{kind=link}
河流溶解硅的生物地球化学循环研究综述
[张乾柱1  , 陶贞
, 陶贞1, * , 高全洲1, 2 , 马赞文1 ]
, 陶贞, 高全洲|
|


作者简介:张乾柱(1989-), 男, 山东菏泽人, 博士研究生, 主要从事河流硅循环与全球变化研究.E-mail:qianzhuzhang@163.com
河流溶解硅(DSi)承载着陆地表生过程的环境信息, 其输入、迁移、转化和输出受多种因素制约。在全球硅酸盐岩风化过程中, 31.53%~64.87%的DSi被陆地植被吸收, 仅12.91%迁移至河流, 在向海洋输送过程中, 河流DSi又受到水生生物吸收、逆风化作用及“人造湖效应”等因素的影响, 输出量进一步减少, 弱化了海洋系统的“生物泵”作用;不多的研究表明全球河流DSi浓度变化介于138~218 μmol/L之间, 空间差异显著, 有必要量化各影响因素的贡献, 建立多因素控制的河流DSi输出通量模型;与地壳主要硅酸盐岩的δ30Si值(约为-0.5‰)相比, 全球河流DSi的δ30Si值变化范围较大(介于-0.2‰~3.4‰之间)且显著正偏, 分馏系数达0.3‰~3.9‰。这是由于流域内Si同位素的无机分馏和有机分馏2种动力分馏过程所导致。因此, 探讨河流DSi来源、迁移及转化机制是未来深入研究河流DSi循环的关键问题。
The riverine dissolved silicon (DSi) brings environmental information on biogeochemical processes of terrestrial surface, of which the input, transferring, transformation and output are influenced by many factors. Among the weathering of global silicate rocks, 31.53%~64.87% of DSi are intercepted by terrestrial vegetation and only about 12.9% are transferred into rivers. During being transported into ocean, riverine DSi gets impacts from aquatic biological absorption, reverse weathering process and artificial lake effect. The quantity of output is further reduced, which weakens the effect of the oceanic biological pump. According to limited data, the DSi concentration of global rivers has a large variation, ranging from 138 μmol/L to 218μmol/L. It is necessary to quantify contribution rates of influencing factors and establish output models controlled by multiple factors. The δ30Si of riverine DSi ranges from -0.2‰ to 3.4‰. Comparing with the δ30Si of silicate rock, which is about -0.5‰, the fractionation factor is significantly partial to positive from 0.3‰ to 3.9‰. That is because of the occurrence of kinetic fractionation process in river basin including inorganic and organic fractionation. Thus, the key problems, sources and transformation mechanisms of riverine DSi during migration and being transported should be solved in future.
硅(Si)是地壳中丰度(28.8%)仅次于氧(O)的第二种元素[1], 是陆生植物和水生生物的重要营养物质[2~4]。地表溶解硅(DSi)源自硅酸盐矿物的化学风化过程, 这一过程同时消耗大气CO2。全球硅酸盐矿物化学风化的固碳通量是8.67× 1012mol/a, DSi释放通量为5.66× 1012 mol/a[5]。藻类生长过程既固定大气碳(C), 也吸收DSi[6]。全球海洋表层硅藻光合固定的C和Si通量分别为2.15× 1015 mol/a与0.24× 1015 mol/a[7]。可见, 地球C循环与Si循环之间存在密切的耦合关系[5, 8~10], 同时河流DSi的输出是陆地生态系统和海洋生态系统碳汇作用的枢纽。
已往Si循环的研究多集中在陆地生态系统[8, 11~14]和海洋生态系统[7, 15~17]。硅酸盐矿物风化释放的DSi, 一部分被植物吸收并转化为生物硅(BSi), 然后通过枯枝落叶方式返回至土壤, 经微生物分解重新释放DSi。因此, 经陆地生态系统迁移至河流的DSi来源于岩石风化释放和植物BSi分解。尤其在森林生态系统中, 岩石风化成因的DSi所反映的环境信息, 在很大程度上已被BSi所改变[11]。
目前对河流DSi的来源、迁移及转化过程缺乏较深入的研究[18]。随着河流富营养化和湖泊效应的加强[19, 20, 21, 22, 23], 河流中Si循环的信息更加趋于复杂。因此, 深入研究地球表层DSi在陆地生态系统和河流生态系统中的迁移、转化过程以及识别流域DSi输出通量的影响因素, 是研究地球表层Si循环的关键。
硅在地球表层不同介质中有不同的存在形态:土壤中Si主要存在于晶质的原生硅酸盐矿物和次生黏土矿物(蒙脱石、高岭石、绿泥石和伊利石等)中, 部分Si存在于隐晶质— 非晶质矿物(如蛋白石、玉髓、水铝英石等)和非晶质的植物硅酸体[24]中, 还有以偏硅酸(H2SiO3)和[SiO3]2-离子存在的DSi; 植物体内的Si以无定形硅(SiO2· nH2O)的形式驻留在植物组织中形成稳定的植物硅酸体[25]; Si在水体中以DSi的形式存在。一般情况下, 地球表层的Si以DSi形式在岩石圈、土壤圈、生物圈以及水圈之间迁移、转化。
硅酸盐矿物风化产生的DSi进入土壤溶液后, 一部分被陆地植物吸收并转化为BSi, 一部分以次生含硅矿物的形式析出[26], 剩余的随径流迁移进入河流。植物体内的BSi随枯枝落叶返回至土壤中。进入土壤的BSi, 一部分被埋藏, 另外一部分被重新分解释放DSi。陆地生态系统中土壤BSi成因的DSi与岩石风化DSi一起参与上述Si循环过程(图1)。由以上陆地生态系统Si循环可知, 土壤DSi包括硅酸盐矿物风化释放和BSi再分解两个来源, 可以反映“ 岩石— 土壤— 植物” 陆地内循环的综合信息。
