{kind=link}
古-中生代之交双壳类演变研究进展*
[黄云飞1, 2  , 童金南
, 童金南2, * ]
, 童金南]
|
|


作者简介:黄云飞(1986-), 男, 河南林州人, 讲师, 主要从事二叠纪—三叠纪之交双壳类的灭绝与复苏方面的研究. E-mail:didahyf@163.com
古—中生代之交, 伴随着显生宙历史上最大的生物灭绝, 发生了最大的一次生态系结构变革, 软体动物双壳类取代了腕足类在底栖生态系中的主导地位。以中国南方为代表的特提斯地区晚二叠世末至早三叠世良好的地层记录和丰富的双壳类化石材料为该时期双壳类及底栖生态系的演化研究提供了良好条件。通过系统收集和整理全球范围内该时期各地区有关双壳类的研究资料及主要成果, 建立各个地区双壳类带的对比关系, 并对30年来的双壳类生物地层工作做了简要的总结。从分异度和生态特征等角度来说, 双壳类在二叠纪—三叠纪之交的生物危机中表现出了中等程度的灭绝, 并在经历了长时间的迟缓复苏后, 于早三叠世
The end-Permian mass extinction was the biggest catastrophe during the Phanerozoic history, which induced the great transition of marine ecosystem from articulated brachiopod-dominated Paleozoic Fauna to bivalves-dominated Mesozoic Fauna. The complete end-Permian to Lower Triassic successions with abundant bivalve fossils enhance our further understanding of the evolution of bivalves and benthic ecosystems during this period. The advances in the studies of bivalves from various areas including South China, Southern Alps, Pakistan and West USA, and the Early Triassic bivalve assemblages and evolution are reviewed and summarized. The
晚二叠世末期发生了显生宙历史上最大的一次生物灭绝[ 1, 2, 3], 造成了90%以上的海洋生物种灭绝[ 3]。关于这次灭绝的机制众说纷纭, 主要有火山作用与全球变暖[ 4, 5, 6, 7, 8]、海洋缺氧/硫化[ 9, 10, 11, 12, 13, 14, 15]等。而在生物的灭绝模式方面, 则主要有一幕式[ 16, 17]、两幕式[ 18, 19, 20, 21, 22, 23]等。软体动物双壳类在晚二叠世较为丰富, 在二叠纪—三叠纪之交(P-T)灭绝事件中遭受了中等程度的灭绝, 但在随后却成为灭绝后的海洋底栖生态系的主导生物[ 24, 25]。这次灭绝同样导致了显生宙历史上最大的一次生态系结构变革, 即古生代型生态系向中生代(现代)型生态系的转变, 其中底栖群落的主导分子由腕足类转变为了软体动物双壳类、腹足类等[ 26, 27, 28], 而这种转变则归因于当时占据浅水环境的腕足类的大规模灭绝、双壳类优势的生理特征使其更适应于浅水环境等[ 29, 30, 31, 32, 33, 34]。在晚二叠世末生物大灭绝之后, 出现了一段长时期的生物迟缓复苏[ 35, 36], 这被认为是由于持续波动高压的生态环境造成的[ 5, 36, 37, 38, 39, 40, 41, 42]。因而, 对晚二叠世到中三叠世这一关键时期分布最为广泛的软体动物双壳类的演化进行深入的研究, 可以更好的理解生物的灭绝—残存—复苏过程及其对生态环境的响应机制。
