
{kind=link}
{kind=link}
{kind=link}
{kind=link}
中国新近纪哺乳动物群的演化与青藏高原隆升的关系
[邓涛1, 2, 3  , 王晓鸣
, 王晓鸣1, 4 , 王世骐1, 2 , 李强1, 2 , 侯素宽1 ]
, 王晓鸣|
|


作者简介:邓涛(1963-),男,四川宜宾人,研究员,主要从事晚新生代哺乳动物、陆相地层和气候环境研究. E-mail:dengtao@ivpp.ac.cn
新近纪哺乳动物的起源、辐射、扩散、绝灭等方面反映了气候环境的重大变化,晚新生代青藏高原的形成对东亚地区哺乳动物的演化具有直接而深远的影响。高原的隆升使自然环境发生改变,环境的变化又导致动物组成和分布的分异,以及动物对环境适应性的调整。中国新近纪哺乳动物化石研究的结果不仅表明哺乳动物群的组成和特征具有明显的区域性差异,而且中国还是一些代表性门类的起源扩散中心,也是哺乳动物洲际迁徙的关键通道。青藏高原新近纪晚期的哺乳动物群具有非常重要的意义,因为在随之而来的第四纪冰期中这些寒冷适应性动物获得了优势地位,由此成为现代动物地理区系和多样性的基础。通过以点带面的综合研究,将能够全面地认识新近纪动物地理区系对重大气候事件的响应,更准确地判断青藏高原隆升对陆地生态系统的影响。
The origin, radiation, dispersal, and extinction of the Neogene mammals provide important insights in our understanding of mammalian geographical distribution and regional differentiations, and such an evolutionary history can, in turn, reflect significant changes in climate and environments. Mammals are very sensitive to climatic and environmental changes, and their distribution is closely related to their natural habitats. Throughout the late Cenozoic, the formation of the Tibetan Plateau also had a profound influence on the evolution of mammals in East Asia. The uplifting plateau was a major drive in changes of natural environments, which have a direct impact on the differentiation of the faunal composition and distribution. The Chinese Neogene mammalian fossil records are unparalleled in the world, and with its unique zoogeographical position, such a record of faunal compositions is sensitive to regional differentiations. Furthermore, China was a center of origin and dispersal for many representative mammalian groups, and also a key intersection for intercontinental migrations. Most taxa of the distinctive modern mammalian fauna of the Tibetan Plateau have long life history in this plateau, which indicate that they have long-term adaptation procedures within the high-elevation plateau. The late Neogene mammalian fossils from the Tibetan Plateau suggest that some Quaternary mammals first evolved in Tibet before the beginning of the Ice Age. The cold winters in high Tibet served as a habituation ground for the members of the megafauna, which became pre-adapted for the Ice Age, successfully expanding to the Eurasian mammoth steppe. As a result, the research on the Neogene mammalian faunas of the Tibetan Plateau will be focused, because these cold-adapted mammals were dominant in the subsequent Quaternary Ice Age, and become the basis of the modern mammalian zoogeographical patterns and biodiversities. Taking advantage of the above favorable conditions, the response of the Neogene zoogeographical regionalism to the major climatic events and more accurately judge the influence of the Tibetan Plateau uplift to the terrestrial ecosystem can be more accurately judged.
