
{kind=link}
{kind=link}
{kind=link}
群落物种多度格局的分形解析
[苏强 ]
]
]
|
|
作者简介:苏强(1979-),男,辽宁沈阳人,讲师,主要从事生物多样性、生态化学计量学研究.E-mail:sqiang@ucas.ac.cn
生物群落物种多度格局(Species Abundance Distribution, SAD)研究是理解物种多样性决定机制的关键。这个问题不仅关系到物种多样性的保护与恢复,更重要的是,它有助于理清生物群落中各个物种之间错综复杂的相互关系,以及物质与能量在群落中的分配方式。群落物种多度格局研究的核心内容是如何构建SAD生态模型。该类研究始于20世纪30年代,相对已有的SAD模型而言,分形模型对生物群落样本的要求简单,适用性较高,便于不同群落间的比较。鉴于此,从分形模型的起源、假设条件、构建方法、分形参数的生态意义以及存在的问题等几个方面进行总结,以期促进此项工作的开展,为物种多度格局和多样性决定机制研究提供帮助。
The study of Apecies Abundance Distribution (SAD) is the key of understanding what determines species diversity. The theoretical exploration of SAD relates to the maintenance and conservation of biodiversity, and more importantly, it is conducive to clarifying complicated relationship among species and the distribution of matter and energy in a community. The research on SAD began in the 1930s, and there are so many kinds of theoretical models of SAD that can fit actual data, such as geometric-series model, log-series model, log-normal model, broken-stick model and so on. However, despite this, these models are often restrictive in their hypotheses and difficult to fit by natural communities. Especially, there is not always a good fit to the community that only has a few species. Thus, Frontier firstly introduced a family of models termed the Zipf-Mandelbrot model. This model is unrestrictive and easily fit by different natural community. Accordingly, this paper reviewed its origin, hypothesis, construction, problems and ecological signification of parameters to promote the research of SAD and the determination of species diversity.
在Science创刊125周年之际, 公布了一批最具挑战性的科学问题。其中, “ 什么决定了物种的多样性?” 被认为是最重要的25个科学问题之一[1]。理论生态学的发展自进化论起, 已有150余年的历史。虽然已经拥有了很多经典理论, 如生态位理论和中性理论等[2~8], 但至今生态学家还无法准确解释生物群落物种多样性的决定机制[1], 相比物理学等基础学科在过去150年间的进展, 这着实是一个令生态学家感到非常尴尬却又无法回避的科学问题[9, 10]!
