{kind=link}
{kind=link}
{kind=link}
{kind=link}
近岸底层水体低氧沉积记录研究进展
[吴伊婧1  , 范代读
, 范代读1, 2*, * , 印萍3 , 胡虞杨1 ]
, 范代读, 印萍|
|


作者简介:吴伊婧(1990-),女,浙江嘉兴人,博士研究生,主要从事海洋地质学研究.E-mail:walada@qq.com*通信作者:范代读(1972-),男,福建大田人,教授,主要从事海洋沉积学研究.E-mail:ddfan@tongji.edu.cn
长时间尺度的沉积记录可以提升对近岸低氧形成机制的理解,从而为预报、预测和发展减缓低氧措施等提供必要的依据。综述了近岸低氧发育与演化历史的主要研究进展,尤其侧重于沉积记录的研究。首先按水体受限程度将近岸低氧发育环境划分为半封闭型海盆/海湾和开阔陆架海2类,分别探讨了二者低氧发育的主要特征。其次分析总结了低氧沉积记录的替代性指标,包括对水体氧化还原环境具有较好指示意义的生物学、矿物学和地球化学指标,着重分析了各类替代性指标的适用性。最后对长江口外海域底层水体低氧发生机制与沉积记录的研究现状作了回顾与分析。目前长时间尺度的低氧沉积记录研究仍然较少,鉴于长江口外海域低氧的动态发育特征,提出了多钻孔、多参数结合的研究方案。
First author:Wu Yijing (1990- ), female, Jiaxing City, Zhejiang Province, Ph. D student. Research areas include marine geology.E-mail:walada@qq.com
Corresponding author:Fan Daidu (1972- ), male, Datian City, Fujian Province, Professor. Research areas include marine sedimentology.E-mail:ddfan@tongji.edu.cn
Sedimentary records are potential to provide long-term evidence for better understanding the development mechanism of coastal hypoxia, shedding some light on the forecast, prediction and controlling-measure development to mitigate hypoxia. Therefore, recent research advances in the formation mechanism and evolution history of coastal hypoxia were briefly reviewed, specially with focus on sedimentary records and proxy methods. First, marine environments with hypoxia were classified into semi-enclosed marginal sea/gulf and open shelf sea based on the degree of bottom-water circulation and ventilation, and main characteristics for the hypoxic development were discussed respectively. Secondly, the methodology was reviewed in the efficiency by using different proxies to reconstruct hypoxia development history from sediment cores, including redox indicators of sedimentology, biology, mineralogy and geochemistry. Ultimately, recent research advance in hypoxic development mechanism and evolution