双壳类作为早三叠世一类分布广泛的宏体生物化石, 具有悠久的研究历史, 最早可追溯到18世纪末期至19世纪中期[ 43, 44, 45]。自19世纪末期到20世纪70年代, 阿尔卑斯地区[ 43, 44, 45, 46, 47, 48, 49]、俄罗斯远东[ 50, 51]、喜马拉雅地区[ 52, 53, 54, 55, 56]、华南地区[ 57, 58, 59, 60, 61, 62]、美国西部地区[ 63, 64, 65]、西澳大利亚[ 66]、格陵兰[ 67, 68]、日本[ 69, 70, 71, 72, 73, 74]、东南亚[ 75, 76]等地的早三叠世双壳类化石属种相继得到大量的报道, 奠定了该时期双壳类研究的基础。20世纪80年代以来, 二叠纪—三叠纪之交双壳类的研究逐步深入, 除属种报道[ 77, 78, 79, 80, 81, 82]外, 在生物地层[ 83, 84, 85, 86, 87]、灭绝—复苏模式[ 88, 89, 90, 91, 92, 93]、生态系演化[ 35, 94, 95, 96, 97, 98]等方面均取得了重要的研究进展。
阿尔卑斯地区包括意大利北部、奥地利南部等地, 古、中生代地层发育齐全, 下三叠统地层为Werfen组, 自下而上再分为Tesero, Mazzin, Andraz, Siusi, Gastropod Oolite, Campil, Val Badia, Cencenighe和San Lucano段, 其中P-T界线位于Tesero段中, 印度阶—奥伦尼克阶界线位于Gastropod Oolite段中部[ 99]。该地区自19世纪中期即有三叠纪地层和双壳类化石的研究报道, 研究程度较高[ 43, 44, 45, 46, 47, 48, 49]。20世纪80年代, Broglio Loriga团队在意大利北部地区(或称为南阿尔卑斯地区)开展了高精度的早三叠世双壳类生物地层工作, 建立了 Claraia wangi- C. griesbachi, C. clarai, C. aurita, Eumorphotis hinnitidea, Costatoria subrotunda, E. kittli, E. telleri, Costatoria costata等生物带[ 83, 84]。Posenato[ 99]分析了意大利北部地区早—中三叠世双壳类属分异度的变化, 建立了该地区的复苏模式, 认为双壳类到Anisian晚期才复苏。Twitchett[ 100]和Metcalfe等[ 101]对早三叠世双壳类 Claraia, Eumorphotis, Unionites等属的个体大小变化进行了初步探讨, 认为双壳类在晚二叠世大灭绝后表现出了小型化现象(Lilliput effect), 直到Griesbachian晚期和Dienerian期个体才逐渐增大。
俄罗斯远东地区下三叠统地层出露于South Primorye等地, 不整合覆盖于二叠纪地层之上, 划分为Lazurnaya Bay组和Zhitkov组。本区的Abrek Bay剖面曾被作为印度阶—奥伦尼克阶GSSP候选剖面之一, 已建立良好的菊石生物地层和牙形石生物地层格架[ 102]。19世纪末期至20世纪早期, Bittner[ 50]和Kiparisova[ 51]等先后报道了本区早三叠世的部分双壳类化石属种。近年来, Kumagae和Nakazawa[ 79]报道了Abrek Bay剖面的早三叠世双壳类化石。该地区的早三叠世化石也多为全球性分布属种, 如 Eumorphotis, Promyalina, Pteria, Neoschizodus, Unionites等。
喜马拉雅地区包括印度北部、巴基斯坦、尼泊尔等地, 其中以西喜马拉雅区的Kashmir, Zanskar, Spiti, Kumaon和中喜马拉雅区的Nepal研究历史较长。本区二叠纪—三叠纪之交双壳类研究的先驱是Bittner[ 53], 随后Diener[ 54]和Nakazawa[ 55, 56]先后报道了克什米尔地区的晚二叠世至早三叠世的双壳类化石, 自下而上建立了 Palaeolima middlemissi, Claraia bioni- Etheripecten haydeni, Eumorphotis venetiana- E. aff. bokharica, Claraia aff. griesbachi- E. multiformis, Claraia concentrica, Leptochondria minima和 Claraia decidens等7个生物带[ 56]。Wittenburg[ 52]最早报道了巴基斯坦Salt Range地区早三叠世双壳类化石7种, Wasmer等[ 81]近年来在巴基斯坦Salt Range和Surghar Range地区采集了大量的Smithian期和Spathian期的双壳类化石, 经鉴定得到11属15种, 其中10种未在其他地区报道过。