物种多样性的决定机制是认识生物群落结构和功能的基础, 也是保护生物学研究中至关重要的领域[5, 7, 8, 11~14]。其现实意义在于, 如果不了解物种多样性的决定机制, 就无法有效地保护生态群落的物种多样性[11]。理论意义在于, 它可以理清群落中各个物种之间错综复杂的相互关系, 如竞争、捕食以及共生等[15]; 更重要的是, 它还有助于认识环境中的物质与能量在自然群落中的分配方式[3, 4, 6, 7, 12~14]。可以说, 揭示物种多样性的决定机制, 是未来群落生态学理论发展的根本支点[14]。
理解物种多样性的决定机制, 关键在于解析群落物种多度格局(Species Abundance Distribution, SAD) [3, 7]。生态学中, 物种多度是指群落中某一物种的个体数量或种群密度, 用来度量该物种在群落中的优势度或稀有度; SAD是描述群落中各物种间多度关系的量化方法, 用来揭示群落的组织结构[7]。例如, 群落A由3个物种a、2个物种b和1个物种c组成; 群落B由4个物种a、1个物种b和1个物种c组成。那么, 群落A的SAD为3:2:1, 而群落B的为4:1:1。因此, 虽然群落A和B的物种数量(S)、总多度(群落中各物种多度之和, NT)以及物种组成(a, b, c)均相同, 但这是2个组织结构不同的生态群落。所以, SAD反映了群落组织结构的本质属性, 这是群落物种数量、种类组成和总多度等参数所不能代替的。
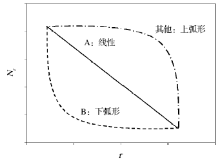
随着群落物种数量的增加, 以及种类组成的多样化, 上例中SAD的描述方法显然不足以有效地揭示群落组织结构的本质属性和构成原因[14~17]。对此, 有学者将群落中的各物种按其多度由大到小排序, 每个物种所对应的点由横坐标的排序位数(r)和纵坐标的多度(Nr)进行标记[18]。那么该散点图的递减方式, 则会揭示SAD的重要信息[19]。如上例所示, 群落A以线性方式递减(linear); 群落B以下弧形递减(concave); 当然, 某些群落也可能以上弧形递减(convex)等(图1)。因此, 如果自然群落的SAD确实遵循着某种普适性规律, 那么这种递减方式就有可能通过某种广义函数进行描述[18, 19]。如果能够找到这个函数关系式(模型), 就可以推导出不同群落组织结构的一般性法则, 进而解释物种多样性的决定机制[10, 20]。
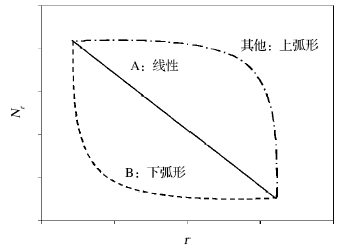 | 图1 生物群落物种多度随排序位数的递减方式Fig.1 The monotonically decreasing pattern of a community ordered in species abundance |
对此, 在过去近百年间生态学家提出了许多细致独特的生态模型, 如断棍模型、几何级数模型、对数级数模型以及对数正态模型等[6~8, 11~13]。基于这些模型, 生态学理论也得到了长足地发展, 可谓经典生态学的黄金时代。对于该阶段SAD模型及生态学理论上所取得的成就, 已有多位知名学者进行过总结[7, 21~23], 本文不再赘述。需要强调的是, 很多SAD模型的假设条件以及对数据资料的要求都比较苛刻, 实际上很难符合[24]。特别是, 对于只有几个物种所组成的群落, 大多数SAD模型都很难适用[19]。对此, Frontier根据分形假设, 最早引入Zipf-Mandelbrot模型来解释SAD[25]。该模型对群落样本的要求简单, 适用性较高, 便于不同群落间的比较。鉴于此, 本文从Zipf-Mandelbrot分形模型的起源、假设条件、构建及推演方法、分形参数的生态意义以及存在的问题等几个方面进行总结, 以期促进此项工作的开展, 为物种多度格局和多样性决定机制研究提供帮助。
