
{kind=link}
海山生物多样性研究进展与展望
[张均龙 , 徐奎栋
, 徐奎栋* ]
, 徐奎栋]
|
|


张均龙(1983-),男,山东潍坊人,助理研究员,主要从事海洋大型底栖生物生态学和软体动物分类学研究.E-mail:zhangjl@qdio.ac.cn
深海的海山生态系统支持着独特的生物群落,是海洋生态系统中物种扩散和进化的重要节点。由于研究的欠缺和认识的不足,国际间对于海山的区系和生物多样性认知存在较大分歧。存在海山特有种假说、物种源汇假说、孤岛假说、绿洲假说等各种假说,且均有争议。综述了海山生物区系和多样性研究的最新进展,指出了目前研究存在的缺陷和不足,并就我国未来有关海山生物区系和多样性研究提出了建议。
Seamounts support unique biological communities and might act as important nodes of species dispersal and evolution in deep ocean ecosystems. Seamounts have been one of the few characteristic ecosystems that remain underexplored in the abyssal world. However, our knowledge of seamount animals remains very limited due to the difficulty in access to technology, resulting in controversial explanation on their fauna, biodiversity, and biogeography. There have been several hypotheses on seamount biota, including the Endemicity Hypothesis, the SourceSink Hypothesis, the Insular Isolation Hypothesis, and the Oasis Hypothesis. We summarize the pros and cons of seamount fauna and biodiversity, and put forward proposals for future investigation on seamount ecosystem.
海山 (Seamounts) 通常是指深海大洋中位于水面以下、高度大于1000 m的隆起地形[ 1, 2],广义的海山也包括在海底中相对高度小于1000 m的海丘 (Hills or Knolls)。大多数海山是火山活动形成的,板块构造运动通常使海山呈链状分布或细长形集群出现。全球海洋中存在约20万个海山[ 3],超过1 000 m高的海山逾3万个[ 4],其中近半数分布在太平洋 (图1)。