气候有其自然的演变规律, 这个规律必然对地球上的生物, 包括从新生代开始繁盛的哺乳动物产生影响。哺乳动物的起源和演化无不受其生活环境的调控和制约, 化石物种及其组合的特点与变化又是了解和再现其生活地区的气候与各种环境因子的最好依据[1, 2]。因此, 哺乳动物是反映气候环境变化的重要标志之一, 基于这一逻辑关系, 哺乳动物化石所诠释的演化历史能够判别新近纪气候环境的重大变化[3, 4]。另一方面, 主要归功于高分辨率深海氧同位素曲线的建立, 新生代全球气候的复杂变化正得到越来越多的认识, 其细节也变得越来越清晰[5], 但包括哺乳动物在内的陆地生态系统的响应则还需要深入的了解, 相关的陆相沉积物尚待进行更细致的工作, 并且要建立精确的年代框架, 以利于哺乳动物地理分布和区系格局形成过程的同时性对比。
动物的演化和区系的分异与动物本身的进化机制、古地理及气候环境等综合因素密切相关。中国现代的自然地理和环境格局, 很可能与哺乳动物多样性的状况相似, 在青藏高原显著隆升的新近纪时期已基本形成。在陆地生态系统的分化过程中, 全球气候的变化显然也对哺乳动物的地理分布和区系格局产生了影响, 使一些类群的分布界限随着气温的变化而改变, 这种变化在古北界和东洋界的过渡地带上反映尤为明显, 具体表现在其范围的扩大或缩小。因此, 新生代晚期哺乳动物地理区系的研究重点是青藏高原的新近纪寒冷适应性动物, 同时与中国不同地理分区内新近纪哺乳动物群的生态特点进行对比, 将能够阐明东部季风影响范围内华南地区、华中过渡带和华北地区哺乳动物的适应性特点, 从动物群演替的稳定性以及动物群组成和属种更迭的速率等角度反演东亚地区哺乳动物的生活环境, 从而建立哺乳动物地理区系格局与青藏高原隆升驱动的气候环境变化之间的相关性。
中国在晚新生代的哺乳动物方面有着世界上得天独厚的条件, 沉积盆地发育、化石地点众多(图1b, c)、标本数量巨大、生态类型丰富, 南北跨越热带、亚热带和温带, 东部为季风森林区, 西部为干冷疏林草原和荒漠。中国新近纪哺乳动物化石的分布相当广泛, 从青藏高原的札达盆地到蒙古高原的中部地区, 从甘肃临夏盆地的红粘土堆积到山西榆社盆地的河湖相砂岩, 从昆仑山口的高寒地带到云南元谋的干热河谷, 都在陆相沉积中发现了非常富集的哺乳动物化石埋藏, 具有其他国家不可替代的研究资源优势[6~8]。如欧洲新近纪的哺乳动物化石地点分散、地层不连续, 因此时代和环境背景都很难准确判断[9]。
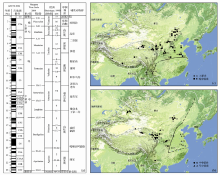
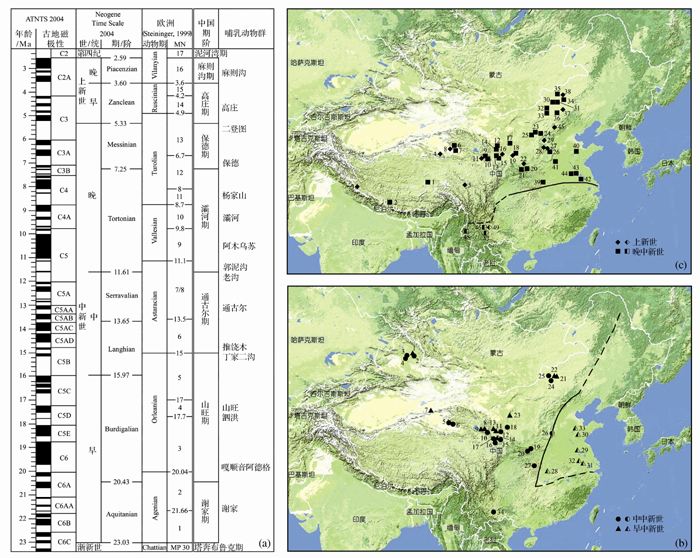 | 图1 中国新近纪哺乳动物群的时代和地理分布(修改自文献[7])(a)新近纪哺乳动物群序列; (b)早、中中新世化石地点; (c)晚中新世、上新世化石地点Fig. 1 Age and geographical distribution of the Neogene mammalian faunas in China (modified from reference [7])(a)Sequence of the Neogene mammalian faunas; (b) Early and Middle Miocene fossil localities; (c) Late Miocene and Pliocene fossil localities |
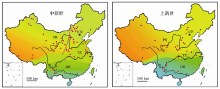