Zipf-Mandelbrot分形模型的前身是广义化的Zipf法则, 该法则由Zipf教授基于语言学研究而提出[26]。但遗憾的是, 当时Zipf法则只是一种经验公式, 缺乏明确的数学依据[27, 28]。此后, Mandelbrot根据由其建立的分形几何学, 给出了广义化Zipf法则的数学推演过程, 并最终形成了Zipf-Mandelbrot分形模型[29]。因此, Zipf-Mandelbrot模型最早应用于语言学、信息学和社会经济等研究领域, 而后由Frontier引入群落生态学研究中[7]。但无论是在哪个领域, Zipf-Mandelbrot模型和广义化的Zipf法则的假设基础都是分形理论[27]。
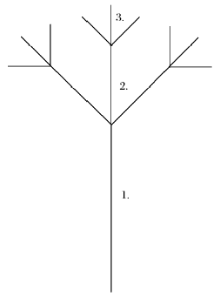
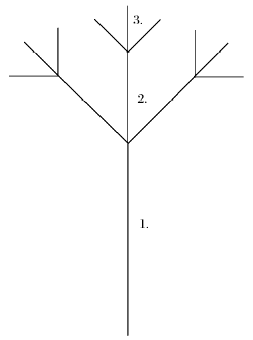
任何具有分形结构的研究对象, 都是由若干个的分形等级所构成; 在每个分形等级内, 又是由相同体量(size)、不同数量(number)的自相似单元组成; 随着分形等级的延伸, 某个等级内自相似单元的数量相对于前等级每增加K倍, 其体量减小k倍, 并且存在K=kd(d是分形维数)。以上就是Zipf-Mandelbrot模型的分形假设[30]。以Frontier给出的分形树为例(图 2), 分形等级2相对于等级1, 自相似单元的数量增加3倍, 分支大小减少2倍; 分形等级3相对于等级2, 自相似单元同样数量增加3倍、分支大小减少2倍; 以此类推。至第三个分形等级时, 共有自相似单元13个, 分形维数d=1.585[30]。
在SAD研究中, Frontier根据分形假设提出, 群落物种多度的排序位数(r)每增加K倍, 该等级上的物种多度减小k倍, 存在K=kd(d是分形维数) [30]。这样, 在第i个分形等级上(i=1, 2, 3, …), 新增物种数量Ri (R1=1, R2=K, R3=K2, …)与该等级上物种的多度Ni (由常数A开始, N1=A.k-1, N2= A.k-2, N3= A.k-3, …)之间存在量化关系(表1)。
| 表1 在第i个分形等级上, 新增物种数量Ri以及该等级上物种的多度Ni如表所示[30] Table 1 At the i-th fractal step, numbers of new species (Ri) and its relative abundance (Ni) during ecological succession is shown in the following table [30] |
如表1所示, 至第i个分形等级时, 群落物种数量的排序为数(ri)可表示为:
ri=R1+R2+R3+• • • • +Ri=1+K+K2+• • • • +Ki-1=(Ki-1)/(K-1)
第i个等级上物种多度, Ni=A.k-i; 由于K=kd, 设β =1/(K-1), γ =1/d, 则:
Ni=β r∙ A∙ (ri+β )-γ
由分形假设可知, 第i个等级上的物种多度相同, 所以Ni=Nr(Nr是排序为数在第r位上的物种多度)。另设A=N0β -γ , 可得:
Nr=No∙ (ri+β )-γ
上式即为SAD研究中的Zipf-Mandelbrot分形模型[7, 19, 21, 22]。在实际应用中, Nr可由该物种的相对多度(Fr=Nr/NT)代替; N0由F0代替, F0为所有频数之和; r为群落物种多度的排序位数, r=1, 2, 3, ... S; γ =1/d, d是该群落SAD的分形维数; β =1/(K-1), K是各个分形等级间物种数量的新增倍数[19, 25, 30]。可以看出, Zipf-Mandelbrot模型对群落样本的数据要求(仅需各个物种多度和物种数量), 以及假设条件(分形假设)都比较简单, 适用于不同群落之间SAD的比较。
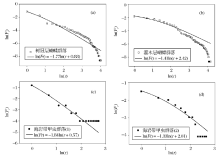