近些年来,深海大洋的生态系统一直受到世界各国政府、科学界和社会公众的广泛关注[ 5]。海山是深海大洋中的独特生境,广泛存在于世界各大洋中,由于其特定的地理学特征和水文条件,造就了其独特的生物学特征,正成为海洋生物多样性研究的热点地区。国际海洋生物普查计划 (CoML) 从2005年开始针对海山开展了名为CenSeam的调查,获得大量新发现和研究成果[ 6]。目前对海山的调查远远不够,据估计全球迄今已被研究或调查过的海山不足300个,并且已有的调查大部分是针对靠近海面或者靠近陆架的海山[ 7]。对北太平洋和大西洋研究较多,而对海山分布众多的赤道太平洋的海山研究较少 (图1)。
海山栖息着几乎所有门类的动物 (表1)。Wilson & Kaufmann[ 10] 统计以往调查过的59座海山,共计发现了596种物种。Stocks[ 7]整合了全球对171座海山的调查,统计有1 971种无脊椎动物。目前,SeamountsOnline[ 9]中收录了246座海山调查所获的17 283条生物记录 (表1),涉及的有效种近2000种 (不包括悬疑种和未鉴定到种的记录)。海山存在的物种数可能要远远比目前记录的要多。迄今,在几乎所有调查的海山中都发现了新种[ 7]。
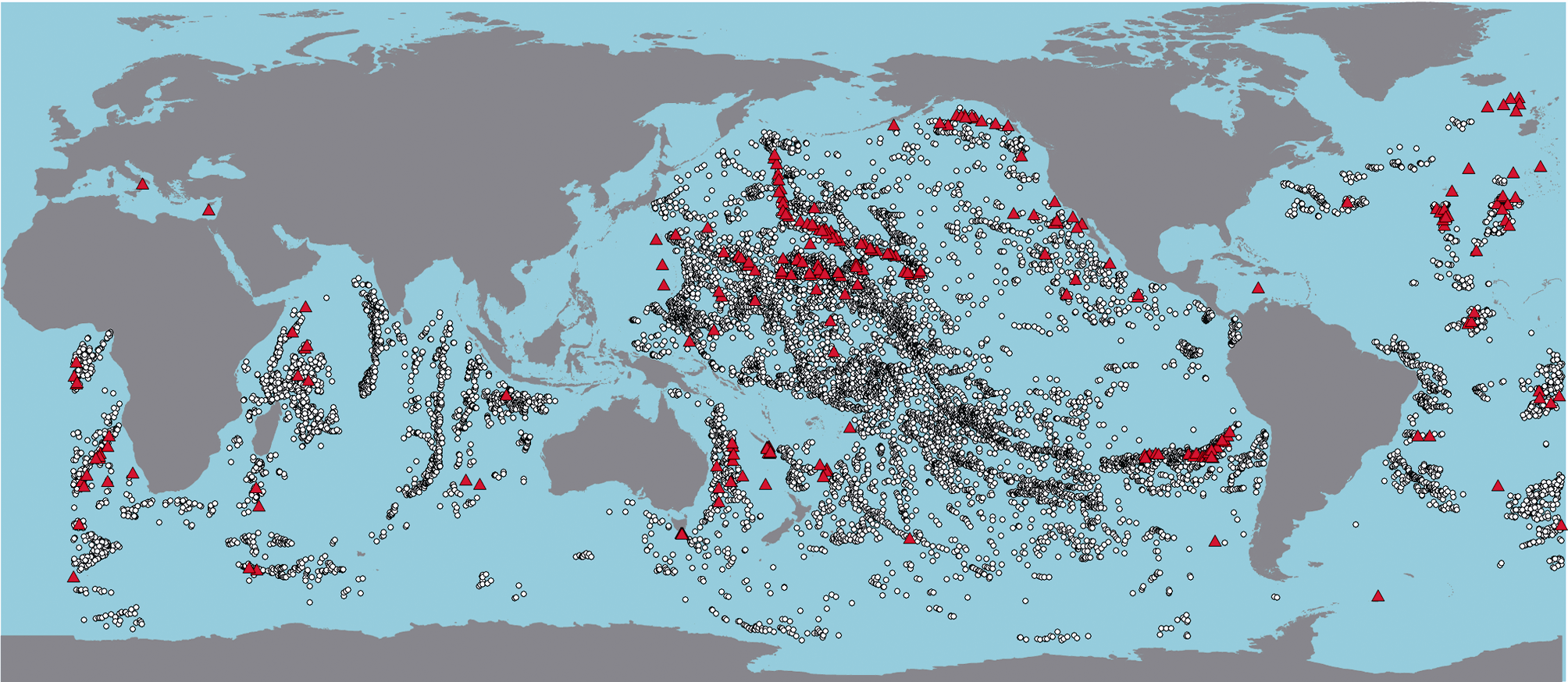 | 图1 全球高度大于1 000 m的海山(圆圈)及已采样过的海山(红色三角)(数据来自Global Seamount Database[ 8]和SeamountsOnline[ 9])Fig.1 Locations of global seamounts over 1 000 m in height (circle) and sampled sites (red triangle)(Data from Global Seamount Database[ 8] and SeamountsOnline[ 9]) |
海山大部分以岩石底质为主,以滤食性、附着或固着生活的物种为主[ 10, 11, 12],优势种有海绵、珊瑚、海葵、海笔、水螅、海百合等。其中,全球分布的深水珊瑚 (或冷水性珊瑚) 是研究较多的类群[ 13, 14]。平顶海山 (Guyots) 上的沉积环境大部分以软泥为主,生物以食碎屑者为主。海山的底内生物群落研究不多,大型底栖动物主要包括多毛类、甲壳动物、软体动物、纽虫、星虫等,小型底栖动物包括线虫、桡足类、铠甲动物和动吻动物等[ 15]。海山也是鱼类大量聚集的地方。
| 表1 海山中各类群记录数 Table 1 Taxonomic groups recorded on seamounts |
过去几十年中,海山生物多样性研究的一个重大发现是在海山中发现了特有种。海山特有种假说 (Endemicity Hypothesis) 认为,海山的地理隔离、水文特点以及独特生境等产生的一系列机制,使得海山生物区系在生态和进化上都与深海其他生境相隔离,从而形成了较高比例的特有种[ 16, 17, 18]。泰勒柱 (Taylor Column) 被认为是产生特有种现象的物种滞留机制之一[ 19, 20]。泰勒柱是指稳定的海流在流经海山时形成的逆时针环流,将物种幼虫滞留和聚集于海山区域,使海山种群与邻近区域种群隔离,造就了海山的高物种多样性。但最近有报道认为这只是由于调查不足引起的,海山的特有种比例较低[ 21, 22],甚至比大陆坡的特有种都少[ 23]。例如,在大西洋东部的海山,只有5.6%~6% 的多毛类是海山特有的[ 24, 25];大西洋海山中的腹足类也显示出较低的特有种比例[ 26, 27];新西兰附近海山特有种比例为5.5%~15%[ 28, 29];而有些研究甚至没有发现特有种[ 21]。
