{kind=link}
{kind=link}
稻田土壤碳循环关键微生物过程的计量学调控机制探讨
[吴金水 , 葛体达, 祝贞科]
, 葛体达, 祝贞科]
, 葛体达, 祝贞科]
|
|

作者简介:吴金水(1960-),男,湖北武穴人,教授,主要从事土壤碳循环与环境效应研究. E-mail:jswu@isa.ac.cn
稻田生态系统碳循环是我国陆地生态系统碳循环的重要组成部分,微生物驱动的稻田土壤碳循环(输入、分配、稳定等过程)的生物地球化学过程是土壤碳循环过程研究的核心。目前,对稻田土壤碳循环过程及其机制的认识缺乏基于生态化学计量学层面的研究。因此,系统解析耦合化学—生物—环境要素的稻田土壤碳循环的关键过程是深入研究当前面临的诸多土壤生物化学问题(如土壤碳循环与土壤肥力、温室气体减排等)的科学瓶颈。在综合分析计量学的基本内涵与土壤计量学发展需求的基础上,论述了稻田土壤有明显区别于其他土壤类型的土壤发生学和生物化学特点,重点评述了稻田土壤碳循环的3个主要过程的研究进展,包括:①稻田土壤新鲜有机质转化、矿化的关键微生物过程计量;②典型水稻土CH4产生的关键微生物过程计量;③典型水稻土微生物CO2光合同化功能的计量。在此基础上,探讨了土壤生物化学过程统计学和数学模型在土壤计量学研究中的应用,并提出了稻田土壤碳循环关键微生物过程的计量学特征研究的发展趋势和科学问题展望。期望能够通过这些探讨对推动我国该研究领域的基础理论建设和新技术发展有所贡献。
The paddy field ecosystem carbon cycle is a significant part of the terrestrial ecosystem carbon cycle in China. The organic carbon cycle in paddy soil (including processes such as input, allocation, and stabilization) is driven by biochemical processes that are dominated by microbes, and is the core of soil carbon cycle research. Current studies on paddy soil carbon cycle processes and their mechanisms lack metrology-level research based on the basic theories of chemistry, biology, and ecology. Therefore, systematic and quantitative analysis of the key processes in the paddy soil carbon cycle, which includes chemical-biological-environmental elements, deal with the bottleneck for the comprehensive evaluation of the carbon-sink-adding/emission-reduction effects of paddy fields, as well as the in-depth study of the current problems of soil biochemistry (such as the soil carbon cycle and soil fertility and greenhouse gas emissions). Based on a comprehensive analysis of the basic connotation of metrology and the development demands of soil metrology. This paper describes how paddy soil has an obvious discriminative soil genesis and biochemical characteristics that differ from other soil types. The paper mainly focuses on three processes of the paddy soil carbon cycle: (①measurements of key microbial processes involved in fresh organic matter transformation and mineralization in paddy soil, ②measurements of key microbial processes involved in CH4 generation in typical paddy soil, and ③measurements of microbial CO2photosynthesis and assimilation functions in typical paddy soil. Based on these measurements, we discuss the application of statistical and mathematical models of soil biochemical processes to soil metrology research and put forward the metrological development trend and the prospects of scientific issues in the research on the metrological characteristics of key microbial biological processes in the paddy soil carbon cycle. It is hoped that these discussions will promote the construction of basic theories in the research field and contribute to the development of new technologies in China.