对Zipf-Mandelbrot模型进行双对数转换, 可以根据图形化的方法对横坐标上的ln(r)和纵坐标上的ln(Fr)进行拟合, 获得参数β 和γ [19]。Frontier在引入Zipf-Mandelbrot模型时, 曾经给出多个群落样本的拟合结果。这些结果几乎涵盖了所有类型的生态群落, 包括昆虫、浮游动物、浮游植物以及鱼类等[25]。本文引用了DeVries与Walla对热带雨林蝴蝶群落[31], 以及Kontkanend对海岸带甲虫群落的调查资料[32], 来验证Zipf-Mandelbrot模型在SAD研究中的适用性。对树冠层(canopy)和灌木层(understory)中蝴蝶群落的SAD进行双对数转换后, Zipf-Mandelbrot模型的拟合效果较好(图3)。树冠层蝴蝶群落的分形参数β 和γ 分别为0.88和1.77(图3a); 灌木层蝴蝶群落的分形参数分别为2.42和1.41(图3b)。同样, Zipf-Mandelbrot模型对甲虫群落的拟合效果也比较好。甲虫群落1(图3c)和2(图3d)的分形参数(β 和γ )分别为0.57和1.84, 以及2.01和1.33。
Zipf-Mandelbrot模型的分形假设, 有2种互不矛盾的解释。这2点分形解析最先由Mandelbrot在其博士论文中给出, 之后出现在其撰写的《自然界的分形几何学》一书中[30]。在生态学中, 这2点分形解释可以理解为:①物种的生态意义(cost of a species in an ecosystem), 即某个物种多度(或相对多度)越低, 则该物种在群落中出现的可能性越小。这说明现有环境中缺少某种珍贵的“ 资源” , 难以满足该物种的生长繁殖。因此, 稀有种生态学的指示意义要高于优势种。②物种出现的先决条件(previous conditions allowing the presence of a species), 即某种生物的出现必然对外界环境产生调节作用, 这种调节作用催生了新的生存环境, 并引入与之相适应的新物种。因此, 群落中每个物种都是遵循着某种特定的顺序依次出现的[19, 25, 30]。
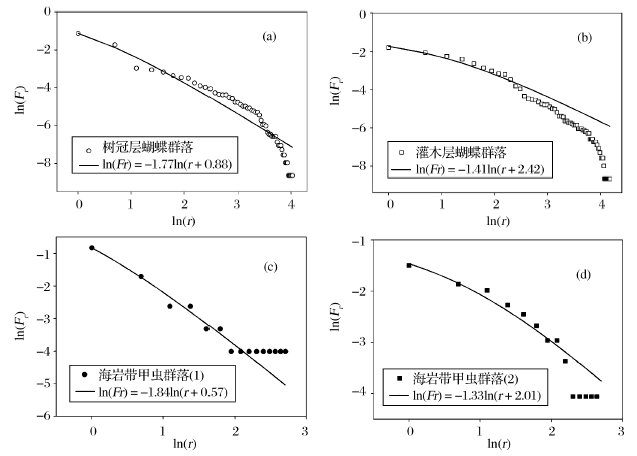 | 图3 Zipf-Mandelbrot模型对蝴蝶和甲虫群落的拟合结果Fig. 3 The fit results of Zipf-Mandelbrot model to SADs of butterflies and beetles communities |
基于此, Frontier认为分形参数γ 决定了某个物种在群落中出现的可能性, 即促使该物种出现在群落中的先决条件是否能够满足; β 决定了这种先决条件所表现出的可能类型, 即潜在的环境多样性[19]。具体而言, γ 越小, 随排序位数的增加, 物种多度下降的幅度越小, 群落物种组织结构更加均衡; 反之亦然。β 越大, 说明群落的均匀度及多样性指数越高; 反之, 则说明优势种的优势度越强, 均匀度和多样性指数越低[29]。这样, 如上例所示, 灌木层蝴蝶群落(图3b)相对于树冠层(图3a)而言, β 更大, γ 更小。所以, 灌木层群落的多样性高于树冠层。同样, 甲虫群落1(图3c)的分形参数比甲虫群落2(图3d) β 更小而γ 更大, 因此甲虫群落1的物种多样性较低。
由此可见, 一方面, Zipf-Mandelbrot模型的拟合结果能够较为准确地反应实际生态群落的组织结构(图3); 但另一方面, 它忽略了生物群落内客观存在的复杂性关系, 例如营养动力学过程、交互作用、优势种的特殊性以及物种间的反馈机制[33, 34]。因此, 虽然在Zipf-Mandelbrot模型中分形参数的生态意义简单而明确, 但是这种过于简化的生态学模型也使得它的研究价值和实际应用很难在经典生态学中得到确立[19]。所以, Zipf-Mandelbrot模型可以较好地对生物群落进行一种静态的刻画, 这是它的优势; 但它不能系统地阐明生物群落的这种存在状态是通过怎样的自然过程(或者机制)演变而来, 这是该模型的短板。