海山孕育了适宜特定类群的多样化生境,许多类群汇集于此并达到非常高的种群密度,成为相邻海山、邻近深海环境或陆坡的物种资源库[ 3, 18, 21, 30]。由此,海山群落也可能成为周围其他非海山区或非最适生境中维持种群密度的幼虫源[ 30]。这被称为海山物种源汇假说 (Source-Sink Hypothesis)。海山区与邻近的陆坡群落结构可能相差很大,但它们之间有很多共有物种。海山与大陆坡之间的距离对海山物种扩散的影响也因种类而异。O’Hara[ 23]和Samadi等[ 21]发现在距大陆坡100 km 到超过 1 000 km的海山中特有种比例也并不是非常高。而对于许多幼虫是卵黄营养的深海物种来说,由于幼虫阶段短暂,仅有几天时间,这些物种仅会在有限的范围内扩散,物种密度和海山群落的相似性随着与大陆边缘距离变远而降低[ 3]。但是,也有许多物种扩散能力较强,能够在它们适宜的水深中广泛分布[ 12, 21],这就使得无论是硬底海山[ 30, 31],还是软底为主的平顶山[ 24, 27],生物区系与大陆坡都非常相似。
海山的水文条件如泰勒柱等,将物种幼虫在海山区域滞留和聚集,使海山种群与邻近区域种群隔离[ 16],使海山如同海洋中的岛屿,有着与周围的深海平原或深渊截然不同的生物区系,形成了海山独特的物种多样性。这被称为孤岛隔离假说 (Insular Isolation Hypothesis)。但有的少量的遗传学研究[ 12, 21, 32]不支持种群隔离的观点。而绿洲假说 (Oasis Hypothesis) 认为,突出的地貌特征和水团的相互作用使该海域营养物质丰富[ 33, 34],使得这一区域的物种较多、种群密度较大,但没有与非海山区的种群形成隔离。
海山间的物种连通水平差别很大,在大洋尺度与局部范围内,海山与海岛、大陆边缘间连通性均存在争议。有些物种扩散能力有限,只能在局部范围内分布;而有些物种能扩散到几百到几千千米以外,在相隔较远的海山上都有分布。有研究显示海山种群与海岛陆坡种群以及大陆边缘的种群在遗传上是相同的[ 35~ 37]。但是在欧洲大陆架边缘和大西洋洋中脊的鱼类和头足类种群中发现了遗传差异[ 32, 38, 39]。在南大洋海山区和非海山区的鱼种群中也发现了遗传差异[ 40]。在太平洋海山中,即便相距遥远的珊瑚,线粒体基因中也存在很大的遗传相似性[ 12]。相反,在夏威夷同一珊瑚床内、同一岛或海山、不同岛以及岛与海山不同珊瑚床之间,珊瑚种群均存在显著遗传差异[ 41]。
海山是研究物理和生物过程相互作用造成物种隔离、分化和扩散的天然实验室[ 42]。迄今,对海山物种的扩散能力以及海洋学过程对其影响仍了解不足。在一定时间和空间尺度范围内对海山间生物种群连通性的研究是理解海山进化、物种丰度、群落种群变化的关键。尽管从遗传学角度针对海山种群连通性的研究较多,但目前仅局限于几个物种,并受到技术手段灵敏性的限制,因而对海山种群基因的连通性仍存争议。