20世纪20~60年代, 华南地区零星的早三叠世双壳类化石不断得到报道[ 57, 58, 59, 60, 61], 随后的区域地质调查工作中, 出版了各门类、各地区的古生物图册, 如《中国的瓣鳃类化石》, 描述了大量的双壳类化石属种, 为随后的生物地层工作奠定了坚实的基础。80年代以来, 双壳类生物地层得到了极大的发展和应用, 既有单个剖面的双壳类生物建带工作[ 86, 103, 104, 105, 106], 也有地区性的生物地层归纳总结[ 85]。殷鸿福[ 85]对华南地区二叠纪—三叠纪之交的双壳类生物地层进行了总结, 建立了 Hunanopecten exilis, Towapteria scythica- Pteria ussurica variabilis, Claraia wangi, Eumorphotis multiformis- Claraia aurita, Eumorphotis inaequicostata- Pteria cf. murchisoni等生物带。殷鸿福[ 85, 89]、李玲[ 90]、方宗杰[ 92]等依据华南地区丰富的双壳类化石数据, 探讨了二叠纪—三叠纪之交双壳类的灭绝模式, 其灭绝模式被认为是单幕式[ 85, 89, 90]或双幕式[ 92]。华南地区早—中三叠世双壳类的属级分异度演化, 表明双壳类经历了长期的迟缓复苏, 直到Anisian晚期才复苏、辐射[ 24]。Hautmann[ 80]等报道了广西乐业Shanggan剖面Griesbachian期的双壳类介壳层, 鉴定得到11属11种, 新生分子和内栖分子的较高比例、低的优势度指标等均指示晚二叠世大灭绝后双壳类的一个快速复苏过程。Chen等[ 95]运用定量化方法对下扬子地区的煤山、黄芝山、丫山、马家山、湖山等剖面晚二叠世末至早三叠世初的宏体生物群落进行了深入分析, 认为群落的演化与二叠纪—三叠纪之交的双幕式灭绝相耦合, 双壳类对腕足类优势度的取代在第一幕灭绝后发生于浅海区域, 第二幕灭绝后扩展到几乎所有的生境。
美国西部地区, 除二叠系—三叠系界线地层外, 下三叠统地层出露齐全, 主要分布于犹他州、内华达州、蒙大拿州等地, 主体为较浅水沉积, 划分为Dinwoody组、Moenkopi组和Thaynes组等。双壳类化石丰富、研究较为深入的层段为Smithian期的Sinbad灰岩段和Spathian期的Virgin灰岩段等, 属级分异度较高。本区早三叠世双壳类研究的早期阶段主要为属种描述[ 63, 64, 65], 后期的研究除系统描述[ 96, 97, 98]外, 多侧重于生态学方面[ 28, 35, 96, 97, 98]。Schubert和Bottjer[ 35]建立了早三叠世的底栖群落, 结果表明群落结构较为简单;Fraiser和Bottjer[ 28]分析了底栖群落的优势度(numerical dominance)和等级次序(Rank order)等指标, 指出双壳类 Claraia, Eumorphotis, Promyalina, Unionites等少数几个属占据了早三叠世介壳层的绝对优势。Hofmann等[ 96, 97, 98]则从定量化的群落分析方面, 指出美国西部地区早三叠世底栖群落在α分异度增长的同时, β分异度却没有显著的增长。
日本除发育远洋沉积外, 也发育底栖化石丰富的浅海碳酸岩沉积环境(日本西南部Maizuru块体)和碎屑岩沉积环境(日本东南部的Chichibu块体和South Kitakami山区), 这两种沉积类型均缺失下三叠统印度阶, 化石丰富的地层为下三叠统奥伦尼克阶。Maizuru块体下三叠统地层为Honodani组, 含14属双壳类化石[ 69, 71, 72, 73]。Chichibu块体不同地区的下三叠统地层分别命名为Iwai组、Shionosawa灰岩、Tao组、Gobangadake灰岩、Kamura组、Kurotaki灰岩等, 含化石层位基本等时, 含7属双壳类化石[ 70, 74]。South Kitakami山区下三叠统地层为Hiraiso组和Osawa组, 化石主要分布于Hiraiso组[ 77], 时代可能为Smithian期。总之本区的研究多为双壳类化石的系统描述。