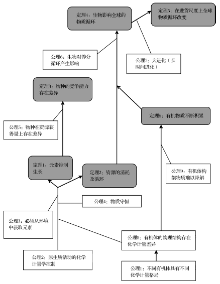
起源于传统化学的计量学(Stoichiometry)的基本科学内涵是依据定比定律、倍比定律、质量守恒等化学原理, 研究并确立物质及其化学反应与产物的元素种类与数量。生物学和生态学引入计量学之后, 从微观(生化过程、细胞、组织、个体)和宏观(种群、群落、景观, 生物圈)上推动了生物地球化学过程的定量化认识[1]。生物计量学和生态计量学不仅具有明显不同的研究尺度, 而且具有各自不同的侧重范畴。生物计量学被作为发展生物科学的统计学和数学方法。生态计量学的基本内涵是生态相互作用(Ecological interactions)的化学元素平衡, 还吸收了物理化学中的热力学第一定律和生态学中的自然选择原理、中心法则、动态平衡原理、生长速率理论等特色理论[2, 3]从而赋予了生态计量学较生物计量学更为鲜明的特色和丰富的理论基础(图1)。
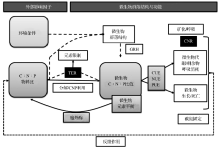
微生物的活动与环境中元素的供给及其计量比之间是相互联系、相互作用的有机整体(图2)。环境因素与元素计量在生态层面调控着微生物群落结构变化, 而营养元素计量比直接控制着微生物的C:N:P比值。元素阈值比率(Threshold Element Ratio, TER)协调微生物自身活动, 微生物通过对外源C的分解、N和P的活化利用或是微生物胞外酶的作用下, 达到微生物C, N, P的元素平衡以及对C:N:P物料比的响应; 在不同的C:N:P比值条件下, 微生物受控于计量学原理, 调节C, N, P的利用率, 通过微生物的呼吸代谢与繁殖和死亡对环境中的C:N:P元素比进行反馈调节, 从而实现在微生物驱动下的物质与元素的循环与平衡[6](图2)。生态化学计量学是生态学与生物化学、土壤化学研究领域的新方向, 也是研究土壤— 植物相互作用与C, N, P循环的新思路[6, 7]。Sistla等[8]在New Phytologist撰文指出, 生态化学计量学是生态系统碳和养分循环的调节阀, 是探索生物过程和养分循环等生态系统过程的推动力。
 | 图2 环境条件和元素计量学对微生物群落结构的影响[6]Fig. 2 Proposed effects of resource stoichiometry and environment on microbial community structure and function[6] |
土壤集中了物理、化学、生物反应过程, 是既丰富又复杂的生物地球化学体系的重要组成部分, 且各种反应过程既相互作用和影响, 又与环境条件存在依存和反馈关系。水稻土是在长期种植水稻下, 人为培育的特殊耕作土壤。水稻土除了具备一般土壤所具有的物理、化学和生物过程之外, 因其湿地属性, 还具有氧化— 还原交替过程及以此诱发的特殊化学和生物过程(如Fe, S, As等元素的氧化— 还原、有机质的嫌气分解和CH4的产生等)[9~11]。因此, 本文在探讨稻田土壤碳循环的关键微生物过程:稻田土壤新鲜有机质转化/矿化、典型水稻土CH4产生以及典型水稻土微生物CO2光合同化功能的基础上, 分析了典型稻田生态系统土壤碳循环的生态计量学调控机制, 并介绍了土壤生物化学过程统计学和数学模型, 期望能为进一步探究稻田土壤碳周转— 微生物过程的生态计量学耦合机理的研究有所贡献。
生态计量学已经发展成为生态学重要的学科分支, 具备了相当完整的理论, 研究对象十分广泛, 如种群及其演变、生物共生关系、生物体与群体营养、生态系统物质循环与限制性元素的诊断等方面, 研究尺度涵盖了生物种群、群落、景观, 生物圈[6, 12]。Elser等[13]从N, P元素计量上明确了个体与生态系统过程的统一, 从而深化了个体与生态系统的营养级、生物多样性和生物地球化学循环的系统认识。Sterner等[4]出版的《Ecological Stoichiometry》, 系统阐述了生态化学计量学的基本理论与方法。