history off the Changjiang Estuary were summarized. It is worth noting that long-term evolution history has been less studied from long cores. It is therefore suggested that a synthetic methodology involving multi-core comparison with different-proxy interpretation should be employed to study the development history of seasonal hypoxia off the Changjiang Estuary.
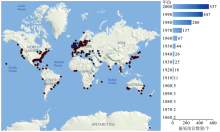
水体低氧的发生受区域内物理、化学及生物过程的共同控制, 从湖泊、近岸到大洋的广袤水域都有孕育低氧的可能性, 地质历史时期甚至发生过几次全球性的大洋缺氧事件[1~4]。近岸低氧(Coastal hypoxia)有别于湖泊低氧和大洋最低含氧带(Oxygen Minimum Zones, OMZs), 其生消过程明显受控于陆海相互作用。自19世纪50年代首次观测记录到近岸低氧现象以来, 被观测到的低氧发生区域不断增加, 且从持续时间、分布范围或氧亏损程度上都有不断加剧的趋势, 尤以近50年为甚(图1)。这一定程度上归咎于人类活动增加导致近岸海洋环境恶化, 严重影响了区域生态健康, 可能造成大面积海洋生物资源严重衰退的“ 死亡地带(Dead zone)” [5]。值得注意的是, 报导出现近期近岸低氧发生频次、强度和面积都在显著增加, 这与现场观测或监测方法与能力的提升、公共关注程度的增加等也有一定关系。当前有诸多国际科学研究计划, 如LOICZ-II(Land-ocean Interactions in the Coastal Zone), IMBER(Integrated Marine Biogeochemistry and Ecosystem Research)和GEOHAB(Global Ecology and Oceanography of Harmful algal blooms)等, 以及专注于大河河口及其河控陆缘海的研究计划RiOMars(River-dominated Ocean Margins)和LDE(Large-river Delta-front Estuaries)等都以此作为核心研究内容之一。
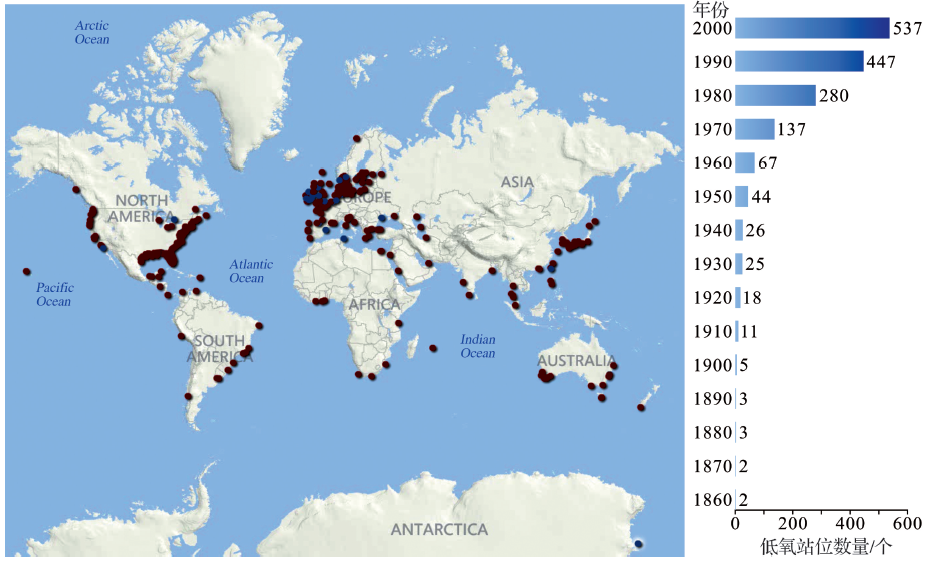 | 图1 19世纪50年代以来全球主要近岸低氧区域分布与数量变化[6] 暗红点表示低氧区域、深蓝点表示低氧程度得到改善的区域Fig.1 World-wide distribution and development of coastal hypoxia and variation in the observed numbers since the 1850s[6] Dark red and blue denoting hypoxic areas and some hypoxia-alleviated areas |
水体溶解氧(Dissolved Oxygen, DO)含量的改变会导致区域内生物量减少、群落结构及模式发生改变。首先表现在食物链顶端的游泳生物逃逸或死亡, 威胁渔业资源[7]; 严重时海水中的部分微生物在缺氧环境下会进行无氧呼吸, 产生有毒H2S气体[8]; 另外, 由于表层沉积物原先的氧化环境被改变, 部分有毒有害化学物质可能再次活化, 造成二次污染[9]; 低氧还会改变早期成岩作用及沉积物— 水界面的物质循环[10], 是温室气体N2O的重要来源[11], 长时间尺度上对海洋酸化有加强作用[12], 对全球气候的影响也亟待进一步研究。
目前基于长期、系统性观测数据所建立的模型使得对低氧水体体积和面积的短期预报成为可能[13, 14], 并发现采用一定控制措施之后, 部分区域的低氧情况出现好转[15, 16]。但在大部分区域, 历史资料的匮乏仍遏制着长期预报能力的发展。通过沉积记录恢复低氧的演化历史可以为模拟、预测、预防甚至采用措施控制低氧发生与发育等提供必要的支撑, 同时也是重建受低氧影响前研究区域生态、地球化学背景的唯一手段[17, 18]。