从Zipf-Mandelbrot模型对蝴蝶群落的拟合结果表明, 其所获得参数(β )与理论分形树有所不同。由于β =1/(K-1), 根据蝴蝶群落的拟合结果可得, 树冠层蝴蝶群落样本的K=2.14, 灌木层的K=1.41。这意味着, 在蝴蝶群落中, 排序位数在各个分形等级上的增加倍数为2.14(或者1.41)。这样, 在理论分形树内的各个分型等级上, 自相似单元以整数倍增加; 而在实际应用中, 增加倍数却为非整数。以树冠层蝴蝶群落为例, 分型等级1的物种数量为1, 等级2上为2.14, 等级3上为4.58, 以此类推。但实际上, 排序位数只能为整数, 这是Frontier在引入Zipf-Mandelbrot模型时忽略的一个问题。
对此, Mouillot等在Zipf-Mandelbrot模型的基础之上引入了随即变量, 使得排序位数的增加倍数等于Int(K)或者Int(K)+1[24]。根据Mouillot等提出的分形模型, 由于K大多为非整数, 所以分形群落中第i个物种有存在3种可能性:①属于第j个分形等级上的首个物种; ②属于第j个等级上的最后一个物种; ③属于第j个等级上的中间物种。这3种可能性出现的概率均与K有关, 依次为:
P(Xij)=P(Int(K(j-1) +1))+(Int(K(j-1))+1-K(j-1))P(Int(K(j-1)))
P(Xij)=(Kj-Int(Kj)P(Int(Kj)))
P(Xij)=P(X(i-1)j)
实际上, Mouillot等的分形模型主要强调, 群落中某个物种的多度取决于该物种所属的分形等级, 这与Zipf-Mandelbrot模型是一致的; 区别在于, Mouillot分形模型是通过K计算某个物种所属某个分形等级的概率, 进而估算该物种的多度。另外, Mouillot模型是一种随机性模型, 通过引入随机变量确保了排序位数必须为整数的基本要求, 这与Zipf-Mandelbrot模型是存在本质区别的[19, 24]。
SAD研究始于20世纪30年代, 其间涌现出大量与实测资料相符合的理论模型[7, 10, 14, 15, 21~23, 35]。Zipf-Mandelbrot分形模型只是McGill等总结的5类理论模型中, 分支过程模型里的一种[22]。相比其他理论模型而言, SAD的分形模型还存在着许多问题。其中, 最主要的有2点:①机制问题。如Tokeshi所言, 分形模型的生物学意义并不比对数级数和对数正态模型更加明确[21]。实际上, 最关键的问题是群落物种多度的分形等级。这种抽象的概念, 在实际的生态群落中难以找到明确的例证。另外, 分形模型还缺少类似于生态位理论或中性理论那样比较明确的生物学机制, 例如死亡率、迁出率、生态位等[2, 3, 6]。这些都使得该类模型尚未得到生态学家的普遍重视。②应用问题。也许正是由于分形模型在生态学机制上的含糊不清, 使得该类模型的推广、验证以及比较等研究工作, 还未能在更大范围内得以实现。特别是在Tokeshi的“ 批判” 以及Halley等[21, 36]的告诫(分形理论在生态学中的滥用)之后, 更使得人们对分形模型(Zipf-Mandelbrot或者Mouillot模型)在SAD研究中的应用变得十分谨慎。
但是, “ 任何可以描述SAD的方法都是值得探讨的, 无论这种方法是否依据了某些确切的群落生态学过程” [21]。所以, 鼓励并尝试着(而不是急于否定)从多个角度解析群落多度格局, 是目前解决多样性决定性机制问题的关键。在这方面, 以物种个体生态学的等价性为基点的中性理论, 就是一个很好的例证[37]。这种颠覆性的假设条件在群落生态学研究中的大胆尝试, 无疑极大地促进了生态学理论的发展[3, 6]。相比而言, 分形理论在物种多度格局研究中的应用还有待深入和完善。如Frontier所言, “ 分形理论在阐析生态学问题上的应用还非常有限, 但是其未来的前景绝非微不足道!” [19]。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|