

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
海洋碳汇对气候变化的响应与反馈
[焦念志1  , 李超
, 李超2 , 王晓雪3 ]
, 李超|
|


作者简介:焦念志(1962-),男,山东潍坊人,中国科学院院士,主要从事海洋生态过程与环境效应研究.E-mail:jiao@xmu.edu.cn
海洋碳汇对气候变化的响应与反馈是一个系统的科学命题,也是当前国际地球系统科学领域的前沿热点问题,需要通过微观与宏观结合、古今链接、多学科交叉融合进行深入研究。在我国科学家发起的海洋生物地球化学“戈登科学前沿论坛”(Gordon Research Conferences,GRC)首届论坛上,以海洋生物泵(Biological Pump,BP)、微型生物碳泵(Microbial Carbon Pump, MCP)以及碳酸盐泵(Carbonate Counter Pump,CCP)等海洋储碳机制为核心,深入研讨了海洋碳汇的过程与效应,起到了引领国际海洋学科发展方向的作用。国内学界也积极行动起来,在第四届地球系统科学大会上组织了海洋碳汇专题,从古海洋碳汇、现代海洋碳循环及海洋碳汇的生物海洋学过程3个方面开展研讨。海洋微型生物生态学过程与海洋碳汇以海洋浮游植物、细菌、古菌、病毒以及不同微型生物间的互作为切入点,探讨了微型生物的储碳和固碳作用的过程及其与全球气候变化的关系。古海洋碳汇方向的报告在时间尺度上跨越了从18~8亿年前的中元古代到距今2.5 Ma的第四纪,涵盖了包括古海洋碳汇形成的古海洋环境、古海洋碳汇的生态环境效应等前沿科学问题;古海洋碳汇的报告为现代海洋碳汇研究提供了有益的借鉴,并有助于本领域科学家对海洋碳汇的历史演化观的认识。现代海洋碳循环过程方面,专题报告结合时间梯度和空间梯度,以南海珊瑚礁碳循环源汇争议为代表,探讨了碳循环中的初级生产力、溶解有机碳的来源与有机碳的降解等过程,对现代海区和全球变化背景下海洋的源汇评估提出了新的想法与研究方向。
The response and feedback of ocean carbon sequestration to climate changes is a international hot topic and requires large spatial/temporal scale, collaborative and multi-disciplinary research. In the first conference of GRC Ocean biogeochemistry, scientists focus on three biologically-driven ocean carbon pumps (Biological Pump, BP; Microbial Carbon Pump, MCP; Carbonate Counter Pump, CCP) and their environmental and climate consequences. As a sister meeting in China, we organized the session to show the efforts and progress of ocean carbon sequestration of Chinese scientists. The microbial ecological processes of phytoplankton, bacteria, archaea and viruses and interactions between them were highlighted in the session. Use coral reefs in the South China Sea as an example, the presenters and the participants come to an agreement that interdisciplinary collaborations are needed to ensure a comprehensive understanding of the interactions between microbes and their geochemical environment and the consequences of microbial processing of carbon on outgassing of CO2 and carbon sequestration. The