根据中国的丰富化石材料, 已经建立起新近纪的哺乳动物区系概念模式(图2)[4], 识别出哺乳动物在欧亚大陆内部以及与北美之间的迁徙事件[10~14], 发现了冰期动物群在青藏高原的起源以及随后的扩散路线[15]。作为生物进化事件与气候环境演变关系研究的基础, 精细的年代地层框架的建立至关重要。正是由于中国新近纪哺乳动物化石的富集, 以其作为主要划分依据的时代分期已经建立(图1a), 这一框架不仅成为中国的标准[16], 也在国际上被接受作为代表亚洲地区的方案[7, 8, 17, 18]。中国在新近纪哺乳动物地理区系与气候环境背景关系的研究方面, 不仅有优异的化石和剖面条件, 而且在研究手段上与国际同行既不相伯仲又有密切合作, 利用釉质稳定同位素、牙齿微磨痕、物种多样性、冠高指数、体重群落线、运动功能分析、多元多变量统计等方法开展了卓有成效的工作[19~25]。
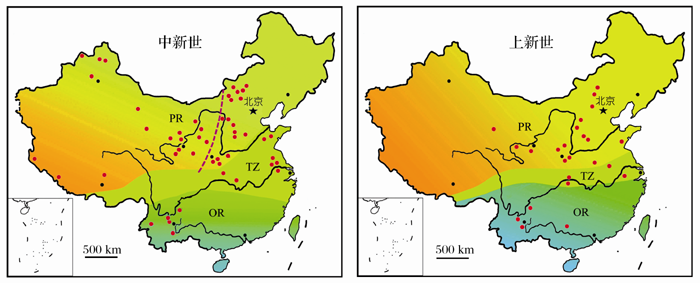 | 图2 中国新近纪哺乳动物地理区系的初步研究结果(修改自文献[4])PR:古北界; OR:东洋界; TZ:过渡带Fig. 2 Preliminary patterns of paleozoogeographic provinces of Neogene mammals in China (modified from reference [4])PR: Palearctic Region; OR: Oriental Region; TZ: Transitional zone |
然而, 关于中国新近纪哺乳动物地理分布和区系格局的特征及其对气候环境变化的响应尚未形成有规模的研究, 近年来在中国报道的重要新近纪哺乳动物化石只利用了化石宝库中的部分材料, 还有不少关键的生物进化事件和气候环境过程有待更多的哺乳动物化石来证实, 以便深入地了解并发现其内外因素之间的紧密关联。如哺乳动物地理区系演化的驱动机制以及洲际间哺乳动物交流的气候环境背景等都是广受关注的重要课题[14, 26, 27]。新近纪大量珍贵哺乳动物化石材料的新发现所引发的更多涉及过去全球变化的关键科学问题亟待回答, 包括:第四纪之前冰期动物群在何种气候环境条件和地质构造背景下起源?为什么现代青藏高原具有大量独特性哺乳动物?现代哺乳动物地理区系的格局在什么时间和什么环境下定型?
从图1和图2可以看出, 由于有丰富的哺乳动物化石地点和连续的地层剖面, 中国东部的新近纪动物地理区系特征较为清晰, 而在西部地区, 尤其是青藏高原, 我们的了解尚处于相当薄弱的状态。近年来的研究表明, 全球环境演化历程中许多重大事件与亚洲有关, 其中青藏高原的隆升被认为是全球晚新生代气候变化的重要因素, 对现代东亚季风环境的形成演化和西风环流的行为有着重要的影响。在青藏高原隆升的驱动下, 东亚古季风逐渐加强, 亚洲内陆逐渐变冷、变干, 中国北方出现了富含哺乳动物化石的风成红粘土堆积[28~30]。植被的光合作用类型在中新世末期从C3循环转变为C4循环[31], 中国西北受青藏高原的隆起而延缓了这个过程, 而中国东部和南部的新近纪地层中则清晰地记录了C4植物的扩散[32~34]。
哺乳动物群的组成和特征以及青藏高原周边地区的巨厚砾石堆积表明, 青藏高原在新近纪持续抬升。最近在西藏札达盆地发现了已知最原始的披毛犀、雪豹和北极狐, 从而推翻了冰期动物起源于北极圈的假说, 证明青藏高原才是它们最初的演化中心(图3)。
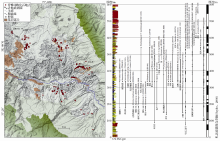