水体氧化还原程度(表1)的替代性指标是恢复低氧沉积记录不可或缺的媒介, 其效果取决于区域低氧程度与时空分布等特征。本文在分析不同近岸环境低氧主要特征的基础上, 总结了部分对水体氧化还原环境具有直接指示作用的替代性指标, 及其影响因素和适用环境。并以长江口近岸底层水体低氧为例, 综述了该区域低氧特征与沉积记录的研究现状, 对替代指标和研究方案提出了改进设想, 以期更好地揭示长江口低氧形成与演化历史。
近岸底层水体低氧现象分布广泛, 可发生于各类近岸海域, 不论是较为封闭的海盆、澙湖、峡湾, 或是开放的河口、陆架等环境[20]。导致底层水体氧亏损的原因众多, 富营养化[2]和水体层化[17]是其中的2种重要机制。实际研究发现, DO消耗— 补给平衡受制于各种物理与生物化学过程, 使得不同区域低氧的程度、分布范围以及持续时间出现明显差异, 据此可大致分为半封闭型海盆/海湾低氧和开放型陆架低氧2类。
大洋最低含氧带水体的含氧量往往受控于水体通风性[3], 半封闭型海盆/海湾区域底层水DO交换同样受到限制, 具孕育低氧的自然条件。
波罗的海深部海盆中发育的永久性低氧因其面积大、程度重而广为人知。沉积记录研究表明, 早在8 500~7 800 cal a BP盐跃层开始稳定形成的同时, 波罗的海深水区就间歇性地被持久性低氧所盘踞, 其发展规律与气候波动相联系[21]。但在过去1个世纪中叠加了人类活动的影响, 使低氧的程度加重、范围扩展数倍[22~24]。黑海是最大的现代永久性低氧海盆, 低氧水体覆盖海盆面积达75%, 20世纪70年代以来其溶氧跃层的深度变浅近10 m, 可能与气候变化和营养盐输入增加等有关[25, 26]。委内瑞拉Cariaco海盆低氧形成历史也十分悠久, 可以追溯至末次冰消期, 由于水体交换受限和高生产力, 海盆深部300 m密度跃层之下也终年存在严重的低氧[27]。
海湾相对于海盆区域水体封闭性较低, 低氧的程度与持续性也较弱。切萨匹克湾发育季节性低氧, 沉积记录研究表明自17世纪初欧洲殖民开始以来, 湾内沉积速率、水体富营养化呈上升趋势, 与此同时低氧现象也在加剧[28]。1985年以来较为连续的低氧观测数据显示, 对比历年夏初记录, 切萨匹克湾低氧呈增强态势, 而夏末低氧记录则相反, 这是因为不同阶段低氧发育的主导因素不同, 水体层化增强而营养盐输入受限共同导致了这一变化[29]。
峡湾中水体交换同样受到较大限制, 部分发育永久性低氧, 如Framvaren峡湾, 但大部分峡湾因存在周期性深部水体更换而只发育季节性低氧[2]。
半封闭型海盆/海湾区域低氧的共同特征是氧亏损程度严重, 能够达到无氧甚至硫化的条件, 且多为自然形成。
相对于封闭海域发生的低氧, 开放型陆架低氧的控制机制更为复杂[30, 31]。根据营养盐和有机质来源, 开放型陆架低氧常发生于有径流输入或(和)上升流发育的区域。
径流冲淡水为河口陆架海区注入大量陆源营养盐和有机质, 同时与陆架环流、潮汐、风、海表温度等物理机制共同作用导致水体层化[32]。一般海洋能量低(弱潮、流速低、风浪小), 而径流量和水体温度梯度大的河控型陆架更易形成持续层化, 从而引发低氧, 并能更好地保持低氧水团的稳定性[1, 33, 34]。如墨西哥湾北部德克萨斯— 路易斯安那沿岸, 径流注入的淡水与物质在夏季风向和环流作用下大部分被限制于陆架上, 与强大的温度梯度共同作用形成大范围低氧[35, 36]。故该区域低氧表现明显的季节性, 在7~8月达到最强, 年均持续时间基本在5个月以上, DO浓度接近无氧条件[37, 38]。
上升流可将底部高营养水体输送至表层, 促进有机质产生。在新泽西内陆架, 夏季百慕大高压和西南风的驱使下上升流发育, 对初级生产力的促进远超陆源营养盐的作用, 沉降到底床的有机质分解导致低氧发生[39]。此外, 上升流还可以增加陆架区域与OMZs的水体交换。OMZs一般分布在水深400~1 200 m的陆坡区域, 处于永久性温跃层底界面附近[3], 但也有统计发现, 非El Niñ o年智利岸外OMZs水体可以入侵至距岸线仅25 m的浅海区域[40]; 还有报道称俄勒冈沿岸内陆架受OMZs水体上涌形成严重的低氧现象[41, 42]。在这些区域, 低氧的发生可能并非受水体层化或有机质降解主导, 而是直接与含氧量低下的OMZs涌升入侵至陆架区有关。气候变暖可导致OMZs扩张、变浅和涌升增强, 这一现象正在被观测记录并呈发展趋势[43, 44]。