session also have presentations focusing on paleo-environmental reconstruction for carbon sinks as well as their paleo-ecological effects in ancient oceans with time spanning from the 1.8~0.8 Ga Proterozoic to the 2.5 Ma Quaternary. These talks provide specific geological cases for the oceanic carbon sink research and convey the emerging geological view of paleooceanic carbon sinks to the research community of modern ocean carbon sinks. As a summary, the discussion in this session of biological pump, microbial carbon pump and carbonate counter pump shows the latest research progress and future development trend in this field.
以地球系统科学为基础的全球变化研究, 需要将海洋与陆地统筹考虑、协同创新, 海洋碳汇是全球变化研究“ 绕不过去的坎” 。同时, 海洋碳汇具有显著的生态环境效应, 包括海洋酸化、缺氧化、富营养化、赤潮等在内的一系列海洋生态环境问题, 都与海洋碳汇密切关联。海洋碳汇不仅在现代, 而且在地质历史时期也发挥了举足轻重的作用。除了人们熟知的石油天然气资源的物质基础外, 海洋碳汇与地球环境变化和生命演化过程密不可分。大气氧化、雪球地球、生命大爆发和数次生物大灭绝, 都与海洋碳库在地质时间尺度上的变动息息相关。因此, 海洋碳汇已成为地球科学和全球变化科学中一个新兴的、迅速发展的前沿领域。2015年, 党中央、国务院印发的《生态文明体制改革总体方案》 明确要求建立增加海洋碳汇的有效机制。为此, 必须首先阐明海洋碳汇的形成过程与调控机理。
认识海洋碳汇对气候变化的响应与反馈是一个系统的科学命题, 需要通过微观与宏观结合、古今链接、多学科交叉融合进行深入研究。
最近, 由中国科学家发起创立的美国 “ 戈登科学前沿论坛(Gordon Research Conferences, GRC)” 的“ 海洋生物地球化学与碳汇” 永久论坛给出了一个成功范例。 海洋生物泵(Biological Pump, BP)、微型生物碳泵(Microbial Carbon Pump, MCP)以及碳酸盐泵(Carbonate Counter Pump, CCP) 等海洋储碳机制及其主要调控因子的深入研讨, 展现了该领域的最新研究进展和未来发展趋势。例如, 分子生物学新技术研究表明微型浮游生物 (pico-, nano-plankton)对BP有重要贡献; 弱光层水体中异养细菌而非浮游动物主导了呼吸作用; 沉降有机质在海洋中层水中的矿化速率在不同纬度上的变化和海洋温度的分布有极大关系, 低纬高温水层活性有机质组分快速降解, 难降解组分被输送到海洋底部, 在高纬低温水域降解较慢, 但可以持续到很深的水层。未来全球变暖情景下MCP相对作用增加。今后的深入研究更趋向于与时间空间尺度上BP和MCP的耦合关系, 以及通过 “ 组学” 手段更深入地了解不同生物类群之间相互作用如何影响有机质在海洋中的转化。全球碳循环有各种尺度, 在冰期和间冰期旋回中, 主要由地外过程驱动。在一个相对稳定的时期里, BP和MCP等则对气候变化的影响举足轻重[1]。
与国际GRC相呼应, 国内的地球系统科学大会提供了一个陆海统筹、学科交叉、古今结合、协同创新的中文学术交流平台, 第四届大会的“ 海洋碳汇对气候变化的响应与反馈” 这一专题吸引了来自中国地质大学、同济大学、厦门大学、中国海洋大学、北京大学、山东大学、暨南大学、广西大学、东华理工大学、中国科学院南海海洋研究所、青岛生物能源与过程研究所、国家海洋环境监测中心等十多个高校及科研院所的15位专家与会报告、百余学者济济一堂热烈讨论。与会专家从古生物学、海洋化学、海洋地质学、古气候学、海洋生物学等多个角度进行交流和研讨, 本文对研讨成果进行简要总结。