影响海山生物区系的环境因素很多,水深是描述海山群落组成时最重要的因素之一[ 13, 43]。不同水深造就的环境因子差异,会影响底栖生物幼虫的沉降、存活率和生长。因而,海山群落随水深呈现出分带现象[ 23, 44, 45]。山顶位于真光层的海山,浮游植物的生产力能够在相对较浅的区域支撑一个物种丰富多样的底栖生物区系[ 3]。如果海山穿过缺氧区,海山顶部的生物将会非常稀少,但在缺氧区以下的陆坡生物将会非常丰富[ 46, 47]。从纬向上,海山区的蛇尾从热带到亚极地也呈现出类似浅海的生物梯度[ 23],这种现象在其他物种中是否具有普遍性尚需研究。
海山地形与海流的相互作用,会影响沉积物沉积速率、食物的丰富程度、浮游幼虫的滞留和聚集、海床的组成等,从而影响了底栖生物群落。珊瑚和其他滤食性生物通常聚集在地势较陡海流较急的区域,而底内生物主要以斑块状分布在的沉积物中[ 44, 48]。生活在硬底质区的珊瑚和海绵等固着生物也为其他生物提供生境,提高了底栖生物的多样性[ 49],而在锰结核区生物稀少[ 50]。在阿拉斯加海湾的三座海山中,生物多样性最高点出现在地形地貌最为多变的地区[ 51]。
很多海山是火山发育而成的并伴随有热液口,这为底栖生物群落创造了独特的物理化学条件,这一群落中有大量的化能合成生物。由于热液口的典型特征,在有热液和没有热液的海山,生物区系组成有显著差异。热液区的高生产力也影响群落中的物种组成和丰度[ 52]。
尽管海山底栖生物环境驱动因子研究取得了很大的进展,但驱动因子间的具体关系及相互作用并不十分清楚。还有很多潜在的重要环境驱动因素尚未引起足够重视,如海底山崩和海底火山爆发、岩石和沉积物的化学性质、海底有机物质和碎屑的分解、生物对沉积物的搬运作用等。
高生产力是海山的一个显著特征。传统的观点认为,上升流和泰勒柱引起了营养物质的上升提高了表层的初级生产力,这些能量又通过食物链传递到了更高的营养级中,增加了高营养级消费者的生物量。但有人认为上升流很少能穿过真光层而且在海山停留的时间不足以影响浮游植物的生长,带来的营养也无法产生足够的能量支撑海山区浮游动物和鱼类。海山区的高生物量可能并非源于当地的高初级生产力,而是由于海流与海山地形作用带来的悬浮性食物[ 53]、浮游动物[ 54]及其捕食者[ 55]等外来的输入引起的。
大量研究显示,海山在滞留垂直移动的浮游动物方面有重要的作用[ 54]。当夜间在深水层的浮游动物上升到表层时,有部分会被海流带到海山区的表层。黎明后,浮游动物开始在海山顶部下降,被海山区鱼类或其他以浮游动物为食的生物捕获。这一机制也许是支持海山浮游生物和底栖生物高生物量的关键营养途径。潮汐会在海山侧面形成海流。潮汐环流也许是深海海山的滞留机制。
海山硬底质区底栖动物群落以滤食性为主,这意味着这一底栖食物链较短且结构简单。但目前少量研究显示海山底栖食物网有多种营养结构,其食物链的长度与其他海区食物链长度相当,而且大型肉食性鱼类可能使这一食物链更长[ 15]。浮游动物在海山生物网能量传递中具有重要作用,中层鱼类是浮游动物的主要捕食者,它们在将能量传递到乌贼、肉食性的鱼类、海鸟、海洋哺乳动物等高等捕食者中起到重要的链接作用[ 56]。海山复杂的地形和海洋学特点使海山生物的生活方式多样,从而使得海山生物营养结构复杂。
生物物理耦合作用、复杂多变的水文条件和海山地形对海山生产力及营养结构的影响机制并不明确。后续研究需要综合一系列的物理作用以及对关键海山区域时间序列的数据,对底栖生物生产机制、浮游生物与底栖生物生产力相互作用机制、以及食物链营养传递过程等进行深入研究。
