗粒水化形成的绢云母(图5j, 红色箭头处), 这也是被认为微生物成因的一种证据。一些学者指出, 绢云母是微生物化石层中普通的矿物, 经常出现在灰色层的石英胶结物中, 可能是原始的黏土矿物被微生物群落捕获、黏结后, 在席层内经过物理、生化作用的结果, 这些过程又影响了层内黏土矿物的形成[7, 33, 34]。
综上所述, 根据研究区MISS的宏观形态、类型以及U型构造、颗粒定向排列、云母颗粒不连续亚平行等5个具体的微观特征, 我们认为研究区的MISS是微生物群生命活动的产物, 是微生物与沉积环境相互作用形成的沉积构造。
 | 图5 豫西荥阳地区陆相二叠纪— 三叠纪之交的微生物成因构造镜下微观特征2 (a)黏土矿物条带; (b)疑似定向排列的丝状体; (c~d)生长构造的多层黏土矿物层和缝隙(红色箭头所指); (e~f)破坏构造的黏土矿物条带; (g~h)不连续、近平行的云母颗粒(绿色箭头所指); (i)被微生物席生长携带、捕获的云母颗粒(红色箭头所指); (j)绢云母(红色箭头所指)Fig.5 Microscopic feature of Microbially Induced Sedimentary Structures in terrestrial Premian-Triassic boundary in Xingyang (a)Clay minerals stripe; (b)Directed arrangement of microbial filaments; (c~d)Clay minerals laminae and gap of the growth structure(red arrows); (e~f)Clay minerals stripe of the destruction structure; (g~h)Parallel and discontinuous mica grains(green arrows); (i)Trapping mica grains by Microbial mats(red arrows); (j)Sericite(red arrows) |
MISS在不同的地史时期均出现过, 特别是在中、新元古界地层中广泛分布, 众多学者对其进行了系统的研究和论述[1~20]。与前寒武纪相比, 由于后生生物的影响, 显生宙的MISS逐渐减少, 仅出现在个别极端时期和极端环境中。研究表明, 作为碎屑岩典型微生物活动形成的MISS, 在显生宙的出现往往代表一种比较特殊的生态环境, 而MISS出现在二叠纪— 三叠纪之交的豫西地区的陆相环境中, 可能是一个特殊的灾后陆相生态环境。因此, 用研究区MISS来揭示二叠纪末生物大灭绝对陆地生态环境影响以及具体反映有重要的生态指示意义。
二叠纪末, 受秦岭— 大别山构造带与华北板块南缘碰撞影响, 华北石炭— 二叠纪克拉通盆地南部的陆表海已经完全退出华北, 盆地南缘出现了新的物源隆起区, 古沉积斜坡由原来的向南倾斜转变为向北倾斜[35], 豫西地区古地理、古气候发生重大变化, 发育了一套灰白、紫红及灰紫色为主的干热气候条件下的河流— 湖泊相沉积体系(即孙家沟组和刘家沟组)[36]。在研究区二叠纪— 三叠纪之交地层中没有发现后生生物实体化石、孢粉化石以及遗迹化石等, 特别是在孙家沟组中部至刘家沟组下部的地层中; 同时在豫西其他剖面, 仅在晚二叠统孙家沟组下部(宜阳)、刘家沟组中部(宜阳和登封)发现了遗迹化石以及刘家沟组上部(登封)发现有孢粉化石[37~39], 显示出孙家沟组中部至刘家沟组下部的这段地层中化石极其稀少。Chu 等[39]在研究豫西宜阳二叠纪— 三叠纪之交的地层时, 发现孙家沟组中部至刘家沟组下部生物多样性明显偏低; 而王自强等[40]也指出华北地区二叠纪末的植被显示出严重的退化演替。而恰恰在此区间, 研究区的MISS却最为发育, 伴生发育了多层的不规则厚薄不均、大小不一、向上增多、排列相对规则的砾岩层, 以及含大量的结核钙质土壤层, 这些现象显示出地表植被的荒芜、水土流失严重和气候的炎热、干燥[26, 41]。诸多现象均表明, 研究区以及豫西地区二叠纪— 三叠纪之交的沉积为一种特殊的陆相沉积环境, 即二叠纪末灾变事件使研究区及豫西地区后生生物急剧减少, 为微生物的大量繁殖创造了有利条件而形成的沉积环境。我们认为研究区这套以MISS为主的沉积组合可能代表着二叠纪末大灭绝后在陆地上残留的一个简单而特征明显的生态系统。该陆相生态系应具有如下特征:①以微生物为基础; ②后生生物少、单调; ③砾岩、钙质结核等特殊沉积发育; ④多出现在砂泥互层中。这个生态系统表明了微生物参与的物理沉积过程, 蕴含了地球生物学和沉积环境两方面的信息[42], 通过对它以及MISS为主的沉积组合的研究, 可以为古生代— 中生代地质转折期的陆相生物绝灭复苏问题研究提供新资料, 而且还可以弥补大绝灭后古生物化石记录在地层中分布与保存的不足, 成为独立于化石记录之外的生态变化指针[43]和二叠纪— 三叠纪陆地生态危机环境的指示器[44]。