古海洋中碳汇的研究对于理解现今海洋碳汇形成和预测其未来的碳汇趋势及其环境生态效应都具有重要意义[2]。本专题的一个鲜明特色就是对地质历史时期海洋碳汇的研究, 体现了古今结合研究海洋碳汇的新方向。本专题古海洋碳汇报告在时间上涉及了从18~8亿年前的中元古代到距今2.5 Ma的第四纪, 在研究主题上涉及了从古海洋碳汇形成的古海洋环境到古海洋碳汇的生态环境效应等; 这些报告不仅为现代海洋碳汇研究提供了许多有益的借鉴, 而且也促成了本专题与会海洋碳汇科学家的新思想动向— — 海洋碳汇的历史演化观。
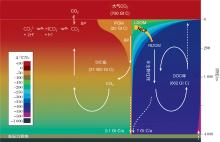
由于距今18~8亿年的中元古代在构造、生命演化等方面表现较为单调被称为“ 沉寂的十亿年” (Boring Billion)。然而这一时期具有气候温暖和海洋广泛的缺氧分层甚至硫化等特点[3, 4], 这些海洋条件理论上应该更加适合古海洋中碳汇的形成。北京大学的胡永云团队以此为突破口, 使用海气耦合气候模式(Community Climate System Model version 3, CCSM3)对这一时期古海洋洋流与层结程度给予研究。在模拟试验中, 他们使用比现在弱10%的太阳常数和两种重建的古大陆分布[5]:大陆偏近于高纬度, 主要分布在热带地区。通过对不同大气CO2浓度进行一系列模拟试验[6], 他们发现:中元古代大气CO2浓度是现今的20倍(7 100× 10-6), 地球的两极也没有永久性冰川的存在(图2)。在这种情况下, 古海洋温盐环流的强度以及海洋表层与深海物质交换效率比现在气候条件下的还高, 而深部海洋层结稳定性与现代海洋层结稳定性相当。这些研究结果表明中元古代的海洋物理条件并不适合大规模的海洋碳汇的形成。
晚新元古代全球大冰期(即“ 雪球地球” [7])之后的埃迪卡拉纪(635~541 Ma)不仅是后生动物诞生的时代[8], 而且在全球范围内记录了地质历史时期已知的最大碳同位素负偏事件(低至-12‰ )[9]。目前, 对于这一极端碳同位素负偏事件解释的一个重要假说是埃迪卡拉纪古海洋中存在一个大型溶解有机碳(Dissolved Organic Carbn, DOC)库; 随着埃迪卡拉纪古海洋的逐步氧化, 这一大型DOC库也逐步氧化, 释放出的同位素较轻的CO2导致了这一强烈无机碳同位素负漂移事件[10~14]。这一假说对于这一时期海洋化学特征和古气候条件有明确的暗示, 由此, 查明这一时期古海洋化学特征和古气候的波动对于验证大DOC库假说和深入理解这一时期古海洋大型DOC形成机制和其环境生态效应具有重要的意义。以中国地质大学(武汉)李超团队[15]为代表的研究人员以Fe-S-C多元的地球化学指标记录以及数学定量模型方法重建了我国华南埃迪卡拉纪古海洋的氧化还原空间分布。研究结果表明新元古代全球大冰期之后的埃迪卡拉纪古海洋具有缺氧分层的空间结构。常华进等[16]的报告也表明这一古海洋特征甚至延伸到了早寒武世。更为重要的是, 以空间硫同位素记录为基础的数学定量模型计算表明:在从极端碳同位素负偏开始前至负偏结束, 海洋硫酸盐含量整体上经历了一次由低到高再降低的过程, 并存在明显的从浅水向远洋减少的空间梯度[17]。这些结果表明当时古海洋氧化还原特征的确适合大型DOC库在深部海洋中积累和形成。李超等团队在此基础上使用了目前新兴的全新的温度代用指标— — 二元同位素温度计(clumped isotope thermometer)△ 47重建了埃迪卡拉纪无机碳同位素出现强烈负漂移及之前(635~551 Ma)的古海洋表层海水的温度变化[18]。结果表明:古海洋表层海水温度波动范围为18.3~62.1 ℃。进一步, 与无机碳同位素记录的对比表明:当无机碳同位素记录出现负漂时古海洋表层海水温度升高(最低43.6 ℃, 最高62.1 ℃, 平均温度46.2 ℃), 而当无机碳同位素记录未出现负漂时, 古海洋表层海水温度则较低(最低18.3 ℃, 最高41.8 ℃, 平均温度32.3 ℃)。这一结果符合大溶解有机碳库假说的预期, 也符合已知的埃迪卡拉纪中期Gaskiers冰期(约580 Ma)的出现(最低温度处), 因此该研究为大溶解有机碳库假说提供了独立的地质证据。研究结果表明微型生物碳泵(Microbial Carbon Pump, MCP)在古海洋碳汇的重要作用, 同时, DOC库演化影响了地球气候的变化。
 | 图1 海洋碳汇的主要生物学机制— — 海洋生物泵、微型生物碳泵以及碳酸盐泵(引自 “ 戈登科学前沿论坛” 的“ 海洋生物地球化学与碳汇” 永久论坛Logo)Fig.1 The main biological mechanism for ocean carbon sequestration— BP, MCP and CCP |