每一次水体氧化还原条件的改变都会在沉积物中留下丰富的沉积学、生物学、矿物学和地球化学标志, 合适的替代指标对沉积环境的判别具有良好指示意义。由于低氧的形成与水体层化及富营养化关系紧密, 故对部分指标进行应用解释时需辨明其具体含义。如纹理发育的沉积层除了反映生物扰动较弱之外, 也与高沉积速率、强水体层化不无关系[18, 21]; 而有机元素、色素、甾醇及生物硅等指标则通过指示水体生产力变化而间接反映低氧[45, 46]。这类指标只能作为重建低氧历史的参考而非直接证据。此外, 即便是对低氧有直接指示意义的指标, 其有效性也不尽相同。且不论多解性或早期成岩作用的影响, 各指标本身对低氧的敏感度亦存在差异。所以因地制宜选择有效的、合适的, 尤其是对低氧有直接指示意义的指标就显得十分重要。
微体化石是沉积环境中微体生物种群的最直接记录。DO作为一种重要的生态因子, 对有孔虫、介形虫、翼足类等微体生物的丰度或优势种属均有显著影响[47~49]。底栖有孔虫由于有部分种属对水体氧亏损耐受度较强和良好保存性而被广泛运用于低氧沉积记录的研究中。当水体氧含量下降时, 底栖有孔虫个体会变小, 耐低氧种属的比例增高同时多样性随之降低, 不过由于捕食者与竞争者的减少, 其丰度在一定氧亏损范围内反而会增多[48]。
由于单一有孔虫种属的丰度易受种间竞争关系及温度、盐度、底质类型等生态因子的复杂影响, 故低氧研究常采用耐低氧有孔虫组合(Low-Oxygen Foraminiferal Assemblages, LOFA)[50]。在某些研究较为成熟的区域, 已发展出利用若干具代表性的种属组合所建立的低氧判别指标。Gupta等[51]在墨西哥湾北部德克萨斯— 路易斯安那陆架低氧沉积记录的研究中, 通过对2处表层沉积物(水深9和21 m)中有孔虫种属的分析发现, 夏季低氧高峰过后, 2处均存在大量Ammonia parkinsoniana活体和Elphidium excavatum空壳, 由此提出了A-E指数:100× NA/(NA+NE), 其中NA和NE分别代表A. parkinsoniana和E. excavatum的数目, 并以此分析了该区域200年来的低氧发育情况。由于A-E指数仅适用于水深30 m以浅的区域[52], Osterman[53]进一步统计分析了采自墨西哥湾北部10~1 020 m 水深的74个表层沉积物样品中的93个底栖有孔虫种属, 从而定义了PEB指数(Protononionatlanticum, Epistominellavitrea和Buliminellamorgani所占百分比之和), 以此揭示了该区域1 000多年来周期性低氧发育的规律, 认为与自然条件变化密切相关, 但近几十年在人类活动影响下发生显著恶化[35, 54]。
无论是A-E指数还是PEB指数都并非适用于所有区域, 因为其中部分种属受生物地理分布的局限性影响显著[18, 55]。同时, 有机质供应、温度和水深变化对底栖有孔虫优势种属的影响也不可小觑[18]。因此在利用LOFA作为低氧判别指标时, 应根据区域具体情况进行相应调整。另外, 不同区域LOFA种属的差异会使历史氧亏损程度对比存在困难, 故进行相关分析时需要参考其他指标。
某些藻类、细菌等生物死亡埋藏后罕有实体化石记录, 但是生物体中蕴含的特定有机化合物可以保存并指示母体类型, 从而反映历史沉积环境信息。水柱中磷脂脂肪酸(Phospholipid Fatty Acid, PLFA) 和细菌藿烷多醇(Bacteriohopanepolyol, BHP)等生物标志化合物分析表明, 甲烷氧化细菌或/和硫酸盐还原细菌在黑海、波罗的海、Cariaco海盆深部的溶氧跃层之下存在[56~58]。在此基础上, Berndmeyer等[59]分析了波罗的海深部哥特兰海盆表层沉积物的生标特征, 发现低氧条件下沉积物中BHPs保存良好并与水柱中的组成特征相似, 认为沉积物BHPs主要来源于底层低氧水体, 因此可通过沉积记录中特征BHPs的研究重建低氧历史。据此分析显示, 末次冰消期以来波罗的海深部区域逐渐由充氧淡水湖泊环境向低氧现代咸水海盆环境转化[60]。