海山生态系统巨大的渔业资源量成为外海渔业经济的目标[ 11, 57],但也十分脆弱,受渔业捕捞作业尤其是底拖网作业破坏严重[ 17, 58, 59]。由于海山有富钴锰铁矿石、锰结核、多金属硫化物等,可用来开采铜、锌、铅和金、银等贵重金属的[ 60, 61]。将可能对海山矿物质的开采也对海山生态系统构成巨大的威胁。矿物质开采带来的物理扰动,直接或间接地影响了沉积物的悬浮和沉积,从而影响了生活于此的底栖生物[ 60]。
深海物种生命周期长、成熟期晚、生长率低、繁殖力差,呈现出明显的“K-选择”生长策略[ 62]。许多海山生物,尤其是形成生源生境的生物 (石珊瑚) 和使群落结构复杂化的生物 (大型的垂直生长的角珊瑚、柳珊瑚、海绵和海百合等) 更是如此[ 13, 15]。海山被认为是古老生物的避难所[ 7],有些生物生活了几百年甚至上千年[ 13]。许多海山物种没有浮游幼虫阶段[ 19],这表示它们扩散速度缓慢,只能在有限的或相近的海山间扩散。因此,海山底栖生态系统对人类干扰的耐受力较低,而且受干扰后恢复周期较长。根据现有研究,海山区底栖生物群落,尤其是跟陆地或其他海山相隔较远的海山,群落一旦受到破坏,恢复需要几十年甚至几百年的时间[ 63]。
已有海山研究受调查区域、取样设备、采样不足及分类研究程度等方面的限制,存在结果代表性不强、可比性差等问题。这是造成海山生态系统各种假说与争议的主要原因。从调查区域/方位上,已有的研究大多针对靠近陆架的海山及海山顶部,而对海山坡和海山底部研究较少,研究显示这些区域的生物群落显著不同。从生物粒级上,研究基本局限于巨型底栖生物 (megabenthos,即通过水底摄影照片即可清晰辨别类群的大型底栖生物),对较小的大型底栖生物相较少,对微型和小型底栖生物则鲜少涉及。从生物门类上,鱼类、甲壳类和珊瑚在海山区报道最多,其他门类较少,热液口以外海山区的微生物群落研究也非常少。而且,海山生物的分类研究十分不足,许多个体较小或稀有生物仅鉴定到较高的分类阶元。大部分海山生物的种类鉴定仅仅依靠形态特征,而表型变化较大的种或形态非常相似的种,鉴定困难。此外,目前大部分对海山的取样调查很大程度上仅关注物种的组成,对整个群落结构变化及功能的研究较少。总体上看,海山生物研究基本还处于调查层面,缺乏时间序列的观测数据和系统的研究。
过去我国对海山的研究基本上受制于深海探测能力的欠缺和研究经费的不足。我国已执行的大洋考察航次早已开始关注海山,尽管这些航次主要目的在于寻找资源 (矿产和生物资源),但环境和生物多样性也是重要内容。今年我国自主设计、自主集成研制的“蛟龙号”载人深潜器,在南海的蛟龙海山、太平洋的魏源海山和采薇海山拍摄了多种高清生物图像和底质资料,获得了许多新发现和宝贵的生物样品。我国最先进的综合科学考察船“科学”号的下水,以及先进海洋探测设备的装备,标志着我国已经具备深海探测与研究的能力。生物区系和生物多样性研究将是未来海山研究的首要问题,解决对海山生物的最基本认知,包括新物种的发现、物种多样性和特有种水平、与非海山区生物区系差异等。在此基础上,着眼海山在海洋生物地理上的作用及物种的扩布和隔离机制等,搞清驱动海山生态系统结构和多样性变化的主要因子,加强对深海生态系统的认识。
对于未来的海山生态系统调查研究,建议关注如下几方面的问题:
(1) 西太平洋发育了全球70%以上的弧后盆地,是全球海山系统分布最为集中的海域之一,是全球海洋生物多样性中心,也将是我国未来海山研究的优先战略选区[ 64]。