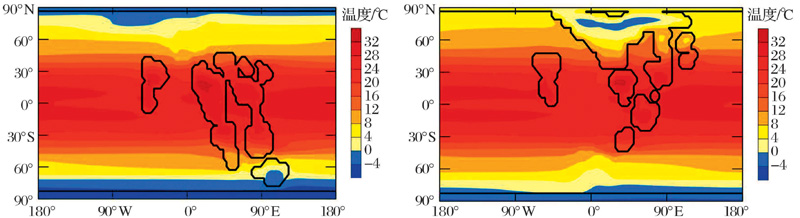 | 图2 模拟给出的20倍现代CO2浓度条件下(7 100× 10-6)2种不同大陆分布条件下的全球年平均温度分布 (a)大陆主要分布在赤道, 代表约14亿年前的陆地分布; (b)大陆主要分布在北半球高纬度, 代表约15亿年前的陆地分布Fig.2 Simulated surface air temperatures for two types of continients during the Boring Billion, The model is forced with 20 times present-day CO2 concentration (7 100× 10-6) and 90% present-day solar constant (1 230 W/m2) (a) Continients located at low latitudets for about 1.4 billion years ago; (b) Continents located at middle and high latitudes of the Northern Hemisphere for about 1.5 billion years ago |
2.52亿年前的二叠系— 三叠系界线是地质历史上的一条重要分界线, 既是古生代与中生代之间的时间分界线, 也是地球气候从冰室期向温室期转变的关键时期, 在该界线附近还发生了显生宙以来最大规模的生物灭绝事件, 随后生物经历了漫长和复杂的复苏过程。华南越来越多的证据显示, 这次生物大灭绝表现出两幕式, 并伴随两幕碳同位素负偏和微生物暴发, 显示了碳循环、微生物与生物危机之间的密切关系[19]。进一步的研究发现, 第二幕的生物危机可能更加严重, 表现在第二幕的生物危机破坏了存在约 2亿年之久的海洋生态系结构, 导致以非移动型动物为主的古生代型生态系结构向以移动型动物为主的现代型生态系结构的重大转变[20]。但目前对造成第二幕如此严重的生物危机的原因还不明朗。中国地质大学(武汉)谢树成团队[21]通过对四川广元上寺剖面的生物标志化合物研究发现, 在第二幕碳同位素负偏和动物大灭绝时, 出现了大量真核微生物和原核微生物的繁盛、水体分层、海水硫化等现象, 这些现象正好又对应于古大气温度的升高以及随后的古海洋酸化[22, 23]。因此, 古温度升高可能导致古海洋分层, 一些微生物繁盛加剧了海洋缺氧并进一步发展为硫化。当时, 生物泵加强、异养呼吸作用加强, 但微型生物碳泵的作用可能减弱, 导致海洋出现酸化和碳同位素的负偏。以上记录说明:微型生物碳泵作用的减弱而生物泵的加强可能对2.52亿年前第二幕生态系统危机产生了极其重要的影响。
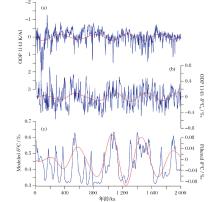
海洋沉积物记录显示上新世— 更新世的碳同位素(δ 13C)具有40万年周期性变化, 这一周期在中更新世拉长为50万年[24, 25]。δ 13C变化可以反映海洋碳储库的变化。前人研究将δ 13C这一长周期变化归因于陆源风化、海洋初级生产力、沉积雨比例等因素的变化[26]。近年来, 海洋溶解有机碳库的研究进展特别是微型生物碳泵概念的提出[27], 为解释古海洋变化提供了新的视角。“ 溶解有机碳” 假说成为解释大洋碳储库长周期变化的一种新假说[24]。同济大学马文涛等[28]利用改进的中等复杂程度箱式模型模拟了2 Ma以来的海水δ 13C变化。模型通过添加微生物环, 将碳循环分成快循环和慢循环2个环路。营养盐和生产力水平控制了碳循环的主要通路。模拟结果显示风化加强, 营养盐输入增加会导致惰性溶解有机碳(Recalcitrant Dissolved Organic Carbon, RDOC)浓度减小, 海水无机碳δ 13C变轻, 生物泵起更大作用; 而营养盐输入减弱时, RDOC浓度增加, δ 13C变重, 微型生物碳泵起等大作用。0~2 Ma地质时间段的模拟结果也显示, 40万~50万年周期变化受控于风化指标变化, 模拟的δ 13C相位和振幅都与地质记录一致(图3), 模拟结果可以支持“ 溶解有机碳” 假说。
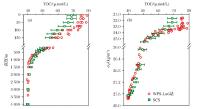