河流Si主要包括未被风化的硅酸盐矿物颗粒(RSi)、土壤DSi及陆地植物BSi 3种形式。随机械侵蚀进入河流的硅酸盐矿物颗粒RSi和陆地植物BSi, 一部分沉积并埋藏在河床, 另一部分被分解释放DSi。上述分解释放的DSi与径流直接输入的DSi组成了河流DSi。河流DSi, 一方面被水生生物吸收、形成水生生物BSi, 另一方面在饱和状态下析出(如黏土矿物和蛋白石)。同样, 水生藻类形成的水生生物BSi, 可以被埋藏或分解释放DSi, 参与河流生态系统Si循环。沉积在河床的次生硅酸盐矿物、蛋白石和BSi, 在紊流搅动下, 重新分解释放DSi。因此, 河流DSi反映外源输入和内生循环过程的综合信息。
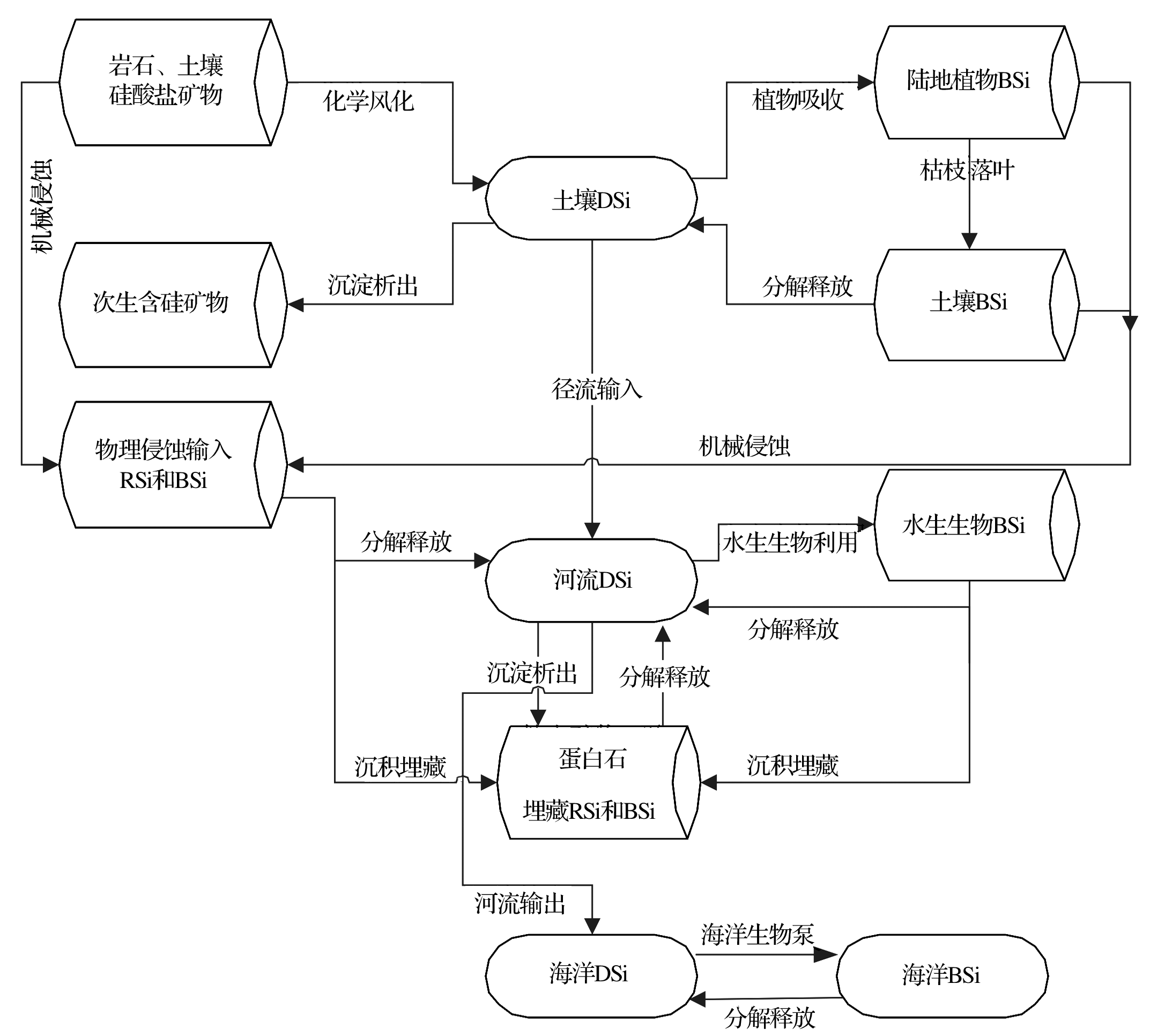 | 图1 地球表层DSi的循环Fig.1 Dissolved silicon cycle of the earth’ s surface |
地球表面的DSi源自硅酸盐矿物风化, 但这一过程释放的DSi通量远低于全球陆地植物吸收通量(60× 1012~200× 1012mol/a[13]), 揭示了土壤中DSi的另一来源。植物生长过程中形成的BSi通过枯枝落叶凋落返还至土壤, 后经分解作用释放DSi, 构成土壤DSi的重要部分。Bartoli等[11]研究表明, 不同植物群落返还给土壤的BSi量不同:落叶林群落枯枝落叶返还给土壤的BSi量占年形成量的85%, 针叶林群落稍低, 达到63%。不同群落中土壤BSi分解释放DSi的强度也存在差异:落叶林系统中Si的生物循环较快, 约85%的土壤DSi来源于陆地植物BSi的释放, 岩石化学风化对其贡献较小; 而在针叶林系统中, 约85%的土壤DSi来源于矿物风化释放, BSi对土壤DSi的贡献仅有15%[11]; 热带雨林系统中BSi对土壤DSi的贡献是硅酸盐矿物风化释放量的2倍[8]。
陆地植物和土壤之间的Si循环延长了Si在陆地生态系统中的滞留时间, 减缓了其向河流迁移的速率, 减少了Si自陆地系统向系统外的流失量, 进而影响陆地(生态)系统Si的表观输出通量。受植物生长差异的影响, 不同陆地生态系统Si向系统外的流失量有所差异, 亚马逊雨林系统Si流失量为183 mol/(hm2· a)[11], 针叶林系统达(433± 117)mol/(hm2· a)[27], 竹林群落高达583 mol/(hm2· a)[28]。
此外, 流域内植物生长差异, 也是导致冰期— 间冰期之间河流DSi通量变化的重要因素。尽管冰期时陆地岩石风化作用较弱, 但河流DSi通量却为间冰期的2倍[15], 其主要原因为间冰期陆地植物生长旺盛、DSi吸收量大。陆地植物生长吸收大量的DSi, 形成BSi储存在土壤中, 抑制了陆地DSi向河流、海洋的迁移[28]。
由于流域内陆地植物的生长吸收, 硅酸盐岩风化释放的DSi不能全部迁移至河流。我们把迁移至河流中的DSi量占总释放量的百分比称为“ 迁移率” , 在迁移过程中的损失量所占的比重, 称为“ 损失率” 。在硅酸盐岩和碳酸盐岩为主导的流域中, 河流中离子来源不同, 河流DSi迁移率与损失率的研究方法也不同。硅酸盐岩流域内, 若不考虑Ca在次生硅酸盐矿物沉淀过程中的流失, 根据河流与基岩中Ca/Si值(经大气沉降校正后), 可计算DSi自岩石风化到河流的迁移率与损失率(公式1)[29]:
据公式(1)计算得出, 在冰岛地区岩石风化释放的DSi中, 约(51± 12)%在迁移过程中由于次生黏土矿物的形成而丢失[29]。
在有碳酸盐岩分布的流域内, 河流中大部分Ca2+源于碳酸盐岩风化, 因此公式(1)并不适用。河流中Na+, K+主要源于大气沉降、蒸发盐岩溶解和硅酸盐岩风化。扣除前2个过程的贡献, 硅酸盐岩风化则为河流中Na+, K+的主要来源。与Si相比, Na+, K+在次生黏土矿物中滞留量较少。不同次生黏土矿物中Si含量不同, Si/(Na + K)比值随Si在次生黏土矿物中含量增多而降低。根据Si/(Na + K)比值在基岩与河流之间的差异, 可估算DSi的迁移率(公式2)[30]:
式中, (Na+K)* 为扣除大气沉降、蒸发盐岩溶解后来自硅酸盐岩风化部分
陆地硅酸盐岩流域多以花岗岩为主, 占地表火成岩面积的20%~25%[31]。因此, 根据全球硅酸盐岩地区河流离子浓度的平均值和花岗岩的元素含量, 按公式(1)和(2)分别计算河流DSi迁移率(表1)。2种方法计算结果相近, 分别是11.67%和14.14%, 为了平衡2种方法带来的误差, 取两者均值12.91%为全球河流DSi平均迁移率。根据风化产物中含Si量的差异, 将岩石脱硅富铝化过程大致分成3个阶段:双硅铝化、单硅铝化及铝铁化, 河流Si/(Na + K)值分别对应2, 2~3.5及3.5[30], 以此计算出的DSi迁移率为44.44%~77.78%, 远高于上述迁移率均值12.91%, 其中损失部分(31.53%~64.87%)被陆地植被吸收利用。
| 表1 全球河流DSi迁移率估算 Table1 Calculation of migration rate of DSi in global rivers |
水体DSi是浮游硅藻的主要营养物质之一。地质历史时期水体DSi浓度和硅藻是调节全球气候变化的重要介质。生物出现之前, 水体DSi含量主要受岩石风化和次生硅酸盐矿物的形成控制。硅藻等水生生物出现以后, 大量消耗DSi, 尤其三叠纪— 白垩纪硅藻的爆发式增长, 导致DSi浓度剧烈降低[32], 同时, 大气中CO2被大量吸收, 使得气候变冷。当DSi浓度降低到一定程度时, 硅藻生长受到抑制, 光合固碳速率降低, 气候则逐渐回暖[32, 33, 34, 35]。现代环境条件下, 河流水生藻类在DSi迁移过程中将DSi转化为BSi, 减少了河流DSi的入海通量。据统计, 全球地表河流BSi浓度为28 μ mol/L, 约(1.05± 0.2)× 1012mol/a(占全球河流DSi通量的17%[36])经河流搬运入海[37]。受硅藻生长的季节性差异影响, 河流中BSi和DSi的浓度同样存在季节变化。一般情况下, 温带地区河流、湖泊中春季硅藻量最高[38]。硅藻生长旺盛时期, 大量的河流DSi被吸收并转化为BSi, 此时河流BSi含量占Si(DSi+BSi)总量的50%~70%, 非生长旺盛期BSi含量仅为10%~20%[37]。在硅藻生长旺盛期, 河流Si (DSi+BSi)含量比非旺盛期少得多, 相差近40%以上[37, 39]。上述现象揭示了在硅藻生长旺盛时期河流DSi被转化为BSi, 沉积并埋藏在河床。
人类活动对河流DSi浓度和通量的影响主要通过以下方式实现:改变土地利用方式、促使水体富营养化及修筑拦水大坝等。