生态计量学为研究营养级动态、生物多样性和生物地球化学循环提供崭新视点, 为连贯细胞、生态系统与演变过程的“ 模糊” 机制提供了一种强有力的工具, 成为生态系统不同尺度(分子、细胞、有机体、种群、生态系统和全球尺度)统一的理论基础和关键研究工具。在土壤微生物种群、结构与多样性方面, Gans等[14]运用Model free、Zipf和Log-Laplace 3种模型, 通过改进算法, 估算出10 g原始土壤中的原核生物分类群高达107种。Harris[15]发现由退化系统到恢复初级植被群落、草原、灌木和森林生态系统演替阶段中, 土壤微生物生物量呈现抛物线变化的现象。Sardans等[7]发现气候变暖和干旱增加暖带和温带干旱生态系统的C:N:P 比率变化指示着生态系统的组成和结构的演变。Sinsabaugh等[16]指出酶活性计量是化学当量理论和代谢理论统一的纽带。在我国, 生态计量学已经逐步应用到土壤— 植物相互作用与碳循环研究[17, 18], 土壤微生物生物量C, N, P景观尺度空间研究[19]。这些研究结果都表明生态计量学在开拓生态科学研究视野、揭示生态系统重要规律等方面发挥着突出作用。
2002年成立的国际土壤学会计量土壤学(Pedometrics)专业委员会是计量学作为土壤学一个分支的标志。2003年国际计量土壤学专业委员会确定的计量土壤学定义是“ 用数学和统计学方法研究土壤的分布和发生” [20]。顾名思义, 目前的“ 计量土壤学” 仅局限于土壤地理与信息科学、统计学交叉领域, 并不涵盖土壤化学、土壤生物学、土壤生态学和土壤环境学等土壤学重要领域的计量学范畴, 更不可能以此作为解决土壤化学、土壤生物、土壤环境等方面研究所面临的计量学问题的理论和方法学基础。因此, 建立涵盖土壤生物化学的土壤计量学势在必行。
土壤集中了物理、化学、生物反应过程, 且各种反应过程既相互作用和影响, 又与环境条件存在依存和反馈关系, 构成十分复杂的生物地球化学体系。本文从水稻土有别于其他土壤的独特的生物化学特性入手, 就碳循环3类主要过程(新鲜有机碳矿化、水稻土CH4产生过程、土壤微生物CO2光合同化计量过程)的相关国内外研究进展进行概略分析。
20世纪50年代以来, 水稻土成为我国土壤科学研究的特色[9, 10]。60年来, 我国土壤学研究人员对水稻土的发生学特点与物理、化学和生物学基本性质、土壤肥力基础与改良技术等开展了大量研究, 取得了丰硕的成果[21]。近30年来, 围绕稻田碳循环与温室气体排放问题, 亦取得诸多重大研究进展, 明确了我国水稻土在近30年中具有显著的碳汇功能, 对缓解全球气候变化具有重要作用[21~25]; Wu等[26]揭示了稻田土壤有机碳积累的特殊生物化学特性, 发现稻田土壤的微生物量及微生物周转速率尽管大于旱作土壤, 但外源添加有机底物条件下的矿化率却明显较低, 且并未产生明显的激发效应, 而旱地土壤的激发效应极为显著, 同时提出了稻田土壤有机碳矿化存在阻滞效应的观点; 发现稻田土壤有机碳矿化及其对温度的响应可能并不受土壤黏粒含量的影响, 而是受底物有效性的控制[27]。从而较系统阐明了稻田土壤长期持续固碳的生物地球化学机制。同时, 系统掌握了稻田CH4排放的基本强度、季节和区域变异规律, 估算出我国和全球稻田CH4排放清单, 纠正了国际上对我国稻田CH4排放过高的估算结果[28~30], 但以往的研究积累都不足以满足建立这些过程的化学— 生物— 环境耦合的计量关系在过程研究的广度和深度上的要求。
土壤有机质转化、矿化过程是土壤中最广泛的生物化学过程, 也是土壤碳循环的最基本过程。一级动力学方程是土壤有机质矿化速率的经典模型[31]。Schimel等[32]建立了酶活性与碳库、分解常数的一级动力学理论模型:dC/dt=KC× 酶活性(其中, K表示一级动力学常数, C表示土壤碳库的量)。Ise等[33]改进了有机质周转模型, 引入了温度和湿度条件, 使其可反映全球气候变化与有机质周转的关系。