边缘海作为连接陆地和海洋间物质及能量交换的枢纽, 在全球海洋碳循环中扮演着重要的角色[29]。厦门大学戴民汉团队[30]研究指出, 在河流和上升流携带的大量营养盐的驱使下, 边缘海往往呈现较高的生产力, 其DOC的净生产量达到约0.38 Pg C/a, 占到新生产力的20%。我国南海北部陆架区冲淡水和上升流影响区域中DOC的净产率分别为(7.1± 7.0)和(11.5± 6.9)mmol/C(m2· d), 在冲淡水向外陆架输运的过程中, DOC的生产逐渐增加, 而在沿岸上升流的输运过程中, 同时存在DOC的生产和消耗, 并通过质量守恒计算得到冲淡水和上升流中大部分净生产的有机碳(60%~63%和59%), 主要通过颗粒有机物的沉降、溶解有机碳的扩散和水平运输等过程被移除出水体。吴凯等[31]对南海溶解有机碳浓度与西菲律宾海进行了比较, 结果发现:①在上层200 m, 南海DOC浓度要明显低于西菲律宾海; ②在中层水(1 000~1 500 m), 南海DOC要略高于西菲律宾海; ③在深层水(2 000 m~), 两者的DOC浓度一致(图4)。
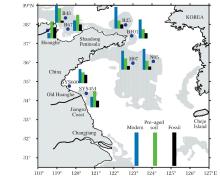
中国海洋大学赵美训团队研究表明, 中国边缘海是陆源和海源有机质的重要碳汇, 而海沉积有机质的来源和分布在过去600年受到快速气候变化及人类活动的影响。总有机质指标和生物标志物的结果显示, 我国边缘海表层沉积物中陆源有机质高值集中在近岸尤其是河口附近, 例如黄河口, 老黄河口及长江口; 海源有机质在陆架海盆处有高值, 受控于海洋生产力和沉积环境。总有机质和生物标志物14C含量测定则显示, 近岸表层沉积物有机质年龄高达7 000年。我国边缘海的有机碳埋藏量约13 Mt/a, 约占全球边缘海沉积物有机质埋藏量(约138 Mt/a)的10%, 其中陆源有机质的碳埋藏量(4.5 Mt/a)约占全球边缘海沉积物陆源有机质埋藏量的8%。若假定非现代有机质(陈化土壤有机质和古老有机质)主要是陆源物质, 则中国边缘海非现代有机质的埋藏量约为6 Mt/a, 约占我国边缘海总有机质埋藏量的46%, 与全球边缘海沉积物中陆源有机质的平均比例(44%)相当(图5)[32]。这些表明我国边缘海在全球海洋碳循环中具有重要地位, 是现代和千年尺度重要碳汇[33]。
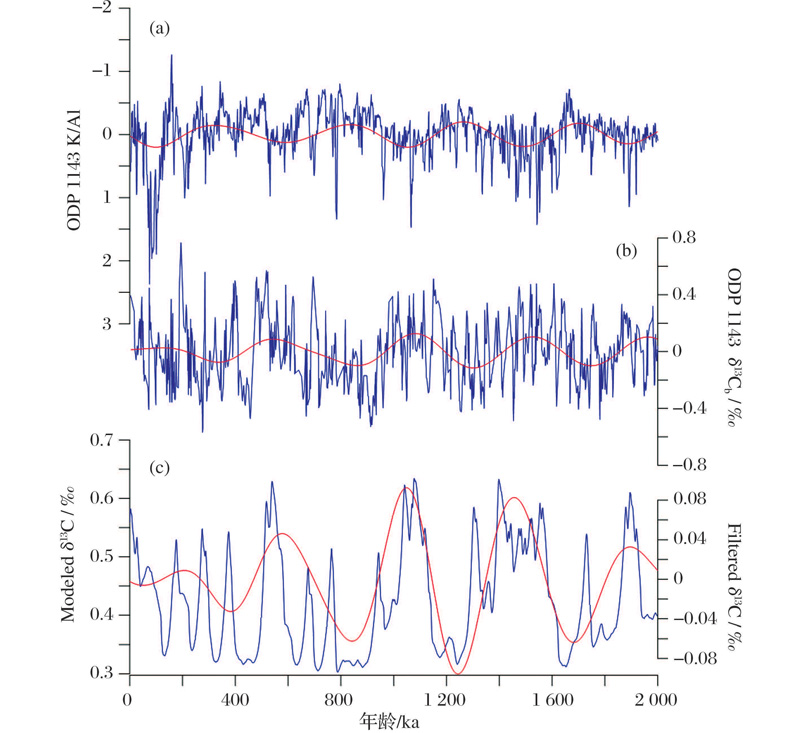 | 图3 0~2 Ma时间序列模拟 (a)ODP 1143站K/Al XRF扫描结果, 用于代表陆源风化和营养盐输入; (b)ODP 1143站底栖有孔虫δ 13C; (c)箱式模型模拟海水δ 13C结果; 红色曲线为40万年滤波结果[28]Fig.3 The simulation result of the last 2 Ma (a) Normalized K/Al record of ODP Site 1143, representing terrigenous weathering and nutrient input; (b) Benthic foraminifera δ 13C data from Site 1143; (c) Modeled global average δ 13C; Red lines are of 400-ka filtering[28] |
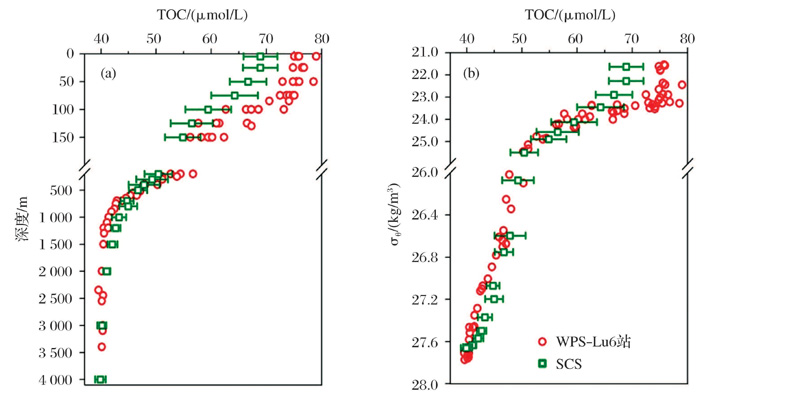 | 图4 南海与西菲律宾海DOC浓度的比较[31]Fig.4 Comparison of DOC concentrations between the South China Sea and West Philippine Sea[31] |
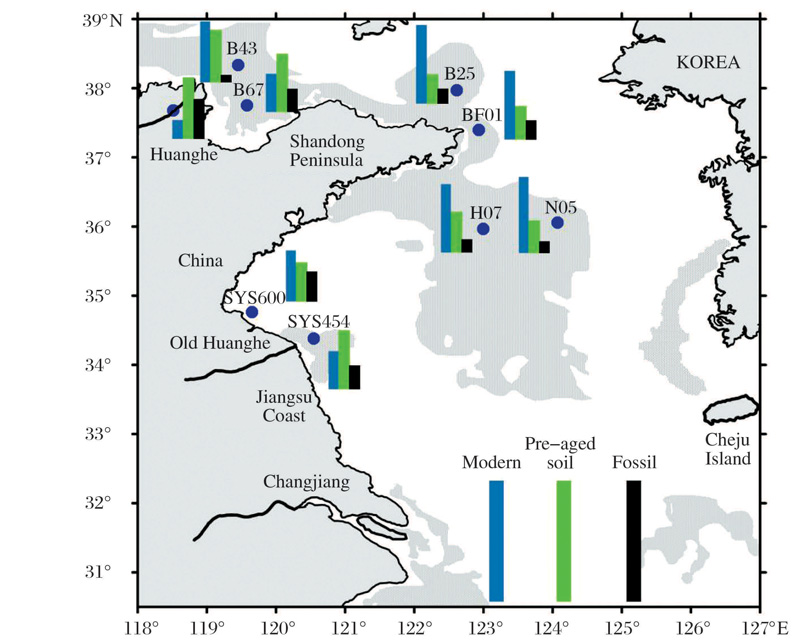 | 图5 δ 13C Bulk-δ 13C Compound=4~7时, 13C, 14C双同位素三元模型估算的渤、黄海表层沉积物中现代有机质(蓝色)、预陈化土壤有机质(绿色)和古老有机质(黑色)占总有机质的相对比例[32] 每一个色条代表一定的比例(色条越高代表百分比越高, 底部色条的高度代表的比例为100%)Fig.5 Coupled isotopic mass balance results for the fractional contributions of OC from modern (blue bar), pre-aged soil (green bar) and ancient fossil (black bar) sources in surface sediments of the BS-YS basin over a range of prescribed δ 13C bulk-δ 13C compound offsets (4~7)[32] The height of each bar is proportional to values (the higher bar correspond to the high percentage, assigning percentage of 100% as the height of bars shown at the bottom) |