自陆地生态系统DSi迁移至河流过程中, 陆地植被的吸收是影响迁移率的重要因素。流域下垫面性质稳定时, 河流DSi通量的季节波动, 反映了陆地植物生长和吸收的季节差异[40]。土地利用方式的更替往往带来植被吸收量的变化, 从而引起流域DSi输出通量的改变[13, 40~42]。Carey等[43]研究得出:河流DSi通量和流域植被覆盖度呈负相关; 植物的季节性生长吸收差异导致河流DSi通量波动幅度达40%。流域植被受到破坏之初, 河流DSi通量显著升高。之后, 河流DSi通量的变化趋势和后来的土地利用方式密切相关。若流域植被自然恢复, 河流DSi通量趋向变低; 若被城市建设用地替代, DSi通量趋向更高; 若用于耕地, 河流DSi通量比自然恢复的流域更为偏低[43]。
富含大量氮、磷、钾等营养元素的工业和生活废水造成水体富营养化, 导致包括硅藻在内的水生生物加速生长[19, 20, 44], 使大量河流DSi被吸收、转化为BSi并埋藏在河床[20, 45], 引起河流DSi浓度显著降低。反之, 当水体富营养化程度减弱时, 河流DSi浓度出现升高趋势。例如由于环境条件的改善, 20世纪90年代莱茵河NO3-与PO43-的浓度约减少了1/3, 河流DSi浓度增加了70%[21]。
流域内拦水大坝的修筑改变了河流水文情势, 使水生藻类大量繁殖[46~48], 导致河流DSi转化为BSi的机率升高。由于库区水流流速减慢, BSi被大量埋藏在库区[23, 49, 50], 河流DSi浓度降低, 学者称该现象为“ 人造湖效应” [22, 51]。例如流域性质相似的欧洲三条河流, 受人工筑坝影响显著的Vistula和Daugava河DSi平均浓度仅为Oder河的15%~50%[52]; 肯尼亚Tana河中, 41%的DSi 滞留在Masinga坝库区[53]; 修筑于亚洲第一大河的长江三峡大坝, 拦截了44%的河流BSi[54]。
海洋中HCO3-、可溶性硅 (DSi与BSi)及碱性阳离子(Cations)和悬浮硅酸盐矿物颗粒(Silicate Mineral)发生反应, 生成富含碱性阳离子的硅酸盐矿物(Cations Silicate Mineral), 同时向大气释放CO2[55]:
其中, 部分HCO3-, DSi及碱性阳离子(Na+, Mg2+, Ca2+及K+)是硅酸盐矿物风化的产物。在上述反应中, 风化产物重新合成新的硅酸盐矿物, 是风化作用的相反过程, 故称之为“ 逆风化作用” 。上述逆风化过程, 使得2亿年以来海水成分基本稳定[55], 同时是调节海水pH和大气CO2浓度的重要机制[56]。
室内模拟实验发现, 由于流域风化物质的输入和大量BSi的分解, 河口区成为逆风化过程进行的重点区域[57]。这一过程直接减少DSi的入海通量, 例如约22%的河流DSi被固定在亚马逊河口区[58], 密西西比河口区更是高达40% [59]。
由于水生生物吸收、营养元素排放及人工筑坝等因素的影响, 一部分河流DSi自上游向下游迁移过程中, 以BSi形式沉积并埋藏在河床, 导致河流DSi入海通量降低。在过去100年间, 水体富营养化和“ 人造湖效应” 使得注入波罗的海的河流DSi通量减少了30%~40%[60]。自20世纪70年代以来, 多瑙河向黑海输送的DSi通量降低了2/3[61]。基于Jansen[62]和Moosdorf[63]估算的北美地区单位流域面积DSi迁移量, Lauerwald推算出了河流DSi滞留率公式:
式中, Fdrainage为流域输出量; Ft为总输入量; Siloss和rDSi为陆地迁移损失率和河流运移滞留率; fMDSi为DSi入河流通量; fCDSi为河流DSi的入海通量。