在土壤新鲜有机质转化/矿化速率与微生物种群和活性之间的关系方面, Hopkins[34]指出, 土壤有机质的微生物分解过程中, 58%~90%的碳通量通过细菌和真菌作用产生, 其余由土壤动物活动产生。Bastian等[35]研究表明, 参与小麦秸秆前期和后期分解的功能微生物种群存在显著差异。Blagodatskaya等[36]发现, 添加新鲜纤维素带来的激发效应大小与土壤中微生物种群的更替密切相关:初期以原生动物快速繁殖为主, 逐渐过渡到以真菌和G-细菌为主, 14天之后以真菌和G+细菌为主。荒漠土壤中有机碳积累量与氧化酶活性呈负相关关系[37]。森林凋落物降解过程中, 单宁类物质的损失量与过氧化氢酶活性呈显著正相关关系[38]。Allison等[39]研究发现, 土壤有机质的木质纤维指数(木质素:木质素与纤维素之和)与多酚氧化酶活性之间的关系符合一元二次方程。Sinsabaugh等[40]用化学计量方法分析全球40个生态系统的土壤酶活性, 发现纤维素分解酶、几丁质分解酶、蛋白质水解酶3个参与碳循环的功能酶活性之间及其与土壤有机质含量均呈显著正相关(p< 0.05), 且这种相关性可以通过数学模型表达。
在土壤新鲜有机质转化/矿化速率和关键微生物种群与环境条件之间的关系方面, Kellner等[41]发现, 森林土壤中降解木质素的真菌漆酶基因多样性、种群结构和表达活性随季节变化而显著变化。土壤中木质素的含量与木质素降解酶活性随土壤深度增加而降低, 且相应微生物种群也发生显著变化[42]。通过对35份研究结果进行统计发现[43], 分解有机质的多酚氧化酶和过氧化物酶活性与土壤pH显著正相关。分解土壤有机质的关键酶(木聚糖酶和胞外纤维素酶)活性与氮素含量显著负相关[44]。秸秆还田和施肥会显著改变土壤微生物群落, 且这些微生物种群结构与土壤碳氮、微生物生物量碳氮含量密切相关[45]。
稻田CH4产生是底物微生物嫌气分解的结果, 其通量与底物类型和浓度、产甲烷菌/甲烷氧化菌的活性和种群结构、土壤环境条件等因素存在计量关系。在甲烷产生速率与关键元素(化合物)之间的关系方面, Huang等[46]发现秸秆还田方式下, 1.49%新鲜绿肥和5.38%稻草以CH4形态排放。Watanabe等[47]研究表明稻草中脂类、可溶性多糖、半纤维素和纤维素影响到CH4排放强度, 之后的进一步研究建立了CH4排放与稻草施用量关系的逻辑斯蒂方程[48]。Lu等[49]表明水稻根区可溶性有机碳与稻田土壤CH4排放存在极显著线性相关关系。
在甲烷产生速率与关键微生物种群之间的关系方面, CH4排放通量与产CH4菌和CH4氧化菌的活性之间存在计量关系[50]。Ma等[51]研究表明, 水稻土中CH4排放量与水稻品种(籼稻、粳稻和杂交稻)相关, 且CH4氧化菌pmoA基因拷贝数在不同水稻品种间有数量级差异, 但其产CH4古菌的16S rRNA基因拷贝数与水稻品种无关。进一步研究发现, 不同灌溉方式对产CH4菌和CH4氧化菌活性有较大影响, 且两种微生物的变化趋势相反:产CH4菌活性与CH4排放呈正相关, 而CH4氧化菌与CH4排放呈负相关[52]。
在甲烷产生速率和关键微生物种群与环境条件之间的关系方面, 许多研究结果表明, 稻田CH4排放与温度、氧气、氧化还原电位(Eh)、pH、淹水深度、灌溉方式等土壤环境因子之间也存在着一定的计量关系[53]。7年田间观测数据表明, 土壤温度显著影响CH4排放, 温度与CH4排放关系的Q10值约为2[54]。Yang等[55]研究表明, 温度在15~37 ℃时, CH4产生与温度呈显著正相关。有机底物分解产生甲烷的量与土壤所处好氧或厌氧条件密切相关, 厌氧条件下, 甲烷产生率显著增加[56, 57]。
产生CH4的临界Eh为-215~-150 mV, 低于此Eh, CH4排放与Eh呈指数负相关关系[58]。在温带和亚北极的酸性泥炭土壤(pH 3.5~6.3)中, 甲烷产生菌的最佳pH 是5.5~7.0, 甲烷氧化菌的最佳pH 是5.0~6.0[59]。Yan等[60]通过分析亚洲稻田土壤甲烷排放相关文献, 发现土壤pH为5.0~5.5时, 甲烷排放量最大。Khalil等[61]长期观测研究发现, 稻田水深与CH4排放呈负相关关系, 即淹水层越高, CH4排放越少, 而较深的水层降低CH4排放可能与其降低了土壤温度有关。此外, 不同灌溉方式也影响CH4排放:长周期和短周期的间歇灌溉比长期淹水减少稻田CH4排放分别达59%和85%[52]。施入黑炭也显著影响农田土壤CH4排放量[62]