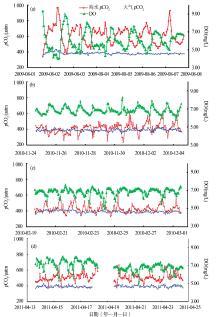
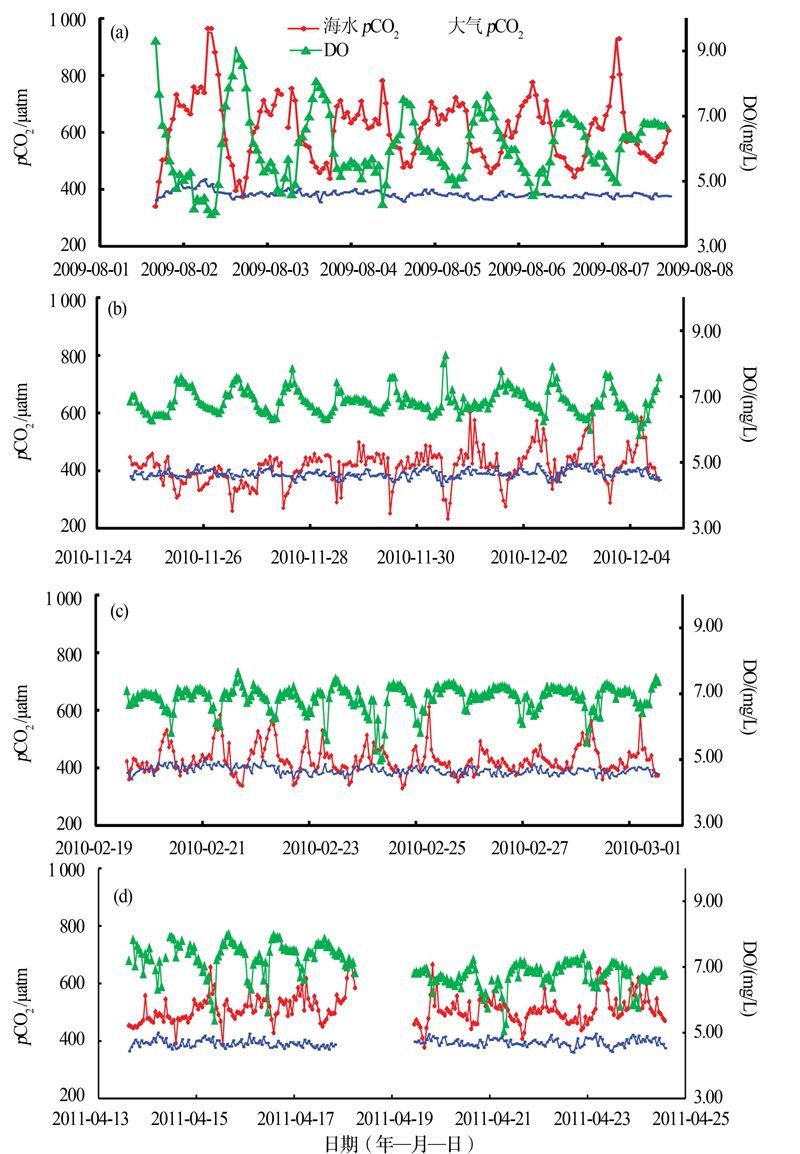
珊瑚礁区是海洋中最主要的碳酸盐生产区, 其碳酸盐年产率达到全球海洋的7%~15%。因为高生产力水平、高CaCO3生产量、快速变化的生态特征和复杂的理化环境, 珊瑚礁区的碳循环过程虽然一直倍受关注, 但关于现代珊瑚礁究竟是大气CO2的“ 源” 或“ 汇” 的认识却一直存在较大的争议[34, 35]。广西大学余克服团队[36]的研究指出, 南海珊瑚礁分布广泛, 通过对南海珊瑚礁的3种主要类型(岸礁、环礁、台礁)开展夏季珊瑚礁区海— 气pCO2的连续观测研究, 得出南海珊瑚礁夏季为大气CO2源的结论; 进一步针对海南鹿回头岸礁进行了不同季节的现场观测研究, 海— 气CO2通量的结果显示, 不同季节鹿回头岸礁均表现为大气CO2的源, 在生产力水平最高的夏季其“ 源” 的作用最强(图6)。
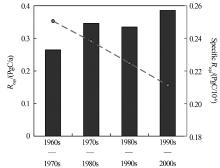
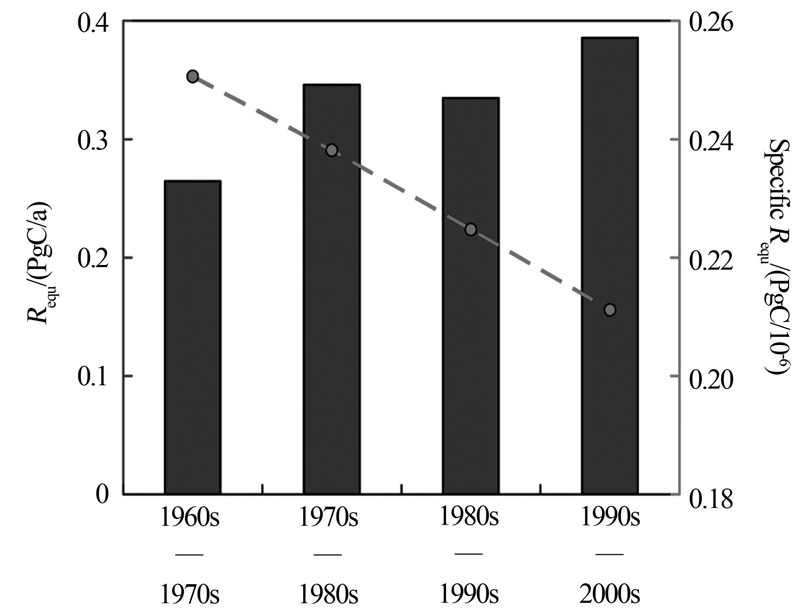
自工业革命以来, 人类因化石燃料使用和水泥生产所排放的CO2大约有41%被海洋吸收, 海洋对于缓解大气CO2的升高和全球变暖起到了重大作用。若海洋吸收人为CO2的能力趋于饱和, 则全球碳循环格局将发生重大改变, 进而威胁人类赖以生存的基本条件, 因此, 这一问题引起了科学家们的高度关注。山东大学海洋研究院翟惟东教授的报告指出, 海洋主要因温盐环流、生物活动以及上层海水的化学缓冲等机制从而大量吸收人为CO2, 根据覆盖全球大洋主要海区的表层碳酸盐体系数据集, 在大洋环流和碱度分布过去几十年稳定的假设下, 从而计算出大洋上层海水单纯由于海— 气平衡而积累人为CO2的速率。研究结果表明, 海表通过化学缓冲机制从大气移除人为CO2的效能在这50年中下降了16%(图7)[37]。该研究还显示, 当前人为CO2持续高强度排放造成海水化学缓冲物质迅速消耗, 海表碳酸盐体系吸收人为CO2的效能与工业革命前相比已损失31%, 预计到21世纪末, 海表通过化学缓冲机制调节大气CO2的效能将进一步下降55%, 接近枯竭[37]。
除了关注海洋表层对CO2的吸收, 厌氧环境下甲烷的产生也成为现代海洋碳循环过程研究的重要方面。梁前勇教授团队[38]研究表明, 南海北部陆坡天然气水合物区(东沙和神狐水合物钻探区)区海水甲烷含量为0~31.4 nmol/L, 平均含量为6.7 nmol/L, 高于全球平均海水甲烷含量, 表明南海海水甲烷含量本底值高于全球平均水平; 该区海水溶解甲烷含量及其分布特征不受区域海水水文特征、海水化学特征及季节等因素影响, 且表层海水— 大气甲烷交换并非单一的汇或者源的关系, 而是根据时间的不同, 海水— 大气甲烷交换存在汇源转换; 综合调查结果表明, 南海北部陆坡天然气水合物区甲烷渗漏对海水、大气甲烷含量没有明显影响, 且水合物钻探对区域环境没有明显的影响。
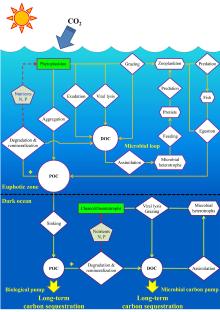
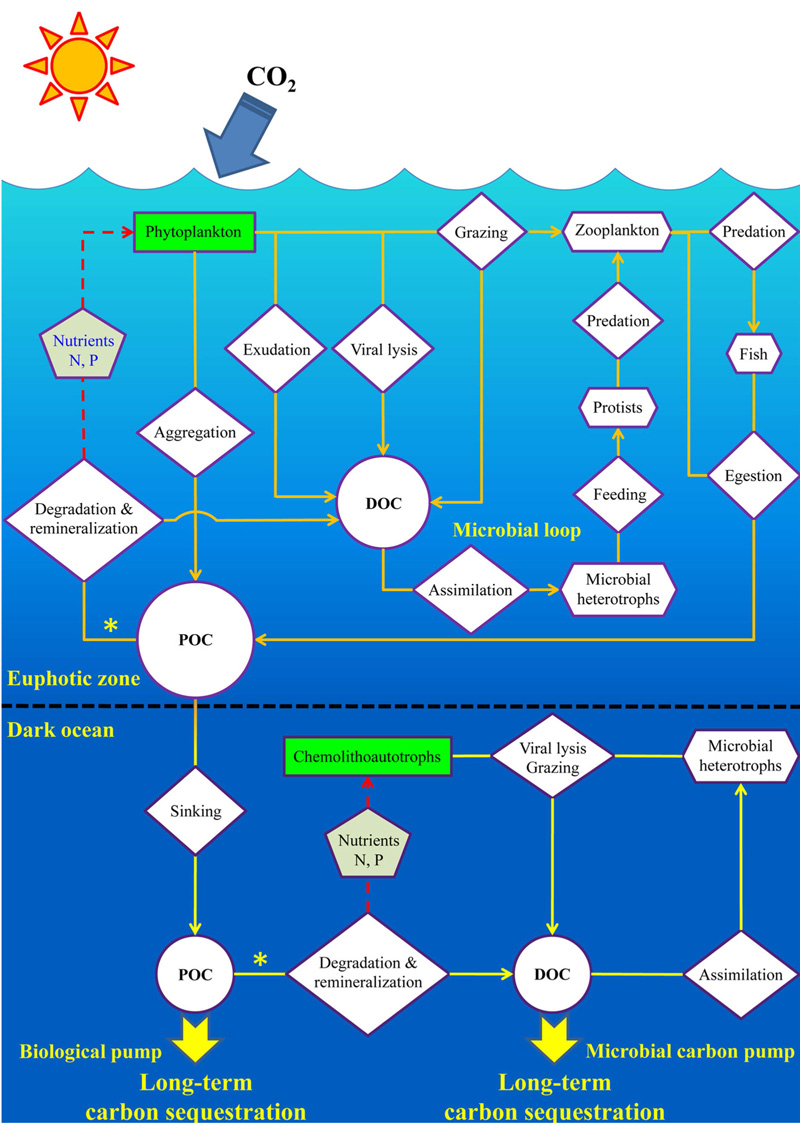
海洋对大气CO2的固定和储藏, 主要是通过一系列生物介导的生态过程来实现。此前人们熟知的海洋生物泵导致的颗粒有机碳向深海的输出十分有限, 到达海底埋葬的有机碳量大约只有海洋初级生产力的0.1%, 绝大多数POC在沉降途中被降解呼吸转化成CO2。全球大洋深海中稳定存在着大约40 μ mol/L的DOC, 这部分DOC一般被认为是惰性RDOC。MCP概念的提出(图8)[39, 40, 27]对海洋RDOC碳汇的成因给出了一个合理的解释。MCP理论框架的建立[27]提供了若干可验证的科学假说, 成为研究热点[41]。近期, 焦念志教授与国际同行共同撰写的有关MCP的BG专刊, 突破了经典理论的束缚、阐释了RDOC的时空属性; 建立了BP-MCP的链接、搭建了古今结合的桥梁; 提出了全球变暖背景下MCP的相对重要性增加的可验证科学假说[42]。长期以来, 有关RDOC碳库的形成有2种假说:“ 稀释假说” 和“ 生物惰性假说” , 焦念志团队[43]发表在Science上的评述文章阐释了深海惰性有机碳库假说, 用“ 微型生物碳泵” 理论框架将2种假说实现了有机链接。