双壳类化石丰富, 可以建立良好生物地层格架, 有利于进行区域乃至全球性的地层对比[ 110]。二叠纪—三叠纪之交双壳类生物地层方面的研究起始于20世纪80年代, 克什米尔地区[ 56]、阿尔卑斯地区[ 83, 84]、华南地区[ 85, 86, 103, 104, 105, 106]、越南北部[ 87, 107, 108]等地区相继进行了良好的双壳类生物地层建带工作(图1)。
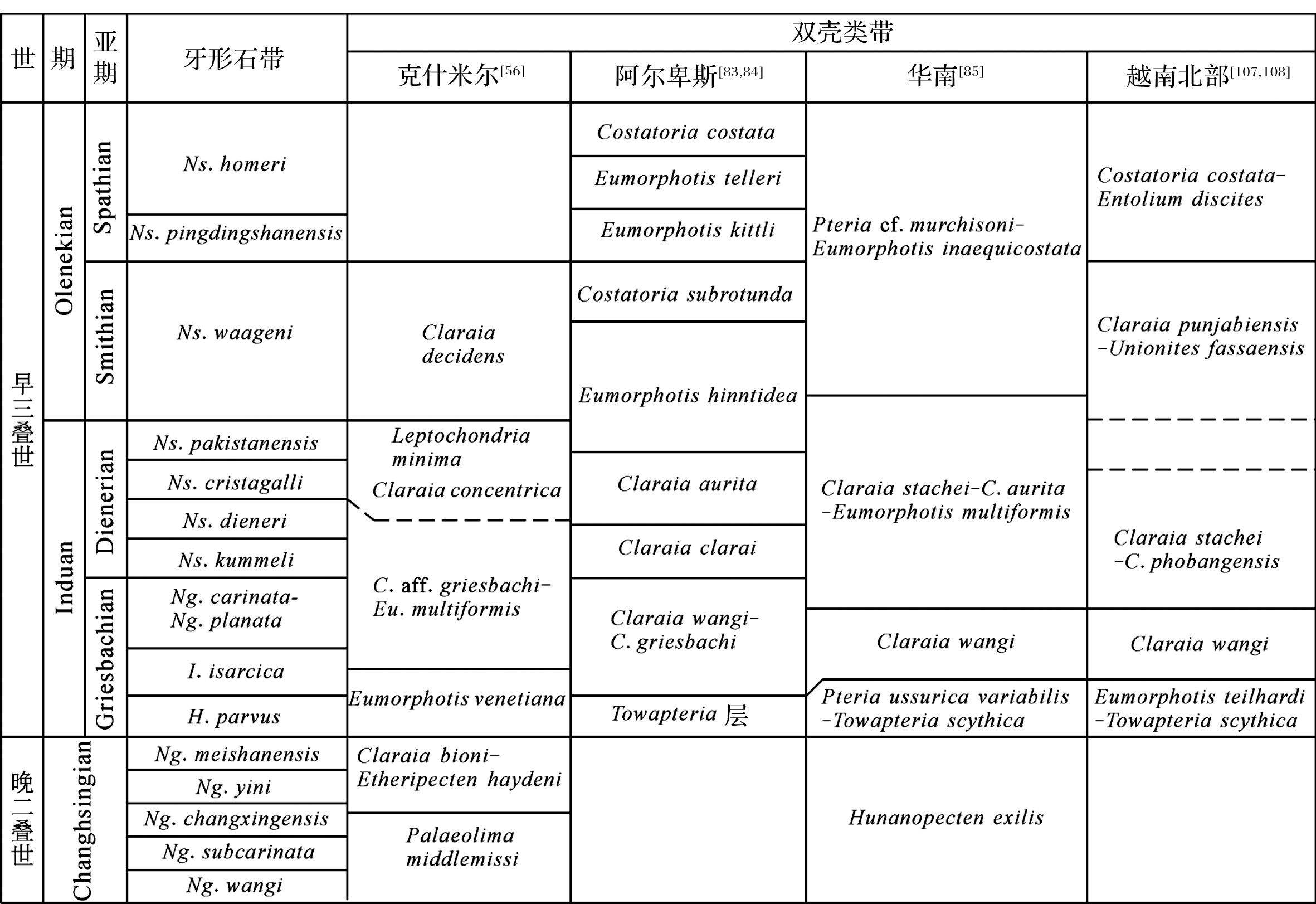 | 图1 克什米尔、南阿尔卑斯、华南和越南北部晚二叠世—早三叠世双壳类生物地层对比图Fig. 1 Correlation of Late Permian to Early Triassic bivalve zonations in Kashmir, Southern Alps, South China and North Vietnam |
在华南地区, Pteria ussurica variabilis- Towapteria scythica组合带是早三叠世最早的双壳类生物带, 大致位于二叠系—三叠系界线之上, Claraia wangi带之下[ 85]。该带主要组成分子有 Pteria ussurica variabilis, Towapteria scythica, Promyalina schamarae, Eumorphotis venetiana等。该组合带的典型分子同样见于克什米尔地区、意大利北部、越南北部及华南地区的浙江长兴黄芝山、江西宜春柏木、贵州盘县、重庆、云南宣威等地[ 19, 24, 108, 111, 112], 其时代归属有一定争议。浙江黄芝山剖面的 Eumorphotis venetiana- Towapteria scythica- Pteria ussurica variabilis组合带曾被认为是跨越了二叠系—三叠系界线[ 24], 进一步的研究表明, 该组合带对应于牙形石 Hindeodus parvus带, 时代为Griesbachian早期[ 19];云南宣威密德剖面下三叠统卡以头组中产出 Pteria ussurica variabilis等分子, 因在同层位地层中也采集到一枚菊石 Ophiceras, 因而其时代被对应于早三叠世 Ophiceras带[ 112];意大利Bulla剖面Werfen组底部产出双壳类 Eumorphotis, Promyalina, Towapteria等属, 因与牙形石 Hindeodus praeparvus共生, 而被归于晚二叠世末期[ 94];越南北部的 Eumorphotis teilhardi- Towapteria scythica组合带[ 87, 108]、克什米尔的 Eumorphotis venetiana带[ 56]大致可与华南地区进行对比。
Pteria ussurica variabilis- Towapteria scythica组合带之上是以 Claraia wangi, C. griesbachi, C. concentrica, C. stachei, C. aurita等分子建立的带, 这个类型的组合带广泛分布于全球各地区, 其时代为Griesbachian期到Dienerian早期。根据不同剖面的实际情况建立了以不同分子命名的生物带:湖北兴山地区的 Claraia stachei- C. griesbachi带和 C. concentrica- C. hubeiensis带, 该组合带相当于菊石 Ophiceras- Lytophiceras组合带和 Prionolobus- Gyronites组合带下部, 牙形石 Isarcicella stachei- I. isarcica组合带、 Neogondolella planata- N. carinata组合带、 Neogondolella discreta带和 Neospathodus kummeli带, 时代属于Griesbachian期到Dienerian早期[ 104];福建西部地区的 Claraia wangi, C. stachei带和 C. aurita带[ 86];浙江黄芝山剖面的 