根据公式(4)计算得出:在北美地区, 每年约2.3× 106mol DSi滞留在流域内, 滞留率达13%; 单位湖泊面积引起的DSi滞留量为72 mmol/(m2∙ a)[64]。
目前全球河流系统中, 大型湖泊造成的河流DSi滞留量占全球河流DSi通量的18%~19%[65]。小型水库因水体滞留时间短, 水库上、下游DSi浓度无明显差异, 对河流DSi消减作用不明显, 但对河流BSi拦截作用明显[52]。
Clarke[66]根据欧洲及北美洲温带地区河流DSi含量, 估算出全球DSi浓度平均值为138 μ mol/L; 在Clarke数据基础上, Livingstone[67]计算得出全球DSi浓度平均值为218 μ mol/L。基于全球60条河流DSi浓度及流量, Meybeck[68]给出了全球DSi浓度平均值为173 μ mol/L。随着全球河流水化学监测工作的大规模开展, Tré guer[7]基于全球250条河流, 将全球分为9个构造类型区, 推算全球河流DSi浓度平均值为152 μ mol/L。Meybeck[69]将内流河考虑在内, 计算得出全球DSi均值为146 μ mol/L。根据中小尺度流域建立的经验模型参数, 同时考虑流域流量、降雨量、温度及岩性等因素, Beusen[65]估算的全球DSi均值为162 μ mol/L。后来Dü rr等[36]基于全球径流量60%的河流、140个流域片区, 在加权均值基础上, 估算出河流DSi均值为158 μ mol/L, 从而推算出世界河流入海DSi通量为(6.2± 1.8)× 1012mol/a[17]。综合上述研究结果, 全球河流DSi浓度变化于138~218 μ mol/L之间, 平均值为(161± 24) μ mol/L。
按照全球河流DSi通量标准化值, 即YDSi/YDsi(YDSi为任一河流DSi通量; YDsi为全球河流DSi通量均值), 全球流域分为热岩区、强活动区、多活动区、中等活动区、少活动区、弱活动区、微弱活动区及无活动区8种类型[17, 36]。其中, 强活动区主要出现在环太平洋地区, 与构造活动一致, 北冰洋地区多为弱活动、无活动区。在注入四大洋的外流河中, 注入太平洋的河流DSi浓度最高(387 μ mol/L), 其次为注入印度洋的河流(200 μ mol/L), 第三为大西洋流域为140 μ mol/L, 接近全球河流平均值, 北冰洋地区河流最低, 仅为102 μ mol/L。然而, 大西洋却接收了全球河流DSi通量的45.6%, 太平洋、印度洋与北冰洋分别接纳了32.3%, 18.6%及5.4%。这可能是由于大西洋区域存在亚马逊河和刚果河等大河, 河流年径流量达全球总量的51%, 为太平洋地区河流径流量的1.88倍[36]。
若将流域视为独立系统, 那么河流DSi的输出通量则为系统内“ 收支” 差值。系统内DSi输入源主要为岩石风化(Fweathering)和大气沉降(Fatmospheric), 系统内的支出为陆地植被吸收(Fvegetation)和水生生物利用(Faquatic), 据此建立流域DSi输出通量(Fdrainage)的经验公式:
考虑径流、岩性、温度、植被及坡度等要素和河流DSi通量之间的相关性, 建立影响机制模型, 能在一定程度上反映各要素对河流DSi通量的影响程度。例如, Jansen等在分析北美地区142个水文监测站资料和DSi数据基础上, 探讨流域各自然要素与河流DSi通量之间的关系, 发现径流量和岩性与河流DSi相关性较强, 建立了径流和岩性对河流DSi通量的影响机制模型[62]:
式中, FDSi为河流DSi通量(tSiO2/(km2· a)), b0, b1~n为模型参数, Ln为不同岩性在流域所占面积, q为流域径流量。
模型(6)计算结果与实测值相关性(r=0.72)低于模型(7)(r=0.94)[62], 表明在通量模型中, 考虑多因素影响获得的预测效果优于单因素, 同时, 在一定程度上说明流量和岩性是影响河流DSi产量的重要因素。河流悬浮颗粒物(SPM)是岩石物理剥蚀程度的重要指标, 岩石物理剥蚀能为化学风化持续提供新鲜物质, 由此SPM与DSi输出通量理论上成正相关, 然而北美洲107个流域的SPM与DSi输出通量并未表现出上述关系。说明径流量与DSi通量之间的正相关关系, 并不能简单理解为径流增加(减少)对机械侵蚀的加强(减弱)作用[62]。不同岩性地区, 流域单位面积产Si量差异明显。辉长岩、辉绿岩及玄武岩等基性火成岩较易被风化, 因此流域产Si量最高。其次, 分别为中、酸性喷出岩(流纹岩、粗面岩等)、未固结成岩的松散堆积物、变质岩、碳酸盐岩、混合沉积岩及砂岩, 酸性侵入岩流域区产Si量最低[62]。
Si同位素研究始于矿物学、岩石学与矿床分布等领域[70]。关于河流DSi的稳定同位素组成(δ 30Si)研究源自De La Rocha等[71]。与其它含硅物质相比, 河流DSi更富集30Si。这是由于Si同位素分馏作用的结果。Si同位素分馏过程分为无机分馏和有机分馏过程, 无机分馏伴随着硅酸盐矿物的溶解与沉积, 有机分馏则由陆地植物和水生藻类的吸收作用导致。地壳主要硅酸盐岩δ 30Si值约为-0.5‰ , 风化过程中形成的黏土矿物富集28Si, 其δ 30Si值在-1.0~-1.9之间[72], 更多的30Si进入溶液中[71, 73], 风化形成的DSi与硅酸盐岩之间的分馏系数为0.5‰ ~1.4‰ 。另一方面, 在 “ 陆地-河流” 迁移过程中, 陆地植物优先吸收28Si, 使得河流中剩余DSi的δ 30Si值正偏[74, 75], 分馏系数约为1.0‰ [76~78]。Si同位素分馏现象在水生生态系统中也普遍存在:水中析出的沉积硅往往富集28Si, 例如蛋白石的δ 30Si值约为-3.4‰ ~0.2‰ , 与全球河流DSi的δ 30Si值相比, 明显负偏[72, 79], 分馏系数达0.4‰ ~6.8‰ ; 硅藻的吸收作用导致水中DSi的δ 30Si值偏重, 其分馏系数为1.0‰ [80~82]。
De La Rocha等[71]研究了美洲和非洲河流的δ 30Si特征, 随后研究者先后又在长江、黄河、亚马逊河等河流[29, 30, 53, 71, 76, 83]开展了DSi的δ 30Si研究(表2)。目前全球河流DSi的δ 30Si变化于-0.2‰ ~3.4‰ 之间, 河口区δ 30Si值变化于0.92‰ ~3.0‰ 之间(测试精度介于± 0.07‰ ~± 0.14‰ ), 与地壳主要硅酸盐岩(-0.5‰ )[72]相比, 分馏系数为0.3‰ ~3.9‰ 。
受岩石风化、陆地植物吸收、水生藻类利用以及沉积硅析出等动力分馏过程的影响, 河流自上游到河口DSi浓度出现逐渐降低且δ 30Si值趋向偏重[76]。
| 表2 世界部分河流的δ 30Si Tab.2 The δ 30Si of several rivers in the world |