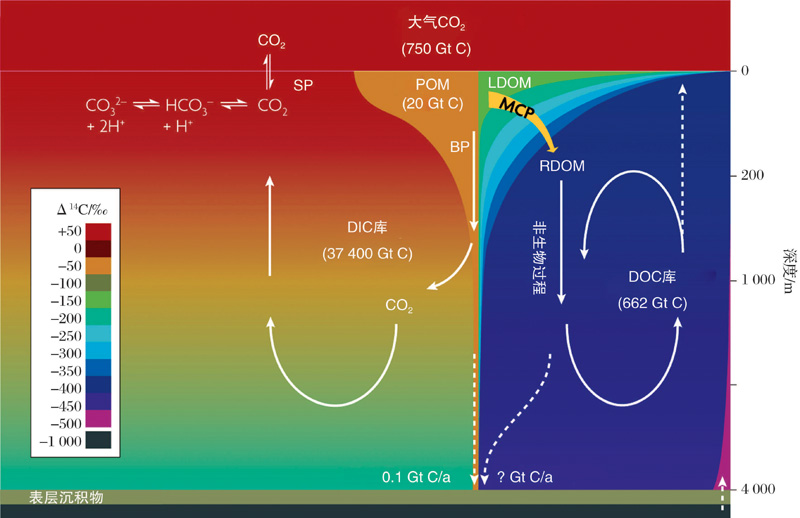 | 图8 海洋碳汇的生物学机制(生物泵、微型生物碳泵、碳酸盐泵)及其环境效应[27]Fig.8 Biologically-driven ocean carbon pumps (BP, MCP, CCP) and their environmental consequences[27] |
然而, MCP过程非常复杂, 是典型的多学科交叉科学命题。就微型生物本身而言, 涉及了自养、异养、原核、真核各个类群、功能各异。无论是DOC还是POC, 其生物学、生态学及生物地球化学过程和机理差异很大, 但二者又是紧密联系和相互影响的。目前上述这些过程尚未完全明了, 若干自然现象及其背后的机理有待回答。
海洋颗粒物及其他固体表面附着微生物往往与海水中自由生活的微生物有着不同的种类组成和功能过程, 它们在海洋营养盐再生、碳氮磷硫铁等生源要素循环、食物网物质、能量和遗传信息流动、环境污染物降解中发挥着重要作用。附着微生物不但通过呼吸作用影响海洋POM降解, 从而影响海洋POM再矿化深度、海洋生物泵储碳作用、海洋对大气CO2浓度及全球气候变化的调节作用, 而且, 附着微生物在降解POM的同时, 加速营养盐再生, 从而促进海洋真光层的光合固碳作用及弱光和无光层的微生物化能自养固碳作用(图9), 对全球气候变化和海洋环境变化具有重要影响[44]。厦门大学的党宏月教授[45]系统分析了海洋附着微生物在不同水层的生态效应, 深入解析了浸海固体表面和海洋颗粒物附着微生物的遗传、生化生理、生物地球化学及代谢组学等特征和规律, 分析了海洋附着微生物对全球变化(如升温、酸化、缺氧等)的响应和影响, 为海洋微生物附着及相关的全球变化等研究提供系统的理论基础和科学指导。
海洋是一个巨大DOC碳库, 其中DOC本身的变化大部分是被异养细菌改造的, 异养细菌转变DOC的途径和速率决定了最终转变成CO2 或者 RDOC的量。存在上千年的溶解有机碳组分随着被生物周转利用的速率、生物可利用性和生物化学特征都是不一样的。微生物过程可以改变DOC库的组成, 反过来, DOC的可利用性也塑造了微型生物的多样性和群落结构。例如Rosebacter进化分支和SAR11进化分支分别主导了富营养和寡营养的海区。每个进化分枝都有自己独特的碳源利用策略, 这样不同微型生物的响应又作用于海洋DOC库的组成[46]。
海洋环境中, 异养细菌与浮游植物以及海洋病毒的互作关系错综复杂, 包含共生、寄生、竞争、抑制或致死、信号通讯、基因转移等。研究不同海洋微型生物的互作对更深入了解异养细菌驱动的DOC转变过程有重要的科学意义。中国科学院青岛生物能源与过程研究所的梁彦韬[47]利用流式细胞技术对多个海域海洋超微型浮游生物丰度的大尺度分布规律进行了研究, 研究发现海洋上层浮游病毒、异养细菌、聚球藻、超微型真核藻类的在高叶绿素浓度的近海和大洋上升流区域丰度较高; 近海聚球藻春季出现低值, 推测可能受到异养细菌生长的竞争抑制; 海洋上层 (0~200 m) 和中层 (200~1 000 m)浮游病毒丰度的变化主要受到自养、异养宿主丰度的影响; 深海(> 1 000 m)浮游病毒丰度变化不大, 其变化主要与非生物环境参数显著相关。中国科学院青岛生物能源与过程研究所的张永雨[48]在对2株聚球藻的藻菌互作体系中发现高密度的聚球藻在自然海水菌群的影响下, 可快速聚集沉淀, 并引起藻类死亡裂解, 并呈现出一定的宿主特异性。同时发现聚球藻生长或释放过程中释放的DOC活性较高, 能够快速被细菌等吸收利用, 从而实现碳元素从浮游植物到异养细菌生物量的传递。证实浮游植物与异养细菌相互关系在海洋微食物环碳传递或分馏过程中起着重要的调控作用。暨南大学杨宇峰教授的团队[49]研究发现, 在近海环境中, 大型海藻栽培, 在增加碳汇、降低N, P营养负荷、提高水体溶氧、控制有害藻华、维持海水养殖生态系统健康方面具有很重要的作用。
此外, 过去通常认为海洋古菌仅在极端环境中生存, 近些年的宏基因测序分析表明, 从表层海水到深层大洋均有大量分布着不同类群的古菌。海洋古菌在不同水层的分布与生态作用是海洋微型生物驱动DOC转化过程不可缺失的有机组成。海洋浮游古菌主要分为自养类群MGI 和异养类群MGII。同济大学海洋地质国家重点实验室的谢伟等[50]通过对珠江口区域不同盐度站点的连续1年的观测发现, 在半咸水区域存在MGII 持续勃发的现象。进一步对影响其勃发机制的探究发现, MGII16S rRNA 的量与该区域藻类的23S rDNA 量存在很好的正相关, 表明其勃发与该区域营样盐输入带来的藻类生长有密切的关系。进一步研究发现, 在以淡水种蓝细菌主导的环境下, MGII 与藻类的相关系数与指数均较小, 而硅藻、绿藻、隐藻、蓝藻混合分布的环境下, MGII与藻类的相关系数和指数均最高, 表明MGII 对不同藻类基质利用的差异性。