Claraia wangi带[ 19];贵州紫云四大寨剖面的 Claraia griesbachi- C. concentrica组合带和 Claraia stachei- C. aurita组合带[ 103];安徽巢湖地区的 Claraia griesbachi- C. concentrica组合带, 相当于菊石 Ophiceras- Lytophiceras组合带和牙形石 Hindeodus typicalis带上部- Neogondolella krystyni带- N. planata带, 时代属于Griesbachian期[ 105];贵州遵义地区的 Claraia concentrica- C. griesbachi组合带[ 106];意大利北部地区的 Claraia wangi- C. griesbachi带、 C. clarai带和 C. aurita带[ 83];越南北部地区的 Claraia wangi带和 C. stachei- C. phobangensis带[ 107]。另外, Claraia aurita分子在全球范围内主要繁盛于Dienerian期[ 113], 为不同剖面的地层对比提供了较精确的时代界定。
Dienerian晚期到Smithian期, 全球范围内的双壳类带较为分异, 但仍可进行地区性的地层对比。华南地区为 Eumorphotis multiformis带和 Eumorphotis inaequicostata带, 如湖北兴山地区的 Eumorphotis multiformis- E. inaequicostata组合带, 对应于菊石 Prionolobus- Gyronites组合带上部和牙形石 Neospathodus dieneri带, 其时代属于Dienerian晚期[ 104];福建地区的 Eumorphotis multiformis带和 E. inaequicostata- Entolium discites microtis- Eumorphotis fujiaensis组合带的下部[ 86];安徽巢湖地区的 Eumorphotis inaequicostata- E. huancangensis组合带, 对应于菊石 Gyronites- Prionolobus组合带、 Flemingites- Euflemingites组合带、 Anasibirites带和牙形石 Neospathodus kummeli带、 N. dieneri带、 N. waageni带, 其时代为Dienerian期至Smithian期[ 105];贵州遵义地区的 Eumorphotis multiformis带[ 106]等。意大利北部地区建立了 Eumorphotis hinnitidea带和 Costatoria subrotunda带[ 83, 84], 与之对应的是克什米尔地区的 Claraia decidens带[ 56]和越南北部的 Claraia punjabiensis- Unionites fassaensis带[ 107]。
Spathian期的双壳类带在不同地区有较大的差别, 该时期生物地层工作开展较好的区域是意大利北部, 依次为 Eumorphotis kittli带、 E. telleri带和 Costatoria costata带[ 83]。华南地区的双壳类被归纳为 Pteria cf. murchisoni- Eumorphotis inaequicostata组合带的上部[ 85], 但是其分布不具普遍性。安徽巢湖地区为 Guichiella angulata带和 Periclaraia circularis带, 其与菊石 Columbites- Tirolites带、 Subcolumbites带和牙形石 Neospathodus pingdingshanensis带、 N. homeri带、 N. anhuinensis带共生, 时代属于Spathian期[ 105];湖北兴山地区为 Posidonia circularis- P. cf. wengensis带[ 104];福建地区为 Eumorphotis inaequicostata- Entolium discites microtis- Eumorphotis fujiaensis组合带的上部[ 86]。由于受到岩相的控制, 华南地区双壳类属种较少, 很多地区采集不到化石, 影响了双壳类带的建立和对比。