另一类生物标志化合物基于不同氧化还原条件下有机质降解产物的差异以指示沉积环境。姥鲛烷/植烷(Pristane/Phytame, Pr/Ph)比值是古氧化还原环境研究中的常用指标, 两者的原始母质主要是植醇, 区别在于前者通过氧化途径形成, 而后者倾向于还原途径, 故相对氧亏损的环境下Pr/Ph值较低[61]。然而近现代沉积物Pr/Ph值受有机质来源和成熟度影响明显[62, 63]。 Bogus等[64]对阿拉伯海表层沉积物研究显示, 姥鲛烷、植烷及植醇三者, 或两两组合而成的指标与DO浓度无明显关系, 指出这可能是由于植醇降解并非姥鲛烷单一来源所致。其他来源包括生物圈中广泛分布的维生素E, MTTCs(methyltrimethyltridecylchromans)都使Pr/Ph值的可靠性大大降低[65]。不过培养实验显示, 植醇的其他降解产物也表现出对沉积环境氧化还原条件的敏感性, 其应用潜力有待进一步探索[66, 67]。
值得注意的是, 相对氧化的环境不利于部分生物标志化合物的保存, 所以应用区域基本局限于发育严重永久低氧的海盆。
Berner[68]总结了对于水体氧化还原条件较具指示性的部分自生矿物, 其中铁硫化物和海绿石已经在近岸低氧沉积记录研究中有所运用。
当水体氧亏损较严重时, 硫酸盐成为细菌呼吸作用的主要电子受体, 反应产生的部分H2S与金属元素结合, 铁硫化物便是其中一种比较主要的产物类型, 黄铁矿更是占了大部分。由于有机质和硫酸盐在近岸海域的广泛分布, 该反应普遍发生, 但形成矿物的组分、结构等特征有所不同。Berner[69]提出, 正常沉积物中有机碳与黄铁矿含量比值(C/S)约为2.8, 严重低氧时, 由于来自水柱的黄铁矿输入, C/S会降低, 理论上能够反映氧亏损程度。但实际上C/S的应用十分局限, 而FeS与黄铁矿中S之比(AVS/CRS)和黄铁矿与总活性铁中Fe之比(DOP)作为更有效的指标被先后提出[70~72]。在对切萨匹克湾低氧的研究中, Cooper等[28]利用DOP重建了近2 000年的低氧历史, 发现20世纪40年代起底层水的氧化还原情况发生了变化; 此后, Zimmerman等[73]发现了同层位AVS/CRS突然增大的现象。另外, 黄铁矿的颗粒大小与形态等特征也能为水体氧化还原环境历史变化提供证据[74, 75]。
海绿石是一种绿色黏土矿物, 一般认为形成于次氧化至弱还原条件、弱碱性的浅海环境中, 其晶格中同时含有二价和三价铁, 因此对周期性的底层水体低氧具指示作用, 也较黄铁矿更易保存[9, 76]。自生海绿石的形成很大程度上受沉积速率的制约, 因为过快的沉积物供应和埋藏作用会切断构成物质的供给, 阻碍粪球粒和生物碎屑的海绿石化[77]。近岸低氧通常发生于陆源物质供应充足、沉积速率较高的区域, 似乎与海绿石形成的有利条件相悖。然而Nelsen等[78]发现密西西比河口短柱样中的海绿石含量变化与其他低氧指标一致, 提出水体氧化还原环境的频繁变化以及人类活动导致的物质输入增加会加速海绿石化作用, 促使其快速生长。此后的研究也进一步证明了海绿石作为低氧指标在该区域的有效性[45]。不过在东海, 海绿石主要富集分布于沉积速率较慢的残留砂沉积区[79], 与传统低氧区域相距甚远, 未必是合适的替代指标。
氧化还原敏感性元素(Redox Sensitive Elements, RSE)是具多种价态的过渡金属元素, 因在氧化和还原条件水体中的溶解度明显不同, 而倾向富集于还原环境中, 但不同元素的富集与稳定保存条件迥异, 故不同区域/氧亏损程度/低氧类型的有效指标也不尽相同。
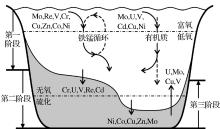
RSE的自生富集过程大致可分为清除、固定与沉积后作用3个阶段(图2)。第一阶段, 表层氧化环境中溶解的离子态RSE吸附于有机质、铁锰氧化物等颗粒表面被输运到底部, 由于载体的分解或再度释放到上覆水体或沉积物孔隙水中[80~83]。第二阶段, RSE在还原条件下(水体或沉积物中)各自以较为稳定的形态沉积下来, 如Ni, Cu, Cd等易形成固态硫化物颗粒, U和V等易成为腐殖酸的有机金属配合体, Re和Mo等则会以有机络合物或无机簇合物的形式而埋藏[84~86]。这一阶段的发生很大程度上受控于水体的还原程度, 因为不同RSE开始自生富集的临界点不一样。比如还原条件下Mo的沉积主要依赖于硫化物的出现, 其富集大多发生在硫化环境中, 而Re和Cd在无可检出O2或H2S的无氧条件下就能自生富集[80, 87, 88]。第三阶段, 当上覆水体氧化还原条件周期性或突发性的改变时, RSE多少都会受再氧化作用而迁移甚至回到水体中, U所受影响尤为明显, Mo和Cu等较小, Cd几乎不受影响[84, 89]。此外, 人类活动的污染会影响Cu, Re和Pb等部分RSE的分布[81, 82]; 一些较为局限的海盆或峡湾中, RSE的供应限制也会导致沉积记录中RSE/TOC比值的波动[90]。