青藏高原的新近纪哺乳动物群具有特殊的意义, 因为在随之而来的第四纪冰期中这些寒冷适应性动物获得了优势地位, 由此成为现代动物地理区系和多样性的基础。鉴于札达盆地的重要性, 它成为探索中国新近纪哺乳动物地理变迁和现代哺乳动物多样性格局形成的关键地区。在地质构造上, 札达盆地位于拉萨地块与喜马拉雅构造带接触部位。盆地东西长约140 km, 南北最大宽度为50 km, 盆地内的新生代地层近水平产出, 超覆于盆地基岩侏罗— 白垩系变质砂岩之上。地层出露厚度在盆地南缘达到最大, 约800 m左右, 下部为快速堆积的含砾砂岩夹薄层泥岩, 中部为砂岩与泥岩、泥灰岩互层, 上部为巨厚砾岩与薄层泥灰岩、泥岩互层。象泉河从盆地穿过, 切开了重重地层, 形成大面积的露头和优良的剖面, 成为研究青藏高原新生代沉积的有利地点。近年来已在札达盆地发现了200多个化石地点和数以千计的脊椎动物化石标本(图4), 已鉴定出的动物化石共计33种, 包括裸裂尻(裂腹鱼类)、鸵鸟(蛋片)和31种哺乳动物, 证实札达盆地是目前青藏高原上脊椎动物化石多样性最高的晚新生代盆地[35~37]。
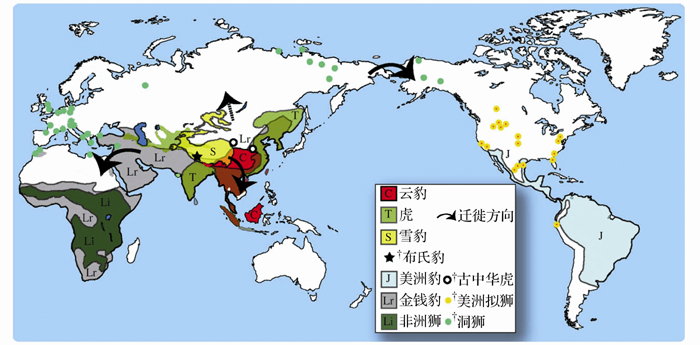 | 图3 雪豹在青藏高原的起源及豹亚科的全球扩散(修改自文献[35])Fig. 3 Origin of the snow leopard in the Tibetan Plateau and the global expansion of the Pantherinae (modified from reference [35]) |
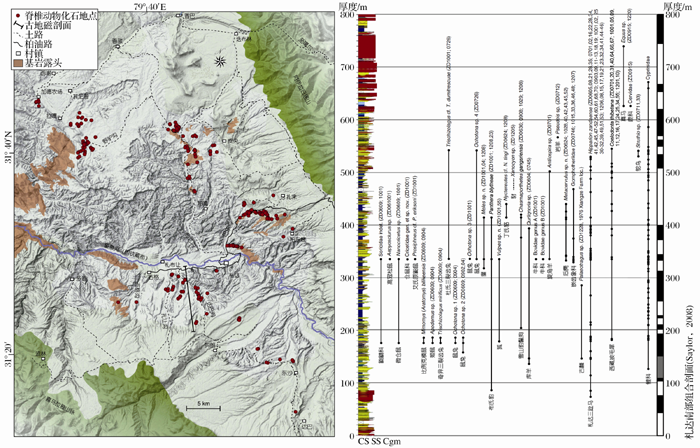 | 图4 西藏札达盆地的新近纪哺乳动物化石地点和层位(修改自文献[36])Fig. 4 Localities and horizons of the Neogene mammalian fossils in the Zanda Basin of Tibet, China (modified from reference [36]) |
冰期动物群曾被假定是随第四纪冰盖扩张进化而来, 即这些动物被推断可能起源于高纬度的北极圈地区, 但此说一直没有可信的证据。来自西藏的化石材料证明, 冰期动物群的一些成员在3.7 Ma前的上新世中期已在青藏高原上演化发展。冬季严寒的高海拔青藏高原使冰期动物群形成对严寒气候的预适应, 在第四纪成功地扩展到欧亚大陆北部的干冷草原地带[15]。
札达动物群的其他成员以及在青藏高原其他地点发现的哺乳动物化石已经显示, 独特的青藏动物群可以追溯到晚中新世时期[38]。岩羊的祖先也出现在札达盆地, 在随后的冰期里扩散到亚洲北部, 与披毛犀的演化历史非常相似。此外, 分子生物学家已经建立起牦牛和盘羊在青藏高原或周边山地的祖先类型与其北美的冰期动物亲戚, 如美洲野牛和加拿大盘羊之间在系统发育上的联系[39~42]。牦牛、藏野驴、藏羚羊和北极狐也都具有青藏高原起源的线索和向北扩散的路径, 藏野驴甚至远及阿拉斯加。在札达发现的豹类化石代表了世界上已知最古老的大猫(即包括狮、虎、豹在内的豹亚科), 由此不仅揭示了雪豹, 而且揭示了整个豹类以及其他豹亚科动物的青藏高原起源[35]。适应寒冷气候的第四纪冰期动物群的起源, 原来一直在上新世至早更新世的极地苔原和干冷草原上寻找。显然, 在青藏高原上发现更多的哺乳动物化石, 将提供进一步的证据来证明全北界的冰期动物群在上新世青藏高原的寒冷环境中渡过了适应进化的最初阶段, 并可能将多数现代青藏高原哺乳动物的历史追溯到新近纪的本地起源, 在哺乳动物演化发展与气候环境背景和青藏高原隆升的关系上取得更大的突破。