显生宙期间, 双壳类遭受了数次规模较大的灭绝, 如奥陶纪—志留纪之交、泥盆纪—石炭纪之交、二叠纪—三叠纪之交、三叠纪—侏罗纪之交、白垩纪—古近纪之交等[ 114]。二叠纪—三叠纪之交的双壳类灭绝吸引了众多研究者的兴趣[ 85, 88, 89, 90, 92], 华南地区由于地层出露完整、化石丰富而研究程度较高。
有关双壳类灭绝的研究多基于科级和属级分异度, 较早的研究认为二叠纪—三叠纪之交双壳类的灭绝是一个自中二叠世开始的长期而逐步的过程, 且这次危机是中等程度的灭绝[ 85, 88, 89]。殷鸿福[ 85, 89]统计了华南地区及世界范围内双壳类和海扇科双壳类的属种分布, 认为双壳类的灭绝与腕足类、珊瑚等的几乎全部灭绝性质不一样, 其危机是一个自中二叠世开始的长期危机, 晚二叠世是危机逐步加深的过程, 至晚二叠世末期危机达到顶点。随后的研究表明, 二叠纪—三叠纪之交双壳类的灭绝过程是一个突变的过程。显生宙双壳类属级分异度演化曲线显示, 从早二叠世到晚二叠世末期, 双壳类属级分异度不断增长, 一直到晚二叠世末期发生大的生物灭绝[ 114]。华南地区的数据也指示晚二叠世长兴期的双壳类分异度(35科58属), 并不显著低于之前的茅口期约24科32属和吴家坪期35科65属[ 92]。
晚二叠世末期生物大灭绝被认为是单幕式或者双幕式的灭绝, 这些研究既涉及单个剖面、单个类群, 也包括多个剖面、多个类群生物。对于双壳类来说, 双幕式的灭绝模式或许更为合适。方宗杰[ 92]统计了华南地区灭绝前后的双壳类分异度的变化, 提出双壳类在第一幕灭绝中遭受了53.4%和96.5%的属级和种级灭绝率, 而在第二幕中属级灭绝率为22%。对华南地区多条剖面高精度的生物地层对比研究, 同样证实双壳类、腕足类、腹足类等均遭受了两幕灭绝, 与珊瑚、三叶虫等类群的单幕式灭绝形成鲜明的对比[ 20]。
在生物大灭绝时期, 双壳类的生态选择性灭绝, 可以有效的指示生物与环境之间的演化关系[ 114], 如在中—晚二叠世[ 115]、晚三叠世末期[ 116, 117]等。而在二叠纪—三叠纪之交生物灭绝时期, 双壳类的生态选择性灭绝研究较少, 很少从生态学角度探讨双壳类的灭绝过程及原因。李玲[ 90]从双壳类生活方式和捕食方式的角度探讨了双壳类的灭绝, 其研究表明深掘穴类、浅掘穴类、非翼蛤类的足丝固着类等与底质关系密切的双壳类受影响较大, 几乎完全灭绝, 进而归因于生活环境或底质变得动荡不安。但是, 其不足之处在于未包含华南地区长兴期全部的双壳类属, 未划分出不同的生态环境, 且双壳类生活方式的划分值得商榷。
晚二叠世生物大灭绝后, 双壳类的分异度达到低值, 生态类型简单, 灾后泛滥种繁盛, 幸存者占据主导地位, 指示双壳类处于残存期。随后, 双壳类即开始缓慢的复苏, 关于复苏的时间则争议很大, 研究的热点则集中于分异度和生态结构方面。
从双壳类属级分异度指标(表1)来看, 双壳类的完全复苏一直到中三叠世Anisian晚期[ 24, 118, 119]或者晚三叠世Norian期[ 116]才实现。McRoberts[ 116]统计了全球的双壳类数据, 发现Induan期有双壳类57属, Olenekian期66属, Anisian期98属, Ladinian期121属, Carnian期171属, Norian期165属, Rhaetian期143属(表1)。这种双壳类属级分异度的变化反映了晚二叠世大灭绝后的复苏发生在Ladinian期。而华南地区早Griesbachian期有双壳类20属, 晚Griesbachian期17属, Dienerian期22属, Smithian期22属, Spathian期10属, 早Anisian期35属, 晚Anisian期67属(表1), 显示双壳类的复苏发生在晚Anisian期[ 24], 以青岩动物群为代表, 发现有双壳类50余属100余种, 且生活方式多样, 伴生有腕足、腹足、菊石、海百合等16个门类的化石, 具有了自晚二叠世灭绝后生物复苏-辐射的特征[ 118, 119]。另外, 南阿尔卑斯地区的数据也显示出类似的模式, 其中Induan期有双壳类5属, Olenekian期有12属, 早Anisian期有15属, 晚Anisian期有32属, 同样指示双壳类的复苏发生于中三叠世晚Anisian期[ 93]。