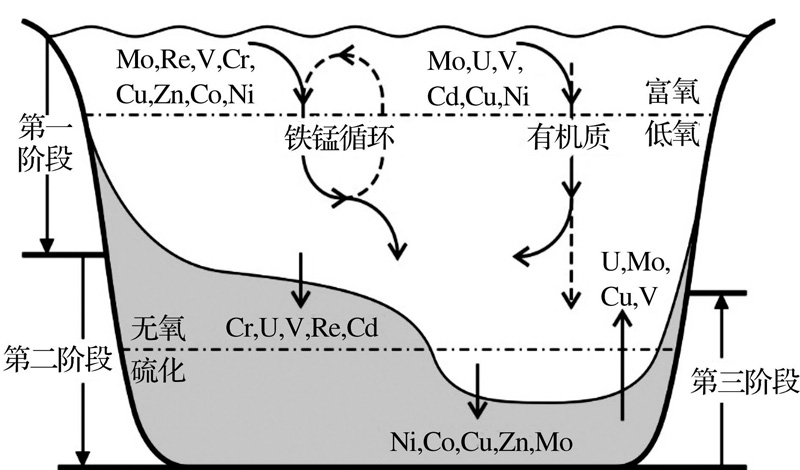 | 图2 氧化还原敏感性元素在低氧环境中的自生富集过程(据参考文献[80~89]编绘)Fig.2 Enrichment procedure of redox sensitive elements under hypoxic condition (modifiedafter references[80~89]) |
上述Mo, V和U等较为可靠的RSE指标已经广泛应用于近岸低氧沉积记录的重建中[35, 91]。综合利用其地球化学行为差异还可以辨别出水体低氧的不同程度与类型[85, 92]。
RSE和矿物含量作为低氧替代性指标固然具有方便快捷的优势, 但是一个不可避免的问题就是多解性, 这既发生在富集过程中, 又受埋藏后早期成岩作用的影响[9]。同位素指标的应用一定程度上能对这些影响加以区分。
元素往往以矿物形态赋存于沉积物中, 且在不同水体条件下会形成不同的矿物, 仅凭元素指标易发生混淆。比如上文提到的Mo除了在还原条件下会自生富集之外, 还可以在氧化条件下通过吸附在锰氧化物上沉积。Barling等[93]通过实验研究发现锰氧化物易吸附轻的Mo同位素, 导致沉积物中Mo同位素组成较海水偏轻约2‰ 。在还原环境下, 首先被从水体中清除的也是轻的Mo同位素, 但充足的S使得海水中的Mo几乎完全形成硫代钼酸盐沉积, 故最后沉积物中的同位素组成十分接近于海水[26, 94, 95]。由此, δ x/95Mo可作为区分2种富集类型的指标, x=97, x=98是目前最常见的2种测试方法。同样与水体氧化还原环境变化密切相关, 还原条件下U的分馏结果却与Mo截然相反, 即水体还原性越强, δ 238/235U越偏正[96], 或与强还原条件下U从沉积物中的移除过程, 以及核场效应(nuclear field shift)对重同位素分馏的影响有关[97~99]。不过埋藏后水体氧化还原环境的变化也会改变原有的同位素组成, 使用时需加以分辨[100]。
低氧条件下形成的沉积物自生黄铁矿本身十分亏损34S, 在波动的水体氧化还原条件下(如季节性低氧区域)易被再氧化进一步加重硫同位素的分馏[101, 102]。由此, 黄铁矿的δ 34/32S可鉴别出早期成岩过程中的一些影响, 帮助解释其他指标的变化。Wijsman等[101]利用这一特性指出, 黑海边缘沉积物中极负并波动的δ 34/32S剖面可能是由底层水体氧含量或沉积速率的变化所致。
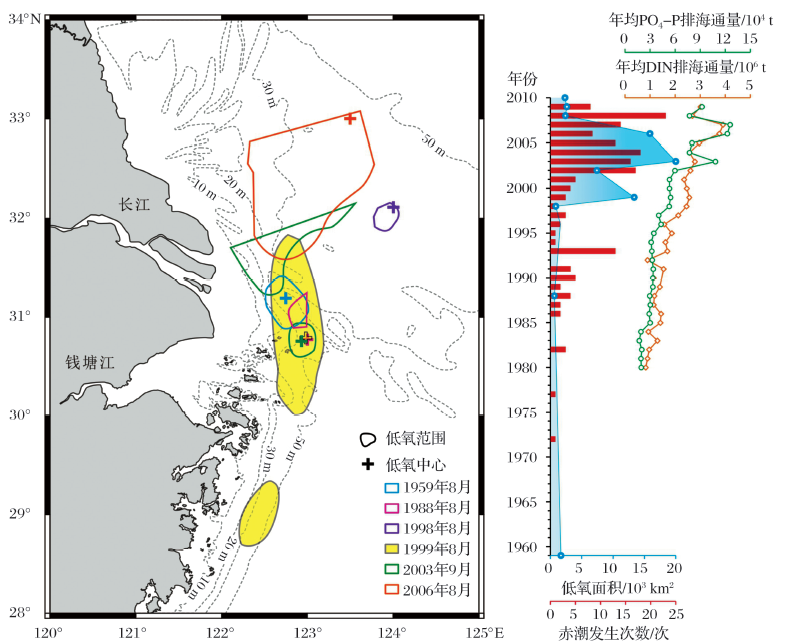
作为一条世界级大河, 长江口发育了世界上最大的季节性近岸底层水体低氧区之一。相关调查研究始于20世纪80年代, 传统认为其发育位置在122° 30'~123° E, 30° 50'N附近, 羽状锋前缘沿15~50 m水域底层[103, 104]。随着观测的深入, 人们逐渐意识到低氧的生消是一个十分动态、复杂的过程, 影响范围逐年波动, 近年来更有恶化的趋势[31, 105]。
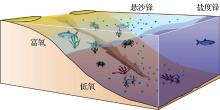
长江口底层水体低氧现象一般发生于夏季, 表层藻华过后大量有机质沉降到底部分解耗氧, 强烈的水体层化与地形又阻碍了DO的补充(图3)。
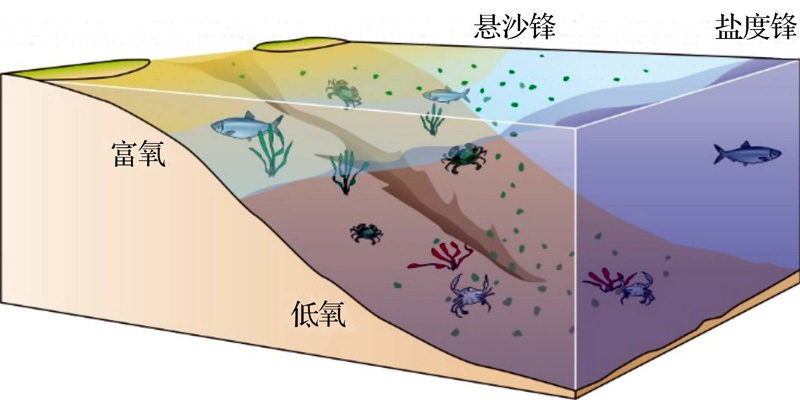 | 图3 长江口外低氧形成机制示意图Fig.3 A sketch diagram to show the formation mechanism of seasonal hypoxia off the Changjiang Estuary |