不仅在青藏高原起源的动物通过扩散影响新近纪的动物地理格局, 中国其他地区的动物多样性特点也受到青藏高原隆升的强烈影响。中新世是我国哺乳动物向现代转变的一个重要时期, 现生哺乳动物中大部分科在这一时期先后出现。晚中新世是我国哺乳动物在地史上多样性最高的时期, 种类和数量都相当丰富, 华北和西北的三趾马动物群与现代非洲草原上的哺乳动物一样繁盛[43, 44]。中新世哺乳动物在我国的分布出现了明显的南北分异, 这种分异尤以小哺乳动物更为显著[6]。中新世北方动物群反映了温带草原环境, 与现代古北界相似; 南方动物群则指示了热带— 亚热带森林环境, 具有现代东洋界的特色; 其间有一个明显的动物分布过渡区, 这一地区混杂了南方和北方的动物类型[45]。我国中新世的哺乳动物分布也发生了东西分化, 东部主要由喜暖湿的成员组成, 西部主要由耐寒、耐旱的草原型动物组成[46]。中新世中期的“ 铲齿象动物群” 反映了相当暖湿的气候环境, 可能与全球15 Ma时的气候变暖有关[47]; 晚期“ 三趾马动物群” 以及“ 三趾马红土” 在华北和西北的广泛出露, 表明了这一时期的环境又发生了一次较明显的变化, 气候变得干热[26]。青藏高原在晚中新世的隆升高度显然已对季风产生了较大的影响, 因而出现了土状堆积的大面积扩展, 导致我国动、植物的分布产生如此明显的分异, 形成与现代古北界和东洋界十分接近的动物区系。随着青藏高原隆起而被抬升了的蒙新高原区, 这时的高度也已使东南和西南季风对其的影响变得很弱, 气候处于半干旱甚至干旱状态, 出现了只适于耐旱动物生存的环境, 而使东部和南部喜暖湿动物无法扩散到这一地区[4, 48, 49]。
中国的上新世哺乳动物群继承了晚中新世“ 三趾马动物群” 的面貌, 但在演化上只发生科、属级的替换。南方的哺乳动物群分化比晚中新世时更为清晰, 繁衍了喜湿热的森林型动物[4, 50]。北方的哺乳动物群主要由适应草原生活的成员组成, 指示了典型的古北区自然环境, 如三趾马、骆驼、羚羊、跳鼠、沙鼠、原鼢鼠、鼠平鼠和鼠兔等[6, 49]。与晚中新世相比, 啮齿类动物不仅在种类和数量上明显增加, 而且齿冠也高, 适应取食硬草的能力得到加强[51~53]。上新世时青藏高原北侧和西侧的中亚和中国西北变得非常干燥, 草原化和荒漠化比中新世更为明显[54~56], 而高原的东部和南部的变化却很小[20, 57, 58]。
中国新近纪的哺乳动物地理区系研究已有很好的工作基础, 以哺乳动物化石为标准, 已经建立起由中新世的谢家期、山旺期、通古尔期、灞河期和保德期, 上新世的高庄期和麻则沟期组成的地质时代分期框架(图1a), 并得到精细的古地磁定年数据支持[7], 部分地区的哺乳动物群性质已得到深刻认识[8]。青藏高原隆升驱动的气候环境变化及其对哺乳动物地理区系的影响也在某些方面得到了解, 已经发现青藏高原是第四纪冰期动物群和现代高山动物群的起源中心。这些成果已经突破了资料积累的瓶颈, 提高到原创性的理论层面, 不仅在科学界引起极大关注, 也为公众认识哺乳动物多样性的深刻内涵普及了重要知识[15, 25, 35, 47]。
开展相关研究工作已有一系列的方法和手段可以采用或借鉴。在哺乳动物地理演变的研究中, 系统发育分析至关重要, 而利用包含形态和遗传特征数据集的“ 全证据(total evidence)” 方法重建祖裔关系使结果变得更为精确[35]。用来构建生命演化树的数据有两种主要的类型:一种是表型数据, 也就是可观察的解剖和行为特征; 另一种是DNA所编码的基因数据。随着学科交叉的日益密切, 构建强有力的系统树需要整合两种类型的数据, 因为无论是基因数据还是表型数据, 单一采用任何一种都会遗漏另外一类有效信息。另一方面, 尽管在有关物种关系的研究中, DNA序列数据起到非常重要的作用, 但是表型数据在直接构建树的过程中起到主要的作用。这些表型数据包括了保存在化石中的特征, 而化石中保存DNA通常是不可能的。因此, 只有把所有的证据都结合起来才能对过去的事件做出信息量最大的重建[59]。