目前, 定量预测稻田CH4排放依然面临一些重大挑战。首先, CH4动态的模拟预测变异巨大[63~65], 其预测不确定性的根本原因可追溯到对土壤微生物过程的计量缺乏基本理解。其次, CH4排放动态几乎完全受土壤微生物过程控制, 目前国际上对其过程机理的理解尚有欠缺。
稻田土壤微生物通过光合作用同化大气CO2并形成有机质的功能(微生物光合固碳功能)是最近明确的土壤碳循环过程[66, 67]。据估算, 土壤微生物的CO2日同化速率在0.01~0.1 g C/m2之间, 同时还明确了我国典型土壤中光合微生物的主要种群和数量以及光合酶(RubisCO)活性与固碳速率之间的关系[68]。Hart等[69]运用同位素标记与色相质谱仪的技术量化了土壤化能自养细菌的固碳能力, 发现在培养40天后有4.52 g/kg 的CO2被土壤微生物所固定。Wu等[70]进一步明确了土壤RubisCO活性与土壤微生物碳同化速率的数量关系(碳同化速率=0.16× RubisCO活性-0.001; g C/m2), 可望成为估算土壤微生物碳同化速率的一种方法。
目前, 相关研究初步探讨了碳同化微生物数量和种群结构对施肥管理、作物类型、土壤质地和土壤深度的响应, 且发现了很多新的微生物固碳基因型[68, 71, 72], 但有关这些固碳微生物在土壤中所起的具体作用还有待进一步证实。基于RNA水平上的同化CO2的稻田土壤自养微生物转录活性计量特征及其调控机制研究尚未见报道。采用碳稳定同位素示踪技术, 结合基于RNA水平的分子生物学技术以及新一代高通量测序技术, 可深入发掘具有碳同化功能的微生物或者与碳同化过程相关的新的功能基因。同时通过研究微生物同化碳在土壤碳库中的转化和稳定过程的计量学调控机制, 可将碳同化微生物在土壤CO2同化过程中的贡献定量化, 这些研究将推进对微生物介导的土壤碳过程及稻田生态系统碳循环机制的认识。
土壤有机质转化、矿化过程是稻田土壤中最广泛的生物化学过程, 也是土壤碳循环的最基本过程。土壤活性有机碳库通过调节土壤能源物质和微生物活性影响着土壤碳库的有效性, 与土壤生态系统中有机碳的迁移、固持和CO2、CH4的释放有密切联系, 成为土壤有机碳分解矿化的动力和土壤质量演变的关键指标[73]。微生物分解有机质导致了有机碳的转化与呼吸利用, 这同样发生于稻田土壤有机质的转化、矿化和养分循环过程中。生态系统的养分状态强烈影响微生物营养物质的浓度和计量比。生态系统碳循环和碳固定效率取决于关键养分元素输入量和需求之间的平衡, 以及养分的有效性。稻田生态系统碳循环的稳定性不仅会受到相关生物体对元素需求的强烈影响, 也会受到周围环境化学元素平衡状况的影响, 在相对稳定的条件下, 稻田生态系统碳循环的微生物过程是由质量守恒原理和其它关键养分元素(如氮、磷、硫等)的计量比控制的。
土壤有机质的活性组分包括轻组分有机碳、水溶性有机碳以及微生物生物量碳、易氧化碳等组分, 由糖类、氨基酸和大部分未分解的有机碎屑组成, 在土壤中不稳定, 周转速率快, 易矿化分解 [74]。稻田土壤中微生物易利用态碳在土壤碳的周转过程中起着重要的作用, 特别是可溶性有机碳(Dissolved Organic Carbon, DOC)和微生物生物量碳(Microbial Biomass Carbon, MBC)含量与土壤有机碳矿化之间呈显著正相关 [75]。而在全球温室效应加剧, CO2浓度不断提高的背景下, 稻田土壤自养微生物参与大气CO2的同化过程, 对于土壤碳库的贡献日益显著 [22]。