长江口低氧区有机质按其来源可分为外源输入和内源自生, 由于长江口以东及东北部海域终年存在营养盐高值区, 为形成有机质提供了丰富物质基础, 故内源自生有机质在此占主要地位[106, 107]。径流和黑潮及其分支是长江口营养物质的最主要来源。前者是夏季长江口海表营养物质的最主要来源, 但由于其氮磷比过高, 使得初级生产力明显表现为磷限制型; 后者终年向东海输入营养物质, 其表层水入侵在春夏季减弱, 而次表层水和中层水的入侵加强, 形成了长江口底部高磷酸盐水, 伴随着夏季发育的上升流抬升至5 m层附近, 进一步促使表层浮游植物勃发[108~111] 。
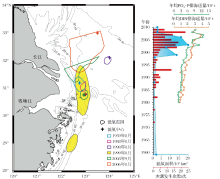
长江口夏季水体层化和海底地形都增强了底部水体的封闭性, 是维持低氧的重要条件。Wang[112]发现历年低氧区中心位置几乎都位于长江口水下深槽内(图4), 这与该处较为封闭的地形延长了水体滞留时间不无关系。大范围的低氧发育则要归功于水体层化。长江流域在春末进入洪季, 大量冲淡水进入东海, 覆于底部高盐水体之上形成盐度跃层; 同时高温表层水与底部冷水团之间形成温度跃层, 共同导致了强烈的层化现象[32~34]。
两者相互作用下, 长江口底层水体低氧自7月形成后不断加强并扩大, 至9月逐渐削弱, 持续时间较短[113]。历年观测到的DO最小值皆高于0.2 mg/L, 尚未达到次氧化环境, 氧亏损程度较低[112]。这些时空分布特征都对替代性指标的应用与解释提出了挑战。
在长江口, 对低氧的研究从现代观测走向沉积记录研究不过数十年, 兴起更只是近10年的事, 可以说仍处于起步阶段。目前有一定研究基础, 较为可靠的指标只有RSE和LOFA; 生产力指标TOC, BSi和色素, 陆源输入指标木质素酚、C/N和δ 13Corg等是常用辅助指标。
由于早期研究认为东海陆架沉积物中元素的分布受粒度影响, 但张晓东等[114]分析了表层沉积物的RSE元素组成特征发现, 虽然在底层水DO浓度正常的环境中RSE的分布显示出明显的粒控效应, 但在低氧区沉积物中仍主要受氧化还原条件的控制。许淑梅等[115]进一步去除粒度效应、陆源碎屑组分和有机质吸附作用的影响之后, 认为Mo, Cd和V的富集程度能够反映长江口低氧区的氧亏损程度, 而U则不甚理想。但是沉积后Fe和Mn受早期成岩作用影响明显, U受到Fe影响, Mo则可能受间隙水中溶解有机质的影响浓度随深度增加[116]。说明RSE的富集不是简单的沉积过程, 尤其是在长江口低氧季节性发育的前提下, 这方向尚缺乏深入研究。
目前长江口耐低氧底栖有孔虫种属仍主要沿用汪品先[117]的调查结果。已有研究通过总结全球低氧区底栖有孔虫特征, 并统计长江口耐低氧底栖有孔虫种属, 提出Epistominellanaraensis, Bolivina spp., Buliminamarginata以及Ammonia convexidorsa等为长江口低氧区的敏感种, 是合适的 LOFA 指标, Quinqueloculina spp.则是合适的氧化环境指标[55, 118, 119]。但是目前研究对长江口LOFA的认识似乎并不完整, 导致不同研究具体使用的有孔虫种属存在差别[118, 120], 且关于提出类似A-E指数、PEB指数等较为简便的指标的相关研究也亟待深入。
除了对上述传统指标的发展外, 一些新指标的应用前景也值得期待。李超等[121]研究长江口沉积物中不同化学相态Fe的分布, 发现低活性铁(FePR)分布特征与低氧区吻合, 或许能成为可靠的低氧替代性指标, 但其在沉积记录中的有效性尚待验证。随着实验条件的改善, 钼同位素作为低氧指标的运用日益广泛[26, 95], 在长江口Mo已是良好的低氧指标, 钼同位素又能对其形成环境与阶段加以区分, 提升结果的可信度。与之相似的还有耐低氧底栖有孔虫壳体的RSE/Ca比值, Groeneveld等[122]发现Buliminamarginata和Globobuliminaturgida的壳体Mn/Ca比值对瑞典西南Gullmar峡湾底层水体DO含量变化具指示作用, 也许长江口LOFA某些种属的壳体RSE/Ca也能起到类似的作用。但是考虑到长江口底层水体低氧持续时间较短、氧亏损程度较低的特征, 进行指标选择时可能要避开一些含量少、易降解的生物标志化合物和特征矿物; 对RSE、同位素和LOFA指标的多解性也要作充分考虑。
由于起步较晚, 基于长江口沉积物岩心的定年数据和替代性指标反演历史时期低氧事件的研究仅见零星报道。长江口低氧的研究在一定程度上可以借鉴同样发生在有大河输入开放型陆架区域的密西西比河口德克萨斯— 路易斯安那陆架低氧区, 但考虑到长江口低氧的持续时间更短、程度较低等差异, 很多指标在长江口可能并不适用, 使用时需重新加以论证。