双壳类的复苏时间比其他门类要晚很多, 牙形石在Dienerian期[ 120]、菊石和有孔虫在Smithian期即开始复苏[ 121, 122]。近年来, 美国西部、巴基斯坦等地奥伦尼克期的双壳类先后得到了详细的研究, 不仅属级分异度很高, 而且出现了很多中生代型新生分子, 或许可以指示双壳类在奥伦尼克期已经进入了一个较快速的复苏阶段[ 91, 96, 97, 98]。早三叠世双壳类化石分异度较低的原因可能包括不充分的采样、环境条件不利于化石保存等。
| 表1 晚二叠世长兴期—中三叠世安尼期不同地区双壳类属级分异度 Table 1 Diversity of Late Permian to Middle Triassic bivalves in different regions |
早三叠世华南各地双壳类生态类型较单调, 最为繁盛的是表栖足丝附着类型[ 25]。南阿尔卑斯地区Induan期双壳类只有2种生活方式:外栖足丝附着和内栖活动类型, 而到了Olenekian期增加到了4种(外栖足丝附着、外栖游泳、半内栖、内栖活动), 早Anisian期出现了外栖固着类型, 晚Anisian期出现了外栖钻孔类型[ 93]。通过对陈金华[ 24]中双壳类数据的分析, 作者发现Induan期有4种生活方式:外栖固着类、外栖活动类、半内栖类、内生浅掘穴类, 到了Olenekian期则增加到5种, 出现了内生深掘穴分子, 而到了中三叠世, 双壳类的生活方式仍然是5种。生态类型的逐步分异指示双壳类的逐步复苏, 但是指示意义并不显著。
二叠纪—三叠纪之交底栖生物群落结构方面的研究较少, 研究对象包括底栖生物群落和异地埋藏的介壳层。华南地区二叠系—三叠系界线层的底栖生物群落的定量化研究表明群落优势度的变化与两幕式灭绝相对应, 第一幕灭绝中双壳类对腕足类的取代发生在较浅海环境中, 第二幕灭绝后则发生在所有环境中[ 95]。Posenato[ 94]对阿尔卑斯地区的晚二叠世末期的底栖群落开展了类似的研究, 腕足类主导的群落多分布于近滨环境, 而双壳类主导的群落则多分布于较深水环境。美国西部地区早三叠世底栖生物群落的定量化研究显示整个早三叠世群落为低分异度、低复杂度, 且其组成分子多为简单的机会分子、灾难分子等[ 35]。
分布于近滨到风暴浪基面之间的介壳层虽然经过了机械分选, 不能代表原地的生态组合, 但是仍然可以反映生态群落优势度、丰度的变化[ 123]。美国西部地区Griesbachian期的介壳层是低分异度, 主要有双壳类 Claraia和 Promyalina及腕足类 Lingula组成。Dienerian至Smithian期的介壳层仍然是低分异度的, 主要有双壳类和小腹足组成。Spathian期介壳层主要组成分子有双壳 Promyalina, Permophorus及海百合茎。整个早三叠世介壳层的低分异度支持了复苏期长期环境压力的假说[ 123]。最近, 美国西部地区早三叠世不同时期的底栖群落的定量研究表明, 整个早三叠世底栖群落的α分异度逐步增长, 而β分异度较低, 未有显著的增长[ 96, 97, 98]。因而, 总体而言, 早三叠世的底栖生物群落定量化研究仍较少, 缺乏不同地区的群落结构对比研究。
小型化(Lilliput effect), 反映了生物大灭绝事件后生物个体的变小。在Urbanek[ 124]的最原始定义中, 小型化仅表现在同一个种中。在现在的定义中, 小型化表现为:①大个体分子的灭绝;②小个体分子的残存;③新生分子个体较小[ 100]。晚二叠世末生物大灭绝后, 有孔虫[ 125]、双壳类[ 100, 101, 126, 127]、腹足类[ 128, 129]、遗迹化石[ 100]等生物门类均表现出了小型化现象。Twitchett[ 100]认为晚二叠世生物大灭绝后的小型化发生在最初的两个牙形石带, 即 Hindeodus parvus带和 Isarcicella isarcica带, 其系统研究了早三叠世多种生物门类的小型化现象, 并对小型化的时间、特征、原因等进行了深入的探讨。Hayami[ 126, 127]根据《Treatise on Invertebrate Paleontology》一书中的双壳类属数据, 发现早三叠世双壳类平均大小是25.8 mm, 小于晚二叠世的27 mm, 也小于中三叠世的27.2 mm。但是这个研究从时间尺度上、从化石数量上均精度太低。意大利北部地区早三叠世双壳类的小型化研究程度较高, 其结果显示 Claraia, Unionites, Eumorphotis等属的个体大小具有变大的趋势[ 100, 101], 但是这些双壳类分子是否真的小型化仍值得深入探讨。