青藏高原的形成使得来自印度洋和太平洋的季风都难以直接影响到亚洲大陆的中心部位, 必然导致这一地区荒漠化和我国自然环境产生分异。由于高原隆升, 蒙新地区气候已变得干旱, 植被趋于草原化, 使那些适应草原和耐旱型动物得以繁衍。哺乳动物颊齿冠高指数的增长与干旱度和生态环境的开阔程度有关, 其与降雨量之间的关系来源于对现生动物食物性质与牙齿磨蚀程度之间的观察。增加的植物纤维、累积的硅质、额外的沙尘、降低的营养物质都增强了牙齿的磨蚀。所以根据高冠指数可以推测植被的条件, 由此与古水汽条件相联系, 进而可以推测降雨量。
在古温度恢复方面, 食草动物和水体的稳定同位素分析是可以利用的有效手段。高冠食草动物牙齿釉质的稳定碳同位素比值与其取食的草本植物相关联, 能够指示生活环境中植被的C3或C4植物类型, 经过地质时期大气二氧化碳碳同位素比值变化的校正, 即可以通过植被类型定性地得到相关的古温度范围, 再通过骨骼化石的氧同位素温度代用指标以及碳酸盐二元同位素温度计得到定量化的古温度数值。
在新近纪全球气候变化的背景下, 东亚地区因青藏高原的强烈隆升而造成生态环境的多元化格局, 因此, 精细的哺乳动物适应性研究分析至关重要。在哺乳动物群内部, 迁徙性较强的大型哺乳动物可以反映广泛区域的气候特征; 小型动物克服分布阻限的能力相对比大、中型动物弱, 在长期的演化和替代过程中更易受周围环境的制约, 其分类阶元的亲缘关系与自然环境的相关性更为明显。这样, 低级类元的动物群体的地理分布范围必然相对狭窄。所以, 小哺乳动物对气候环境变化的反映比大型哺乳动物更为敏感, 因而研究小哺乳动物能更精确地反映地区性的气候环境特征。
构成陆地生态系统重要成分的哺乳动物所记录的各种变化不但能为从地形和沉积物所得出的自然环境变化提供极有价值的补充, 还能用来以较高的精度和可靠性重建古气候的波动情况[60]。陆地生态系统在新近纪发生了剧烈的变化, 森林植被为草本植被所代替, 随即导致了动物群的分异和演化, 森林动物大量消失, 哺乳动物从铲齿象动物群突变为三趾马动物群, 造成喜暖动物群的分布范围向现代的亚热带和热带地区迁徙[61], 而干旱和湿润作用的叠加还引起哺乳动物地理区系在东西方向的差异和变化[46]。根据植物光合作用类型与食草哺乳动物骨骼形成上的联系, 利用哺乳动物化石牙齿釉质的碳、氧稳定同位素组成已经重建了青藏高原不同新近纪盆地的古环境和古高度, 为检验构造、气候和生物演变之间的联系提供了一个独特的窗口[34]。剖析新近纪气候频繁波动的历程中哺乳动物地理区系的变化模式, 判断动物群的起源、辐射、迁徙、消亡和绝灭的具体细节, 这些研究方向近年来倍受古生物学家和古气候学家以及更广泛的同行学者共同关注, 同时也是发展最快的自然科学前缘领域之一, 在这些方向上取得的成果还将为预测和应对未来的全球变化提供极其重要的过去全球变化的记录和信息, 因此具有回应社会迫切需求的功能。
新近纪青藏高原的隆起对东亚地区哺乳动物的演化具有直接而深远的影响。由于高原的逐渐隆升, 南亚夏季风和北亚冬季风的强化, 我国西北部的气候越来越干旱, 青藏地区越来越高寒, 自新近纪以来所造成的恶劣生态环境已使青藏高原成为许多哺乳动物不可逾越的障碍。然而, 在新近纪青藏高原的极端环境下仍然有丰富化石所反映的繁盛哺乳动物生存, 研究它们的适应辐射机制, 不仅能够提供独特生态系统的结构特点, 还能为第四纪的冰期动物群以及青藏高原和北极的现代耐寒动物找到起源的原始地域和演化的详细脉络。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|