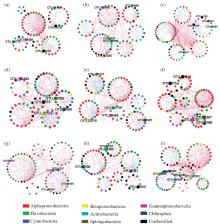
日益严重的海洋酸化现象正成为一个严峻的全球生态问题。海洋生物均受到海洋酸化的明显影响。然而, 作为海洋生地化循环过程中重要的组成部分, 海洋微生物群落如何受到海洋酸化的影响却并没有一个明确的定论。基于pMEN分析方法研究了不同pCO2条件下微生物间的相互联系, 研究发现微生物群落通过其相互联系, 在群落水平来抵抗酸化的影响, 这一结果指出微生物在未来海洋环境中更加重要的生地化作用, 并为我们进一步研究海洋酸化对于微生物群落的影响提供了新的视角(图10)[51]。
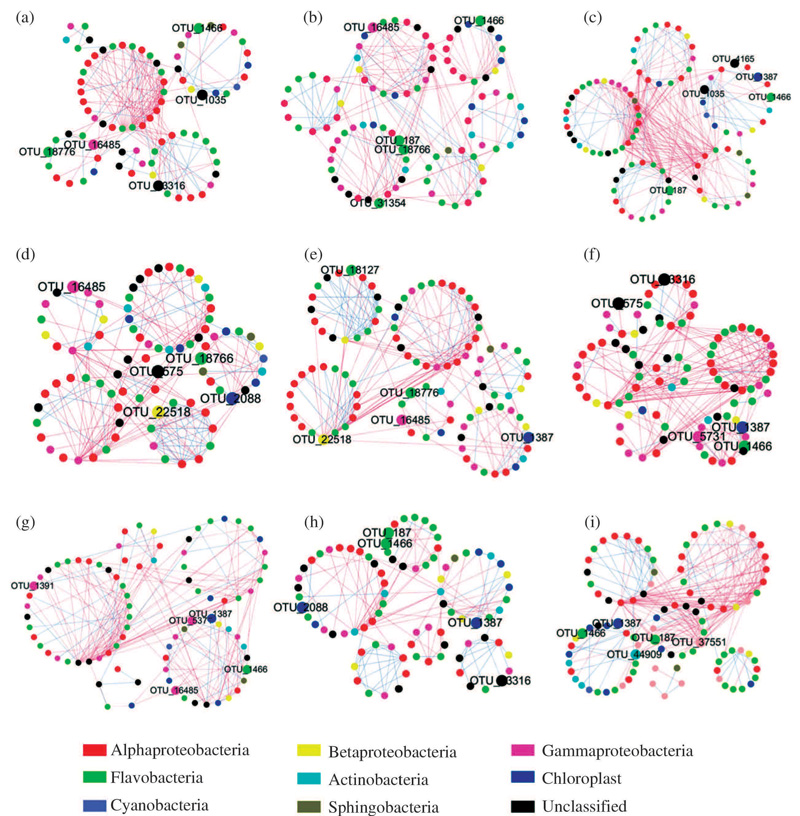 | 图10 在175(a), 180 (b), 250 (c), 340 (d), 425 (e), 600 (f), 675 (g), 860 (h), and 1 085 (i) μ atm pCO2下微生物网络相互关系 展示了每个网络中的5~7个模块, 每个点代表一个OTU, 点的颜色代表不同的门, 2个点之间的连线代表2个物种间的联系, 蓝色为正相关关系, 暗示为共生关系, 红色为负相关关系, 暗示为竞争关系; 丰度最高的5个OTU用较大的点表示。点围成的圈表示一个模块, 这些点在模块内的联系比模块外更加紧密[51]Fig.10 Bacterioplankton network interactions under 175 (a), 180 (b), 250 (c), 340 (d), 425 (e), 600 (f), 675 (g), 860 (h), and 1 085 (i) μ atm pCO2 Five to seven modules were formed under different pCO2 concentrations. Each node represents an OTU, which signified a species. Node colours indicate different phyla. Each line connects two OTUs. A blue line indicates a positive interaction between two individual nodes suggesting a mutualism or cooperation, while a red line indicates a negative interaction suggesting predation or competition. Top five highest-abundance OTUs are indicated by bigger dots and marked with OTU identification numbers. The cycle composed of several nodes is a module in the pMEN, which is more correlated in a module than between other modules. Only the module containing more than five OTUs are displayed |
如本文开篇指出, 海洋碳汇是一个宏大的科学命题。本专题系统梳理了从冲淡水到上升流、从上层水二氧化碳的吸收到厌氧环境甲烷的产生、从初级生产力到新生产力、从溶解有机碳到颗粒有机碳、从沉积有机质来源及碳汇意义到南海珊瑚礁碳循环及其源汇争议、从现在海区的源汇评估到未来海洋的碳汇预测等方面的研究进展, 从空间尺度上诠释现代海洋碳循环研究, 从河口到陆架, 从浅海到深海, 从水体到沉积物, 进一步探讨现代海洋(边缘海、珊瑚礁区等)CO2源与汇问题, 了解人为CO2在当前全球变化背景下的归宿。本专题研讨可望成为该领域研讨的一个良好开端, 本文旨在以飨读者, 希望本领域科技工作者在此基础上通过学科交叉与融合进一步深入探讨海洋碳汇及其可能产生的生态环境效应、对气候变化的影响、对人类活动的响应认识等。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|