综上所述, 双壳类在二叠纪—三叠纪之交的地层中数量丰富、分布广泛, 其易于识别且研究历史较长, 在属种描述、灭绝—复苏模式、生态系演化等方面均取得了许多重要进展。但是, 该时期的双壳类研究仍然存在许多问题, 值得下一步深入思考。
①早三叠世双壳类生物地层具备良好的研究基础, 但是不同地区、不同相区的双壳类生物带的精确时代归属和对比问题仍亟待解决。双壳类化石在浅海碎屑岩相区较为丰富, 但因为其通常分布时限较长, 因此地层划分和对比的精度不及同时期的菊石和牙形石。但这类沉积相区通常缺乏牙形石, 菊石化石也比较少见或保存较差, 因此进一步挖掘双壳类的生物地层价值对于碎屑岩相区二叠纪—三叠纪界线及早三叠世地层学研究具有重要意义。
②二叠纪—三叠纪之交双壳类是否存在生态选择性灭绝及与当时的环境因素之间的关系仍需深入探讨。双壳类表现出不同的生态类别, 包括生活方式、摄食方式、壳质等, 有利于开展灭绝模式和生态选择性灭绝方面的研究, 从而进一步探讨导致双壳类灭绝的环境因素。
③晚二叠世大灭绝后至中三叠世安尼期, 双壳类迟缓复苏的过程及其原因值得深入分析。目前为止, 双壳类复苏模式的建立主要依据于地区性的属级分异度演化, 研究精度较低, 未考虑不同纬度区、不同水深区的分异度变化。三叠纪初双壳类的迟缓复苏是受控于内因(生物因素)还是外因(环境因素)的研究, 将能为生物与环境的协同演化提供关键性的实证。
④双壳类尤其是机会分子或灾难分子是否发生小型化仍需要进一步探索。有孔虫、腹足类等多个门类的生物被认为表现出了小型化现象, 但是双壳类作为一类抗灾变能力强, 在早三叠世广泛分布的生物类群, 其显然是二叠纪末大灭绝事件的受益者, 在早三叠世占主导的双壳类群中, 具有机会生物生态特征的灾难分子, 有时也发育出较大的生物个体, 但正常演变发展的双壳类群是否仍发生了小型化现象, 则值得深入剖析。
此外, 早三叠世一些占优势的灾难生物(如 Claraia等)不仅数量丰富, 分布广泛, 而且地区性分异也十分显著, 因而也导致了不同学者对这些类群的鉴定和分类工作上的混乱。目前急需要对这些重要类群进行精确系统的古生物学和分类学总结研究。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|
| [66] |
|
| [67] |
|
| [68] |
|
| [69] |
|
| [70] |
|
| [71] |
|
| [72] |
|
| [73] |
|
| [74] |
|
| [75] |
|
| [76] |
|
| [77] |
|
| [78] |
|
| [79] |
|
| [80] |
|
| [81] |
|
| [82] |
|
| [83] |
|
| [84] |
|
| [85] |
|
| [86] |
|
| [87] |
|
| [88] |
|
| [89] |
|
| [90] |
|
| [91] |
|
| [92] |
|
| [93] |
|
| [94] |
|
| [95] |
|
| [96] |
|
| [97] |
|
| [98] |
|
| [99] |
|
| [100] |
|
| [101] |
|
| [102] |
|
| [103] |
|
| [104] |
|
| [105] |
|
| [106] |
|
| [107] |
|
| [108] |
|
| [109] |
|
| [110] |
|
| [111] |
|
| [112] |
|
| [113] |
|
| [114] |
|
| [115] |
|
| [116] |
|
| [117] |
|
| [118] |
|
| [119] |
|
| [120] |
|
| [121] |
|
| [122] |
|
| [123] |
|
| [124] |
|
| [125] |
|
| [126] |
|
| [127] |
|
| [128] |
|
| [129] |
|