河流δ 30Si在一定程度上反映了流域生物地球化学环境特征, 同时也是研究流域DSi迁移率的重要指标。在岩石风化元素迁移过程中, K元素比 Na元素更易于被植物吸收利用。陆地植物倾向于吸收利用28Si, 致使河流DSi相对富集30Si。因此, 河流DSi的δ 30Si与K/Na值呈反相关关系(公式8)[30]。
在地表陡峭、降雨多及径流量大的山区, 由于强烈的侵蚀作用, 基岩不断被暴露风化, 同时次生矿物形成受到抑制。流域DSi往往具有高迁移率、低滞留率及弱分馏作用, 河流DSi浓度高且δ 30Si值低, 河流δ 30Si值与DSi迁移率呈直线线性相关。在地表坡度和缓、降雨少及径流量小的地区, 岩石风化速率慢, DSi迁移率低、滞留率高, 分馏作用强, 河流DSi浓度低且δ 30Si值偏高, 河流δ 30Si值与DSi迁移率呈曲线线性相关[29]。
陆地生态系统中土壤DSi是 “ 岩石— 土壤— 植被” 系统Si循环的“ 桥梁” , 反映了岩石风化和陆地植物吸收作用的环境信息。陆地DSi进入河流系统后又承载了河流水文过程和水生生物活动(藻类生长)的信息。全球硅酸盐岩风化过程中, 31.53%~64.87%的DSi被陆地植物吸收, 仅12.91%迁移至河流。在向海洋输送过程中, 河流DSi又受到水生生物吸收、逆风化作用及“ 人造湖效应” 等因素的影响, 输出量进一步减少。
河流DSi是陆地DSi输入海洋的唯一通道。有限的研究表明全球河流DSi浓度变化范围(变化于138~218 μ mol/L之间)大, 空间差异显著。
与地壳主要硅酸盐岩的δ 30Si值(约为-0.5‰ )相比, 全球河流DSi的δ 30Si值变化范围较大(变化于-0.2‰ ~3.4‰ 之间)且显著正偏, 分馏系数达0.3‰ ~3.9‰ 。这是由流域内Si同位素的无机分馏和有机分馏两种动力分馏过程所导致。前者主要为硅酸盐矿物的溶解(分馏系数为0.5‰ ~1.4‰ )与沉积(分馏系数为0.4‰ ~6.8‰ ), 后者为陆地植物与水生藻类的吸收利用(分馏系数为1.0‰ )。
(1)河流DSi的来源、迁移和转化过程复杂、影响因素多。据研究, 河流DSi的来源除矿物风化源、BSi溶解源外, 人为活动(家庭洗涤、造纸工业及废水处理等)可以使河流DSi通量增加2%[85]。搬运过程中, 颗粒含硅物质(硅酸盐矿物、BSi)的分解输入, 在某种程度上可增加河流DSi浓度[86]。例如, 硅酸盐矿物风化释放DSi速率约6× 10-14mol (cm2∙ s)[87]; 河流BSi分解速率为2× 10-9mol (cm2∙ s)[88]。虽然陆生植物生长过程的吸收作用导致DSi迁移率降低, 但是根劈作用和有机酸的释放[89], 陆地植物能加快硅酸盐矿物的风化, 导致DSi加速释放, 然而何种状态下能达到平衡却不得而知。因此, 对河流DSi来源、迁移及转化机制的研究是未来深入研究河流DSi循环需要解决的关键问题。
(2)河流DSi变化受多因素控制, 精确测量全球陆地输向海洋的DSi通量不太现实。因此, 建立多因素控制的河流DSi输出通量模型, 量化各影响因素的贡献, 应成为将来河流DSi循环研究的热点。
(3)目前Si同位素示踪技术主要应用在海洋和陆地生态系统Si循环研究中, 河流DSi同位素研究相对薄弱。河流DSi的δ 30Si值变化可以示踪流域地表过程的强度, 因此, 河流DSi的同位素研究值得关注。
The authors have declared that no competing interests exist.
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|