通过对短柱状样百年沉积的研究, 许淑梅[123]和冯旭文[124]发现自20世纪60~70年代以来, 低氧核心区柱状样中Mo和Cd等的RSE/Al值明显上升, 与δ 13Corg指示的海区生产力变化趋势相同。Li等[55]统计的短柱状样中LOFA含量也在过去60多年里显著上升, 尤其是在20世纪80年代之后。2种指标的指示结果都与调查资料所显示的长江口低氧恶化趋势相一致, 反映了人类活动的影响。
虽然百年尺度沉积记录将低氧的矛头指向了人类活动影响, 但利用LOFA反映的千年尺度变化则显示长江口低氧区自古以来便存在。蔡庆芳[120]发现YD0903孔记录了波动式增强的低氧历史:8 600 cal a BP之前, 研究区处于滨— 浅海过渡环节, 不适宜低氧发育; 8 600~3 400 cal a BP, LOFA含量不断上升, 指示低氧呈发育态势; 3 500 cal a BP左右, 发生低氧的中断, 不过持续时间不长, 到3 400 cal a BP整体进入低氧状态, 且强低氧事件和低氧减弱事件交替出现。Wang等[118]对ECS-0702钻孔的研究结果则有所不同, 有孔虫指标显示:长江口低氧始于约4 600 cal a BP, 在4 600~2 000 cal a BP有所减弱, 600~400 cal a BP一度消失, 此后又恢复且程度更甚于以往。两岩心的记录存在差异, 一方面是用于计算LOFA指标的种属不同, 蔡庆芳[120]仅选取Bolivina spp., Epistominella naraensis和Bulimina marginata计算 YD0903 孔LOFA 指标, 而Wang等[118]使用的有孔虫种属多达15种。另一方面也可能是由于历年长江口低氧中心位置及范围不断变化所致[92], 相较ECS0702孔, YD0903孔更接近水下深槽、历年低氧中心较集中的区域(图4), 所能记录到的低氧现象更多, 表现为氧亏损开始加重的时间更早。这一差异也体现了单一钻孔对低氧演化历史反映的片面性, 多孔结合可能更好地恢复动态的区域低氧历史。
总之, 沉积记录显示了长江口近岸底层水体低氧现象自然存在, 受径流、黑潮等自然过程本身的周期性所控制, 但随着人类社会的发展, 低氧有不断恶化的趋势。早期对低氧形成机制的不了解、不重视所造成现状难辞其咎, 目前一些区域低氧情况得到改善的事实证明, 在厘清各因素的作用机制之后, 低氧是可以控制并恢复的。国际上已经建立起多种统计学、动力学模型以对低氧发展进行预测[13~15]。长江已有比较详细的营养盐和淡水入海通量数据(图4), 有望建立起多参数之间的定量函数关系, 尝试预测的同时也进一步验证机制研究的正确性。
近岸底层水体低氧现象因其对自然生态系统、人类活动和全球气候变化的影响而成为研究热点。每一次水体氧化还原环境的显著改变都会在沉积物中留下丰富的沉积学、生物学、矿物学和地球化学记录, 包含这一记录的沉积物是用来恢复低氧演化历史的理想载体。本文通过综述前人部分研究, 对近岸底层水体的沉积记录研究进行了回顾, 得到以下结论:
近岸低氧按其发育环境可分为半封闭型海盆/海湾低氧和开放型陆架低氧, 前者受控于自然条件, 低氧程度严重, 甚至孕育硫化环境; 后者在上升流区域发育很大程度上受气候变化控制, 在河口区发育则受人类活动影响显著, 多表现为季节性低氧, 程度相对较弱。
不同指标体现低氧影响的阈值不同, 各种低氧的替代性指标皆不具备普适性, 针对某一研究区域需先行论证指标的有效性, 才能进一步应用于沉积记录中。各低氧指标在形成过程中互相影响、相辅相成, 同时由于各自的缺陷, 在解释时又需要互相补充。故解释时需要选择合适的指标, 并结合多指标相互验证。
在长江口低氧区, LOFA和RSE指标的应用已有一定研究基础, LOFA指标的具体有孔虫种属组成尚未有定论, RSE较为快捷却缺乏关于季节性低氧情况下保存效率的研究, 要建立形成机理清晰、认同度高的指标还需深入研究。一些新指标的加入可能能够为此提供线索。
由于低氧的生消是个十分动态、复杂的过程, 单一孔的记录很难反映低氧演化的全貌, 结合多孔解释可能更好地反映空间上的变化过程。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|