(2) 海山环境条件复杂,在水深超过2 000 m 的海山区取样非常困难,需要借助特殊的取样设备。因此,在未来的调查实践中,应注重促进科学和技术的结合发展[ 65],在取样设备和方法等方面应与国际保持一致,以便增强数据的可比性。
(3) 生物的个体大小及扩布能力是影响其分布的主要因子,而以往的研究结论或假说大多是基于巨型底栖生物的研究结果,由此往往得出不同甚至是截然相反的结论。未来研究应涵盖从巨型、大型到小型和微型底栖生物,理清不同粒级的底栖生物区系组成、多样性特点、食物网关系、能流传递作用及其环境驱动因子。
(4) 训练有素的分类专家仍是未来海山生物研究的先行者,新物种和新生命现象的发现和认知仍是海山研究的重点之一。分子遗传学是研究海山生物进化和连通性的重要手段。形态分类结合分子生物学技术对海山生物进行研究,将会为探索海山区高物种多样性成因,分析物种分布模式、扩散途径及进化等提供重要的基础数据。
鉴于海山生态系统的独特性及其在全球生态系统中的重要意义,加强海山生物区系和多样性的研究,不仅将获得大量分类新发现,揭示许多不为人知的新物种及其生命过程,填补我国目前在海山区生物新物种的发现方面的空白状态,而且还有助于厘清海山生物区系和多样性特点及其与环境的作用关系,为证实/证伪迄今的种种假说提出切实论据。同时,为探讨海山区物种多样性成因、物种分布模式和扩散途径、物种进化等提供基础数据,有助于加深对特殊生境生物多样性和生态系统的整体认识,为海洋生态系统和功能、生命现象与生命过程等提供新的视角。对海山的生物资源进行探测和研究,将是促认识和利用深海大洋,发掘战略性海洋生物资源的重要部分。
| [1] |
|
| [2] |
|
| [3] |
|
| [4] |
|
| [5] |
|
| [6] |
|
| [7] |
|
| [8] |
|
| [9] |
|
| [10] |
|
| [11] |
|
| [12] |
|
| [13] |
|
| [14] |
|
| [15] |
|
| [16] |
|
| [17] |
|
| [18] |
|
| [19] |
|
| [20] |
|
| [21] |
|
| [22] |
|
| [23] |
|
| [24] |
|
| [25] |
|
| [26] |
|
| [27] |
|
| [28] |
|
| [29] |
|
| [30] |
|
| [31] |
|
| [32] |
|
| [33] |
|
| [34] |
|
| [35] |
|
| [36] |
|
| [37] |
|
| [38] |
|
| [39] |
|
| [40] |
|
| [41] |
|
| [42] |
|
| [43] |
|
| [44] |
|
| [45] |
|
| [46] |
|
| [47] |
|
| [48] |
|
| [49] |
|
| [50] |
|
| [51] |
|
| [52] |
|
| [53] |
|
| [54] |
|
| [55] |
|
| [56] |
|
| [57] |
|
| [58] |
|
| [59] |
|
| [60] |
|
| [61] |
|
| [62] |
|
| [63] |
|
| [64] |
|
| [65] |
|