全球性的二叠纪末生物大灭绝事件, 不但重创了海洋生态系统, 导致海洋约95%的物种绝灭[45], 而且造成了陆地生态系统的崩溃, 使约70%的陆地物种灭绝[46]。但因陆相二叠系— 三叠系界线地层因化石不易保存、稀少、单调, 使陆相二叠系— 三叠系界线地层及早三叠世生物复苏研究较海相相对薄弱[47]。近年来, 豫西地区[26, 39, 42, 44]和滇东黔西地区[41]的二叠系— 三叠系界线地层研究取得了较大进展, 但仍有不足。二叠纪末生态危机后的陆相宏体化石不均匀分布、可能的非原地埋藏以及剖面上不连续的特征, 使化石数据的可靠性存在争议, Gastaldo 等[48]就曾对南非Karoo盆地的植物化石记录提出质疑。微生物成因构造具有原地保存、容易发现、特征明显和蕴含生物和环境信息等优势, 能反映陆相微生物生态系与沉积环境的变化特点[32, 42, 44], 能为二叠纪末生态危机后陆相生态系统的演变和沉积环境变化提供重要依据。我们通过荥阳、宜阳[39, 42, 44]和登封[38]的MISS与遗迹化石、植物化石对比研究, 发现地层中MISS和遗迹化石、植物化石以及与环境之间存在着特殊的此消彼长的关系。在空间上表现为:MISS发育, 则遗迹化石、植物化石减少; 遗迹化石、植物化石增多, 则MISS减少或消失; 仅在局部地方共存。表明二叠纪末大灭绝后, 研究区陆地生态环境恶化, 后生生物遭到重创, 微生物大量繁殖并逐渐占据陆地环境, 形成了大量的微生物成因构造。早三叠世, 后生生物开始逐渐复苏, 从与微生物成因构造共存并以微生物席为食, 到逐渐增多, 直至占据正常陆地环境, 显现出陆地生态系统逐步复苏。那么, 我们是否可以根据地层中MISS与后生生物化石此消彼长的耦合变化的发育规律, 将研究区陆相MISS与豫西其他地区陆相MISS进行地层对比(甚至可以与华南地区二叠纪末大灭绝后发育的微生物岩的地层进行对比), 根据其发育的层位、变化规律来卡定界限范围, 结合沉积学、地球化学等证据, 来进行界线地层的研究呢?这为二叠纪— 三叠纪之交的地层划分对比提供了一种新思路。再者, 研究区陆相MISS的出现与同为微生物席形成的华南地区二叠纪— 三叠纪之交微生物岩相对应, 也能为二叠纪末灾变事件后的微生物席在全球的空间分布提供了新认识。因此, 研究区陆相微生物成因构造为二叠纪末灾变事件地层划分对比和微生物席的全球空间分布提供了新的研究资料。
一些学者指出[49~51], 寻找灾变事件保存的古代微生物席是今后微生物岩研究的主要任务之一, 而微生物成因构造出现在豫西陆相二叠纪— 三叠纪之交的地层中, 其特殊的层位有着特殊的意义, 虽然这方面的研究[39, 42, 44]还不够成熟, 也不够全面, 仍然有很多的研究工作要做, 但是作为重大灾变期出现的陆相碎屑岩微生物成因构造, 对于全面了解二叠纪末豫西以及全球的陆相生态系统变化和复苏有着重要的意义。
荥阳地区发育了形态不同、识别容易的微生物成因构造, 可分为3大类, 微生物席生长构造、微生物席破坏构造和微生物席腐烂构造, 11个小类, 瘤状突起、隆脊构造、变余波痕、纺锤状脱水裂痕、多边形脱水裂痕、树枝状脱水裂痕、曲形脱水裂痕、似正弦状脱水裂痕、微生物席砂片、砂火山构造和似雨痕构造。通过镜下观察发现其具有5个主要特征:①脊纵切面显示U或大U型, ②脊处石英颗粒相对围岩纯净且胶结程度稍高, ③脊处石英颗粒具定向排列且磨圆度较差, ④岩石石英颗粒间发育暗色黏土矿物, 且脊处较多, ⑤脊处或纵切面有黏土矿物条带, 一些云母颗粒存在不连续、近平行的现象。这些特征表明了其为微生物群生命活动的产物, 是微生物与沉积环境相互作用形成的沉积构造。
通过研究我们认为研究区发现的陆相微生物成因构造是一种发育普遍、容易识别的陆相特殊沉积构造, 代表着以微生物为基础、后生生物少、钙质结核发育等特殊沉积的二叠纪末灾后陆地生态系统, 其蕴藏的微生物演化信息以及与后生生物演化的关系的沉积记录, 为海陆相微生物席的空间分布提供了新的资料, 对全面了解二叠纪末豫西陆相生态系统有着重要的意义。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|