土壤中的氮、磷是植物生长、微生物活动所必需的矿质营养元素, 是生态系统中最常见的限制性元素。土壤N素的有效性往往和土壤C库联系在一起, 这是由于土壤本身可利用C, N基质的数量是土壤有机碳矿化的主要限制因素[76, 77]。由于稻田生态系统的复杂性, 稻田土壤碳、氮转化之间的关系较为复杂, 并没有一致的结论。外源 N, P, S等元素的输入能促进或阻碍土壤有机碳的矿化(产生CO2和CH4), 引起正的或负的激发效应[78~80]。同位素示踪培养法的研究则认为, 添加无机氮使土壤有机碳矿化表现出不同程度的下降, 无机氮对土壤有机碳转化的贡献在于相对增加了土壤有机碳的固持, 表明无机氮对土壤固有碳的矿化起到抑制作用[81, 82]。而Nelson等[83]对比添加N和未添加营养物质的土壤溶液矿化实验发现, 表层土壤碳矿化释放率提高了25%, 底层没有明显变化, 这可能是由于表层土中大量的根系分泌物和可溶性根系组织的输入, 提高土层DOC的微生物降解率, 降解处于N限制状态; Keller等[84]研究结果显示, 施用N, P 肥料和石灰对微生物碳循环没有太多影响, 但是增加了土壤碳矿化。目前关于P的有效性对碳矿化影响的研究较少, 有培养试验表明, 外源 P 输入量的增加, 土壤有机碳的矿化速率和累积矿化量均增大 [85]; 相同 P输入水平下, 淹水处理时土壤有机碳的累积矿化量, DOC和可溶性无机碳含量均高于非淹水处理 [86]。因此, 土壤中碳氮磷的可利用性以及供应与输入水平, 对土壤碳循环起着关键的调控过程。
碳、氮、磷等元素对生物个体和生态系统机构与功能都具有重要作用, 而且碳、氮和磷元素的循环过程相互耦合, 所以, 关键养分(如碳、氮、磷)及相关生物体对元素需求的改变将明显影响生态系统碳循环的微生物过程[87, 88]。人类活动对碳、氮、磷素的生物地球化学循环的影响逐渐加剧, 而C素的生物地球化学循环也取决于其和N、P等养分元素间的平衡关系, 基于生态化学计量学原理, 土壤C固定取决于N和P对微生物的影响[84, 89]。微生物可调节所分泌的生态酶的比例, 进而获取自身所需的元素。如果养分供应不足将会导致微生物对所缺元素生态酶生产能力的增加, 且同样表现出相应的计量特征 [90, 91], 进而影响到土壤中有机质的分解速率。研究表明, 土壤中微生物生物量碳(MBC)与微生物生物量磷(Microbial Biomass Phosphorus, MBP)的比值(MBC:MBP)可用于评价微生物是通过矿化有机物释放磷素或从环境中竞争吸收磷的重要指标。MBC:MBP比值降低表明微生物会经土壤有机质矿化释放进而补充土壤有效磷库, 且加剧了微生物的碳限制, 进而分泌相应计量比值的生态酶来加强碳获取能力; 而MBC:MBP比值增加则表明微生物经同化有效磷, 并与植物竞争磷素, 表现出微生物的固磷能力。土壤中有机质的分解为土壤对土壤的生态服务功能具有重要的意义, 可表明碳库的源汇效应, 然而这一过程会受到氮磷元素的影响[90, 92], 因此, 研究营养元素计量比值对土壤碳循环具有必要的意义, 可以为提高稻田土壤肥力和减缓稻田温室气体排放提供必要的理论依据。
现代生物统计学和数学模型的发展, 为研究土壤生物化学过程中的计量关系提供了重要的研究手段。基于区域变量理论的地统计学方法由分析空间变异与结构的变异函数及其参数和空间局部估计的Kriging(克里格)估值法2个主要部分组成, 是用于分析土壤元素、生物特征及地理环境等变量的空间变异性的有效方法[93]。由区域变量理论发展出的普通克里格、协同克里格及回归克里格已成功应用于土壤有机碳、氮磷空间变异规律研究及预测[94]。理解土壤生物化学过程中土壤元素、生物、地理环境及气候等因子互作有重要意义。回归模型、主成分分析、因子分析及典型相关分析等线性数学模型方法常用于分析土壤养分、微生物与地理环境及气候等因子的相关性及其影响因子。Taylor等[95]采用甲烷排放量与作物生物量或籽粒产量的回归关系来预测甲烷排放。Huang等[46]研究结果表明稻田产生的甲烷被氧化的比例与地上部生物量与收获时最大生物量的比值有关。Lu等[49]利用培养实验发展了预测甲烷产生的回归模型。Elser等[96]通过回归分析, 建立陆地和海洋草食动物食物中C:N及C:P比值与动物的初级生产力之间的关系。Chow等[97]建立了土壤碳矿化模型, 该动力学概念模型可以较好的描述土壤SOC、DOC浓度和碳矿化的关系, 有利于理解土壤碳有效性与碳矿化的关联性。该模型认为土壤微生物可以分别利用SOC和DOC作为基质矿化产生CO2, 微生物菌也可以利用 SOC转化为 DOC, DOC浓度和碳矿化的相关关系取决于kDOC与kSOC(kDOC: DOC矿化产生CO2的反应速率, kSOC: SOC矿化产生CO2的反应速率)的大小, 假如由微生物菌形成的不稳定C在DOC中比SOC高, 那么kDOC> kSOC, 土壤碳矿化速率和DOC 的浓度正相关, 土壤碳矿化主要是由 DOC 来控制的, 多数的研究也表明如此; 反之, 土壤碳矿化速率和 DOC的浓度负相关。动力学概念模型虽然比较好的描述了有机碳与矿化速率的关系, 却难以解释总碳矿化潜能。最初为了模拟美国农业土壤N2O的排放而研发的DNDC模型, 能够模拟和预测温室气体排放量, 而且能够评估不同管理方式的减排效果, 还可以通过模拟土壤生物化学过程的交互作用, 定量评估一种温室气体的减排措施对其他温室气体排放量的影响, 并且验证该措施是否具有其他的不利影响, 如农作物是否减产等[97, 98]。陈海心等[99], 为探讨长期不同施肥条件下土壤有机碳和作物产量的变化规律及DNDC模型在关中地区的适用性, 长期定位施肥试验数据, 分析了20年来土壤有机碳(SOC)含量、 碳循环以及农作物产量的变化趋势等基本特征。利用实测数据标定和验证了DNDC 模型, 通过均方根误差(RMSE), 标准化的均方根误差(N-RMSE)以及符合度指数(d)对模型适用性分析, 模型预测精度较好, 可用于预测该地区作物产量和农田土壤有机碳动态。线性数学模型具有模型结构简单、结果解释性强的特点, 然而线性数学模型要求满足变量关系的全局平稳假设。实际研究中土壤属性、生物特征及地理环境等因子的空间分布是非平稳性, 这导致基于全局统计模式的线性模型很难输出可靠的结果。为了有效地表征空间变量的局部关系, 地理加权回归模型(geographically weighted regression)[100]、局部回归克里金方法(local regression kriging)相继提出, 这2种方法构建空间依赖关系时不再用样本的全局统计信息而是用与未知位点最邻近的k个样本。与基于全局模式的线性数学模型方法相比, 地理加权回归模型和局部回归克里金方法由于考虑了变量空间分布的不平稳性, 模型输出结果更可靠和准确。
由上述国内外研究现状分析, 稻田碳循环微生物过程的计量学特征所在研究领域的研究发展趋势和科学问题展望如下:
(1)尽管计量学成为确立化学、生物学和生态学的定量方法的基本理论和方法基础, 在土壤学中的应用却存在很大的局限性, 建立涵盖稻田土壤生物化学的土壤计量学是土壤学发展的迫切需要。
(2)新鲜有机质转化/矿化、CH4产生和大气CO2微生物光合同化等是密切关系到土壤肥力、环境和全球变化的关键土壤生物地球化学过程。尽管以往的研究基本明确了这些过程的基本构成(如主导底物、中间过程主要产物和最终产物、功能微生物种群, 以及土壤物理性质、pH、Eh、底物丰度、养分供应状况等环境条件), 也探讨了其中某些要素之间的关系, 但远未满足构建土壤生物化学计量学的研究积累, 也难以为解决当前土壤生物化学研究所面临的重大科学问题(土壤肥力提升、温室气体减排等)提供充分的理论基础。
因此, 运用化学计量学、生物计量学、生态计量学的基本原理, 以典型水稻土为对象, 通过对水稻土新鲜有机质转化/矿化、CH4产生和大气CO2微生物光合同化等3类较具普遍性, 且化学— 生物— 环境耦合性强的关键过程的主导底物及其参与物质(如相关有机分子、O2、CO2微生物光合同化过程的光子等)、中间过程的主要中间产物和最终产物(CO2、CH4、有机碳)的比率及其与驱动这些过程的微生物(功能群数量、活性及其关键土壤酶计量特征等)和环境条件(如土壤物理性质、pH、Eh、底物丰度、养分供应状况)的定量关系, 探讨建立土壤生物化学计量学的可能性(包括计量学基本原理的选定、土壤计量学的基本方法体系、典型过程的计量学解析), 同时在计量学层面系统和深入地阐明当前高度关注的水稻土碳循环3个基本过程的化学— 生物环境耦合关系与生态计量学特点。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|